
c-Myb截短突变体的构建及其对c-Myc蛋白表达的调节
2014-11-29 04:16秦玺程龙饶春明高凯杨琦史新昌徐小洁叶棋浓王军志
生物技术通讯 2014年4期
秦玺 ,程龙,饶春明,高凯,杨琦,史新昌,徐小洁,叶棋浓,王军志
1.中国食品药品检定研究院 生物制品检定所,北京1 000502;2.军事医学科学院 生物工程研究所,北京 100850
转录因子c-Myb 由原癌基因c-Myb编码,分别由DNA结合区(DNA binding domain,DBD)、转录激活区(central transactivation domain,TAD)、负调节区(negative regulatory domain,NRD)和亮氨酸拉链构成,是一个进化上高度保守的蛋白[1],与白血病、结直肠癌、乳腺癌、黑色素瘤等恶性肿瘤的发生发展相关[2-4]。c-Myb 促进细胞的增殖,主要通过调控其靶基因的转录来实现自身功能。c-Myc是c-Myb 的靶基因,c-Myb 能够结合到c-Myc基因启动子的2个区域,调节c-Myc基因的转录和表达[5-8]。c-Myc 在20%的肿瘤患者中处于活化状态,c-Myc 表达的增强,会促进细胞恶变,最后导致肿瘤的发生[9-10]。临床标本中c-Myb 的表达与c-Myc 的表达呈正相关性,但c-Myb 各结构域对c-Myc 的调节及作用机制还不清楚。为了进一步明确c-Myb 各结构域对c-Myc 的调节,我们构建了c-Myb 不同结构域的截短突变体,分别命名为c-Myb-1、c-Myb-2、c-Myb-3、c-Myb-4、c-Myb-5和c-Myb-6,并在乳腺癌细胞中检测了这些突变体对c-Myc 表达水平的调节,为进一步研究c-Myb各结构域的功能及其在肿瘤中的作用机制奠定基础。
1 材料与方法
1.1 材料
人胚肾细胞293T、乳腺癌细胞MCF-7、真核表达载体pcDNA3.0-FLAG、c-Myb野生型质粒pcDNA3.0-FLAG-Myb、c-Myc由本室保存;大肠杆菌感受态细胞DH5α、细胞裂解液购自天根生化科技有限公司;DMEM 培养基和胎牛血清购自GIBCO公司;限制性内切酶KpnⅠ、XhoⅠ,DNA 连接酶,高保真Taq酶试剂盒购自NEB 公司;质粒提取试剂盒、胶回收试剂盒、PCR 回收试剂盒购自Promega 公司;c-Myc 抗 体、HRP-FLAG 抗体和GAPDH 抗体购自Sigma 公 司;SDS-PAGE 胶 和PVDF 转膜系统购自Invitrogen 公司;PCR 引物(表1)合成和DNA 测序由北京奥科鼎盛生物公司完成。
1.2 c-Myb截短突变体的构建
以野生型c-Myb为模板,分别进行重组PCR 扩增,胶回收PCR 产物;用KpnⅠ和XhoⅠ分别酶切PCR 产物和空载体pcDNA3.0-FLAG,胶回收酶切产物,16℃连接4 h,连接产物转化大肠杆菌感受态细胞DH5α,涂布含氨苄西林的LB 平板,37℃过夜生长;挑取平板上的克隆做菌落PCR鉴定,阳性克隆提质粒后用KpnⅠ和XhoⅠ酶切鉴定。
1.3 Western印迹
转染48 h 后收集细胞,加入细胞裂解液及SDS-PAGE 加样缓冲液,煮沸15 min,12 000 r/min离心2 min,上清液进行SDS-PAGE,电泳后将蛋白转至PVDF 膜,用5%脱脂奶粉封闭1 h,用一抗孵育1 h,TBST洗膜3次,每次7 min,二抗孵育1 h,TBST洗膜3 次,每次7 min,将含辣根过氧化物酶的ECL底物滴于膜上反应5 min,吸干后用X线胶片显影。

表1 PCR引物及序列
2 结果
2.1 c-Myb 截短突变体表达载体的构建及酶切鉴定
野生型c-Myb结构域如图1所示[14]。其中,DBD分为R1、R2和R3 区,均起到与DNA 结合的作用。用引物Y1和Y2 扩增c-Myb 的DBD和TAD 区基因,即为c-Myb-1基因序列,长度为978 bp;用引物Y1和Y3扩增c-Myb 的DBD 区基因,用引物Y4和Y5扩增c-Myb 的NRD 区前段基因,用引物Y1和Y5 重组PCR 将c-Myb 的DBD和NRD 前段基因扩增到一起,即为c-Myb-2基因序列,长度为833 bp;用引物Y1和Y3扩增c-Myb 的DBD 区基因,用引物Y6和Y7扩增c-Myb 的NRD 区后段基因,用引物Y1和Y7 重组PCR 将c-Myb 的DBD和NRD 区后段基因扩增到一起,即为c-Myb-3基因序列,长度为1160 bp;用引物Y1和Y5 扩增c-Myb 的DBD、TAD和NRD 区的前段基因,即为c-Myb-4基因序列,长度为1230 bp;用引物Y1和Y2 扩增c-Myb 的DBD和TAD 区基因,用引物Y6和Y7 扩增c-Myb 的NRD 区后段基因,用引物Y1和Y7 重组PCR 将这2 段基因扩增到一起,即为c-Myb-5基因序列,长度为1556 bp;用引物Y1和Y3 扩增c-Myb 的DBD 区基因,用引物Y4和Y7 扩增c-Myb 的NRD 区基因,用引物Y1和Y7 重组PCR 将c-Myb 的DBD和NRD 区基因扩增到一起,即为c-Myb-6基因序列,长度为1526 bp。将上述6 段PCR扩增片段与pcDNA3.0-FLAG 分别酶切、回收、连接、转化后,挑取单克隆PCR鉴定,阳性克隆提取质粒后进行双酶切鉴定,结果c-Myb-1~c-Myb-6酶切条带均在预期大小位置出现(图2)。将酶切鉴定为阳性的克隆测序,测序结果与预期序列一致,说明c-Myb各突变体构建成功。

图1 c-Myb野生型和截短突变体结构图
2.2 c-Myb突变体的表达鉴定
将c-Myb的各截短突变体阳性克隆提取质粒后转染293T 细胞,阴性对照转染pcDNA3.0-FLAG 空载体,阳性对照转染pcDNA3.0-FLAG-Myb,48 h 后收集细胞,裂解后用HRP-FLAG 抗体进行Western印迹检测,用GAPDH 抗体检测内参。结果显示,野生型c-Myb和各截短突变体在膜上相应位置均检测到明显条带,表明各c-Myb突变体均可表达(图3)。
2.3 c-Myb 各突变体对c-Myb 下游靶基因c-Myc 表达的影响
用野生型c-Myb和c-Myb 各突变体分别转染乳腺癌细胞株MCF-7,6 h后换液,在37℃培养48 h后收集细胞,裂解后做Western 印迹,结果见图4。相比于野生型c-Myb,含有TAD 区的突变体c-Myb-1、c-Myb-4、c-Myb-5 都能够促进c-Myc 的表达,而不含TAD 区的突变体c-Myb-2、c-Myb-3、c-Myb-6 则对c-Myc 没有促进作用。说明c-Myb 对c-Myc 的调控发生在c-Myb 的转录调控区,而DNA 结合区和负调节区则没有对c-Myc的调控作用。
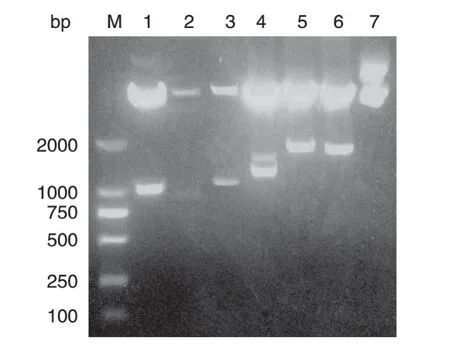
图2 c-Myb各突变体重组质粒的KpnⅠ/XhoⅠ双酶切电泳图谱
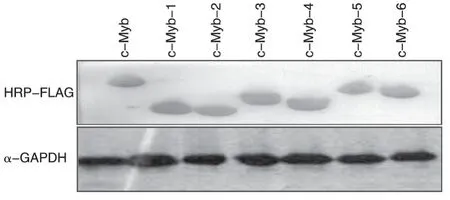
图3 Western印迹检测c-Myb及各突变体在293T细胞内的表达

图4 Western印迹检测MCF-7细胞中c-Myb及c-Myb各突变体对内源c-Myc表达的影响
3 讨论
c-Myb 包含DBD、TAD、NRD 等3 个结构域和1个亮氨酸拉链,在本研究中,我们构建了不同结构域的截短突变体,突变体均以DBD 为基础,加上TAD或NRD,并在乳腺癌细胞中检测了上述突变体对c-Myb 下游基因c-Myc表达水平的调节,发现含有转录激活区的突变体均能上调c-Myc 的表达,说明c-Myb 对c-Myc 表达的促进作用依靠c-Myb 的TAD 区来执行。c-Myb 对其他下游靶基因的调节是否也依赖于TAD结构域,还须实验来证明。
c-Myb 在乳腺癌的发生发展中发挥重要作用,在所有雌激素受体(ER)阳性的乳腺癌细胞中,c-Myb 均高表达[15-16],c-Myb 是雌激素受体阳性乳腺癌细胞增殖所必需的,在对化学诱导的乳腺癌细胞分化中,c-Myb 的表达被下调,显示了c-Myb 下调增强了乳腺癌细胞的分化和凋亡[17]。ERα和c-Myb 在乳腺癌细胞中的异常表达和相互作用,提示c-Myb 参与了雌激素诱导的乳腺癌发生、发展、演变等诸多过程[18]。本研究以乳腺癌细胞为模型,证明c-Myb 的TAD 结构域能够升高乳腺癌细胞中c-Myc 的表达,由于ERα也能够促进c-Myc 的表达,因此,c-Myb 对c-Myc 的调控是否需要ERα的参与,以及c-Myb 直接调控还是通过ERα间接调控c-Myc,还须进一步的实验证实。
构建c-Myb 不同结构域的突变体,深入探讨c-Myb 不同结构域的功能,有利于研究c-Myb 在恶性肿瘤中发挥功能的分子机制,能够更明确c-Myb 作为成药靶点的区域和调节作用,对于研发特异性高,毒性低的抗肿瘤靶向新药具有重要意义。
[1]Kobe B,Deisenhofer J.Crystal structure of porcine ribonuclease inhibitor,a protein with leucine-rich repeats[J].Nature,1993,366(6457):751-756.
[2]Weinstein Y,Ihle J N,Lavu S,et al.Truncation of the cmyb gene by a retroviral integration in an interleukin 3-dependent myeloid leukemia cell line[J].Proc Natl Acad Sci USA,1986,83(14):5010-5014.
[3]Thompson M A,Ramsay R G.Myb:an old oncoprotein with new roles[J].Bioessays,1995,17(4):341-350.
[4]Schomburg C,Schuehly W,Da Costa F B,et al.Natural sesquiterpene lactones as inhibitors of Myb-dependent gene expression:structure-activity relationships[J].Eur J Med Chem,2013,63:313-320.
[5]Bechard M,Dalton S.Subcellular localization of glycogen synthase kinase 3 beta controls embryonic stem cell self-renewal[J].Mol Cell Biol,2009,29(8):2092-2104.
[6]Minghetti P P,Norman A W.1,25(OH)2-vitamin D3 receptors:gene regulation and genetic circuitry[J].FASEB J,1988,2(15):3043-3053.
[7]陈蕊,张莹,赵丽.c-Myc 与c-myb 基因在白血病中的研究进展[J].肿瘤防治研究,2011,38(10):1207-1210.
[8]Wang M,Wei X,Shi L,et al.Integrative genomic analyses of the histamine H1 receptor and its role in cancer prediction[J].Int J Mol Med.2014,33(4):1019-1026.
[9]Nesbit C E,Tersak J M,Prochownik E V.MYC oncogenes and human neoplastic disease[J].Oncogene,1999,18(19):3004-3016.
[10]Li Z,Van Calcar S,Qu C,et al.A global transcriptional regulatory role for c-Myc in Burkitt's lymphoma cells[J].Proc Natl Acad Sci USA,2003,100(14):8164-8169.
[11]Wilson A,Murphy M J,Oskarsson T,et al.c-Myc controls the balance between hematopoietic stem cell self-renewal and differentiation[J].Genes Dev,2004,18(22):2747-2763.
[12]Hock H,Hamblen M J,Rooke H M,et al.Gfi-1 restricts proliferation and preserves functional integrity of haematopoiet-ic stem cells[J].Nature,2004,431(7011):1002-1007.
[13]Guo Y,Niu C,Breslin P,et al.c-Myc-mediated control of cell fate in megakaryocyte-erythrocyte progenitors[J].Blood,2009,114(10):2097-2106.
[14]Greig K T,Carotta S,Nutt S L.Critical roles for c-Myb in hematopoietic progenitor cells[J].Semin Immunol,2008,20(4):247-256.
[15]Cesi V,Casciati A,Sesti F,et al.TGFβ-induced c-Myb affects the expression of EMT-associated genes and promotes invasion of ER+breast cancer cells[J].Cell Cycle,2011,10(23):4149-4161.
[16]Mitra P,Pereira L A,Drabsch Y,et al.Estrogen receptor-α recruits P-TEFb to overcome transcriptional pausing in intron 1 of the MYB gene[J].Nucleic Acids Res,2012,40(13):5988-6000.
[17]Drabsch Y,Robert R G,Gonda T J.MYB suppresses differentiation and apoptosis of human breast cancer cells[J].Breast Cancer Res,2010,12(4):R55.
[18]Edavana V K,Penney R B,Yao-Borengasser A,et al.Fulvestrant up regulates UGT1A4 and MRPs through ERα and c-Myb pathways:a possible primary drug disposition mechanism[J].Springerplus,2013,2:620.
猜你喜欢
广州大学学报(自然科学版)(2019年1期)2019-05-07
生命科学研究(2018年1期)2018-05-29
食品科学(2018年10期)2018-05-23
安徽医科大学学报(2016年12期)2017-01-15
天津科技大学学报(2016年1期)2016-02-28
湖北师范大学学报(自然科学版)(2015年2期)2016-01-10
天津医科大学学报(2015年2期)2015-12-22
西南医科大学学报(2015年1期)2015-08-22
中国当代医药(2015年9期)2015-03-01
山东医药(2015年40期)2015-02-28
