
低温胁迫对玉米幼苗光合特性的影响
2012-08-08 12:22顾万荣庄文锋张道园
东北农业大学学报 2012年1期
杨 猛,魏 玲,胡 萌,顾万荣,庄文锋,张道园,孙 继,魏
(1.东北农业大学农学院,哈尔滨 150030;2.哈尔滨市农业科学院,哈尔滨 150070;3.重庆市南川区农业委员会,重庆 408400)
温度作为重要的环境因子之一,时刻影响玉米的生长发育。玉米生育期间的低温胁迫,是中国北方玉米产区的主要气象灾害,是高产稳产的主要限制因素之一。建国以来,黑龙江、吉林、辽宁三省 1969、1972、1976、1983、1987、1989、1992、2002、2009年因低温冷害损失粮食百亿斤,导致粮食大幅度减产,造成严重的经济损失[1-2]。国内外学者对低温胁迫从叶绿素、蛋白质、活性氧代谢、内源激素平衡、显微超微结构、分子反应机制等诸多方面进行了广泛研究[3-4],低温胁迫下过氧化物酶、过氧化氢酶、超氧化物歧化酶活性以及丙二醛和蛋白质含量均有增加[5],弱光胁迫、低磷胁迫、水分胁迫等胁迫条件下的光合特性也有很多报道[6-8],弱光、盐胁迫均导致辣椒叶片净光合速率(Pn)、气孔导度(Gs)、蒸腾速率(Tr)下降,各逆境导致净光合速率下降的限制因子不同[9]。但关于玉米低温胁迫下光合特性的变化,尤其是叶绿素荧光参数的变化未见报道。因此,本研究选用3个玉米品种,在苗期进行低温处理,测定光合参数的变化,探讨这些变化与不同品种耐低温特性的关系,揭示低温伤害机理,为玉米耐低温育种提供理论依据。
1 材料与方法
1.1 材料
选用3个玉米品种:金玉5、兴垦3、吉单198。分别来源于黑龙江省久龙种业、德农种业、吉农高新北方农作物优良品种开发中心。
1.2 试验设计
播种育苗:采用室外盆栽播种,盆直径21 cm,高20 cm,每盆保苗8株,每个品种5次重复。苗期正常管理。
低温胁迫处理:玉米长到三叶一心时,在人工智能培养箱(HPG-280HX)中进行低温处理,设3个温度梯度,分别为18℃/9℃、16℃/7℃、14℃/5℃(昼/夜)。以室外正常生长幼苗为对照(CK),对照在处理期间的日平均温度为25℃。每个温度水平分别处理2、4、6、8、10 d。
1.3 方法
1.3.1 光合指标的测定
选距生长点第2片完全展开叶,应用LI-6400便携式光合仪测定叶片净光合速率(Pn)、气孔导度(Gs)、蒸腾速率(Tr)、胞间CO2浓度(Ci)、水分利用率(Water use efficiency,WUE)。
1.3.2 叶绿素荧光的测定
用德国产PAM-210便携式测定。叶片暗适应20 min后设置原初的荧光参数,关闭光化学光和远红光,打开测量光测定初始荧光(Fo),再打开一个饱和脉冲光测定最大荧光(Fm),然后打开光化学光(光强600 μmol·m-2·s-1)照射叶片,当稳态荧光(Fs)出现后,再打开一个饱和脉冲光测定光下最大荧光(Fm),最后关闭光化学光,打开一次远红外光测定光下最小荧光(Fo),计算出可变荧光(Fv)、PSⅡ最大光能转换效率(Fv/Fm)和光系统PSⅡ的潜在活性(Fv/Fo)。每个样品重复测定3次。
1.4 数据统计
采用Excel软件进行数据分析。
2 结果与分析
2.1 低温胁迫对玉米幼苗叶片Pn的影响
由图1可知,与对照相比,低温胁迫降低了玉米幼苗叶片的Pn。3个品种随着低温胁迫程度和胁迫时间的加长,Pn一直在下降,随着低温胁迫的加强,Pn下降的越快。各个品种降低的幅度不同,吉单198在第4天有一个迅速下降的过程,兴垦3在第6天有一个迅速下降的过程,金玉5一直都是在缓慢的下降,14℃/5℃低温胁迫的第10天时,吉单198、兴垦3、金玉5分别下降了41%、35%、32%。

图1 低温胁迫下玉米幼苗光合速率的变化Fig.1 Change of maize seedlings Pn under low temperature stress
2.2 低温胁迫对玉米幼苗Fv/Fm的影响
Fv/Fm表示PSⅡ最大光能转换效率,常用于度量植物叶片PSⅡ原初光能转换效率。如图2所示,随着低温胁迫时间的延长,不同品种玉米叶片的Fv/Fm整体上均呈下降趋势。具体表现为2~6 d时Fv/Fm先缓慢下降,6 d后迅速下降。不同温度处理下,温度越低,Fv/Fm越低,各温度下Fv/Fm高低顺序为:CK>18℃/9℃>16℃/7℃>14℃/5℃。Fv/Fm降低,表示其光能转换效率降低,说明低温胁迫时间延长加剧了玉米幼苗叶片的光抑制,从而影响了光合作用。对照温度下,3个品种Fv/Fm随处理时间的延长变化幅度不明显。不同低温处理下,随着低温胁迫时间的延长,3个品种Fv/Fm均呈下降趋势。温度越低,下降幅度越大。

图2 低温胁迫下玉米幼苗Fv/Fm的变化Fig.2 Change of maize seedlings Fv/Fm under low temperature stress
2.3 低温胁迫对玉米幼苗叶片Fv/Fo的影响
Fv/Fo用来表示光系统PSⅡ的潜在活性。如图3所示,低温胁迫下,对照的Fv/Fo没有明显变化。随着低温胁迫时间延长,各处理玉米叶片的Fv/Fo整体上均呈下降趋势,表明低温胁迫破坏了光合系统的正常运行,降低了PSⅡ的潜在活性。但各温度处理下幼苗的变化程度存在差异。14℃/5℃下降程度最大,18℃/9℃下降程度最小。对照温度下,3个品种Fv/Fo最高,并且随时间的延长变化幅度不明显。同一低温处理下,随着低温胁迫时间的延长,3个品种Fv/Fo变化不同,但整体上呈现下降趋势,下降幅度表现为吉单198>兴垦3>金玉5,下降幅度越大说明耐低温能力越弱。
2.4 低温胁迫对玉米幼苗叶片气孔导度的影响
由图4可知,与对照相比,3个玉米品种Gs均随低温胁迫时间的增加呈下降趋势。吉单198、兴垦3、金玉5在第10天18℃/9℃处理气孔导度分别下降了20.7%、16.3%、14.4%,16℃/7℃处理下降25.9%、21.8%、20.7%,14℃/5℃处理下降33.8%、27.4%、25.9%。吉单198与其余2个品种相比,在各个低温胁迫下的下降幅度最大,这说明低温胁迫对吉单198气孔导度的影响最大。3个品种随低温胁迫强度的增加,气孔导度下降幅度也随之增大,温度越低对玉米幼苗气孔导度影响越大。

图3 低温胁迫下玉米幼苗Fv/Fo的变化Fig.3 Change of maize seedlings Fv/Fo under low temperature stress
2.5 低温胁迫对玉米幼苗叶片蒸腾速率的影响
3个玉米品种在低温胁迫下Tr的变化呈下降趋势,且与光合速率和气孔导度的变化规律一致(见图5)。与对照相比,3个玉米品种Gs均随低温胁迫时间的增加呈下降趋势,并且低温强度越大下降的幅度越大。在低温胁迫下,吉单198前6 d的下降幅度大,之后缓慢下降;兴垦3前8 d下降幅度大,随后缓慢下降;而金玉5在8 d后Tr迅速下降。

图4 低温胁迫下玉米幼苗气孔导度的变化Fig.4 Change of maize seedlings Gs under low temperature stress
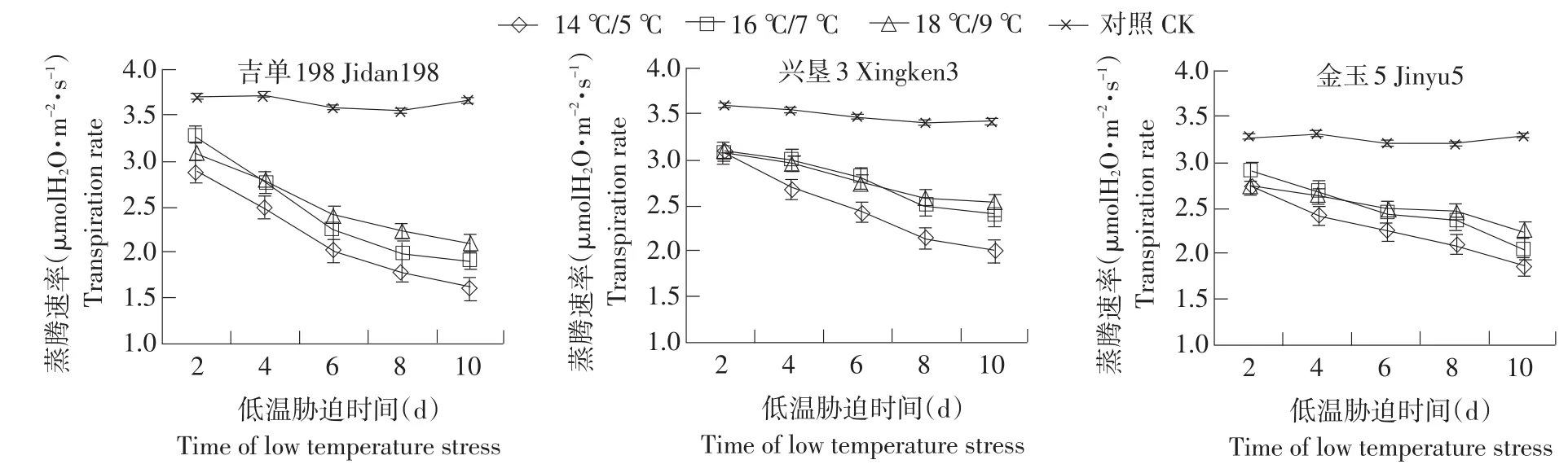
图5 低温胁迫下玉米幼苗蒸腾速率的变化Fig.5 Change of maize seedlings Tr under low temperature stress
2.6 低温胁迫对玉米幼苗胞间CO2浓度(Ci)的影响
如图6所示,对照处理中,3个品种的胞间CO2浓度对照处理相差不大,都在270~300 μg·mL-1之间。18℃/9℃、16℃/7℃低温处理下3个品种都有逐渐降低的趋势,14℃/5℃低温处理下呈现出先降低后升高的趋势。14℃/5℃低温处理下3个品种在第4天开始上升,在处理后第10天金玉5达到最高值330 μg·mL-1。低温强度不同,Ci的变化规律也不同,随着低温强度的加大,Ci有增加的趋势。

图6 低温胁迫下玉米幼苗胞间CO2浓度的变化Fig.6 Change of maize seedlings Ci under low temperature stress
2.7 低温胁迫对玉米幼苗叶片水分利用率的影响
如表1所示,在低温胁迫条件下,3个品种的WUE都高于对照,说明在低温胁迫下,植株可以通过提高WUE这个途径缓解逆境造成的部分损失。各个品种随着胁迫时间的增加,WUE呈现出先增加后稳定的趋势,18℃/9℃、16℃/7℃、14℃/5℃低温处理的WUE差异不大。胁迫10 d各个低温处理吉单198增加26%~36%,兴垦3增加15%~21%,金玉5增加15%~19%。18℃/9℃、16℃/7℃、14℃/5℃处理和对照都达到显著差异。

表1 低温胁迫下玉米幼苗水分利用率的变化Table 1 Change of maize seedlings WUE under low temperature stress
3 讨论
人工智能培养箱在人工模拟鉴定中对光照强度与时间长短、温度强度和时间长度和空气湿度都能进行准确的调控,已经成为人工模拟法研究作物温度胁迫的首选。同时也存在着局限性,首先人工智能培养箱温度范围设置存在局限性,最低温度只能设置5℃,限制了实验设计的最低温度,再次,人工气候室的空间容量有限,生理指标的测定需要大量的材料,多个箱体同时使用造成被测材料存在时间和空间的差异。本试验低温处理采用人工智能培养箱来处理,虽然会存在一定的误差,但能够模拟自然状态,不会影响到试验的准确性。
光合作用是植物生长发育的基础,是植物合成有机物质和获得能量的根本源泉,可以反映植株生长势和抗性强弱[10-11]。低温几乎影响作物光合作用的各个环节,包括光能吸收、气体交换、碳同化过程等,一般情况下,低温胁迫处理的植株叶片Pn、Gs、Tr明显小于对照[12],导致Pn降低既有气孔因素也有非气孔因素。低温处理导致玉米幼苗Pn下降的同时,Ci下降,说明Pn的下降由气孔因素引起。低温处理导致玉米Pn下降的同时,Ci先降低后升高,表明Pn下降由气孔因素逐渐转变为非气孔因素占主导地位[9,13]。从本试验结果可以看出,3个玉米品种Pn在低温胁迫下的变化趋势与前人研究结果一致,说明低温处理明显降低了玉米的光合速率,在18℃/9℃、16℃/7℃低温处理下为气孔因素引起,14℃/5℃低温处理下为非气孔因素引起。
叶绿素荧光可以利用植物体内叶绿素作为天然探针,快速、灵敏、无损伤地研究和测定光合生理状况以及多种因子对植物光合作用的影响,已成为人们研究光合代谢的最新手段,极大地推动了光合过程中光能利用、逆境条件下光合作用的光抑制、光损伤及其防御机制的研究[6]。Fv为可变荧光,Fm为最大荧光,它们的比值Fv/Fm代表PSⅡ光化学的最大效率或PSⅡ原初光能转化效率[14],低温胁迫可导致Fv/Fm下降,与前人研究结果一致。Fv/Fo是Fv/Fm的另一种表达形式,也可以作为反映低温胁迫程度的理想指标[15],本研究中Fv/Fo的变化趋势与Fv/Fm相同(见图2、3),耐低温能力强的品种能够减轻低温胁迫对玉米叶片PSⅡ反应中心的伤害,提高光能转化效率,从而提高玉米的光合能力。Fv/Fm和Fv/Fo在低温胁迫下下降,说明PSⅡ活性中心受损,Pn快速下降,本试验中吉单198在第4天有一个迅速下降的过程,兴垦3在第6天有一个迅速下降的过程,金玉5一直都是在缓慢的下降,说明在低温前期植株通过降低气孔导度(Gs)、蒸腾速率(Tr)和增大水分利用率(WUE)的自我保护机制进行调节,吉单198在第4天、兴垦3在第6天自我保护机制失效,导致光合速率迅速下降。本试验是在受控条件下对玉米幼苗进行的研究,低温胁迫下不同玉米品种全生育期的光合代谢调控还需要进一步研究。
4 结 论
随着低温胁迫处理时间的延长和程度的加强,3个品种表现出一致的规律,光合速率(Pn)、气孔导度(Gs)、蒸腾速率(Tr)、PSⅡ最大光能转换效率(Fv/Fm)和光系统PSⅡ的潜在活性(Fv/Fo)呈下降趋势,胞间CO2浓度(Ci)18℃/9℃、16℃/7℃低温处理下3个品种都有逐渐降低的趋势,14℃/5℃低温处理下呈现先降低后升高的趋势,水分利用率(WUE)先增高之后趋于稳定。
低温胁迫使光合作用降低,叶光系统Ⅱ(PSⅡ)活性中心受损,植株通过调节气孔导度(Gs)、蒸腾速率(Tr)、胞间CO2浓度(Ci)和增大水分利用率(WUE)的自我保护机制进行调控。本研究结果说明,参试的3个玉米品种在低温胁迫条件下光合指标的变化均有一定规律性,其品种间具有显著的差异。这些研究结果对玉米新品种培育和利用都具有重要意义。
[1] 王瑞,马凤鸣,李彩凤,等.低温胁迫对玉米幼苗脯氨酸、丙二醛含量及电导率的影响[J].东北农业大学学报,2008,39(5):20-23.
[2] 魏湜,王玉兰,杨镇.中国东北高淀粉玉米[M].北京:北京农业科学出版社,2010:122-124.
[3] 杨德光,唐心龙,席景会,等.植物在低温胁迫下的分子反应机制研究进展[J].玉米科学,2009,17(2):99-101.
[4] 简令成,卢存福,李积宏,等.适宜低温锻炼提高冷敏感植物玉米和番茄的抗冷性及其生理基础[J].作物学报,2005,31(8):971-976.
[5] 王小丽,裴玉贺,郭新梅,等.低温胁迫下玉米幼苗的几种生理生化指标的变化[J].植物生理学通讯,2009,45(5):487-490.
[6] 刘明,孙世贤,齐华,等.水分胁迫对玉米苗期叶绿素荧光参数的影响[J].玉米科学,2009,17(3):95-98.
[7] 常敬礼,杨德光,谭巍巍,等.水分胁迫对玉米叶片光合作用的影响[J].东北农业大学学报,2008,39(11):1-5.
[8] Schreiber U,Gademann R.Assessment of photosynthetic performance of prochloron in lissoclinum patella in hospite by chlorophyⅡfluorescence measurements[J].Plant and Cell Physiology,1997,38:945-951.
[9] 张志刚,尚庆茂.低温、弱光及盐胁迫下辣椒叶片的光合特性[J].中国农业科学,2010,43(1):123-131.
[10] Sharp R E,Poroyko V,Hejlek L G.Root growth maintenance during water deficits:physiology to functional genomies[J].J Exp Bot,2004,55:2343-2351.
[11] 庄文锋,魏湜,赵东旭.稀植栽培下春小麦旗叶光合性状的变化规律[J].东北农业大学学报,2008,39(3):22-25.
[12] 杨华庚,林位夫.低温胁迫对油棕幼苗光合作用及叶绿素荧光特性的影响[J].中国农学通报,2009,25(24):506-509.
[13] 朱新广,王强,张其德.冬小麦光合功能对盐胁迫的响应[J].植物营养与肥料学报,2002,8(2):177-180.
[14] 朱先灿,宋凤斌,徐洪文.低温胁迫下丛枝菌根真菌对玉米光合特性的影响[J].应用生态学报,2010,21(2):470-475.
猜你喜欢
农业工程学报(2022年3期)2022-04-16
干旱地区农业研究(2022年1期)2022-01-28
西南农业学报(2021年7期)2021-08-23
节水灌溉(2021年5期)2021-05-26
生态学报(2021年6期)2021-04-25
农业工程学报(2021年3期)2021-04-15
节水灌溉(2020年9期)2020-09-16
中国农业科技导报(2020年12期)2020-03-15
装备制造技术(2019年12期)2019-12-25
科技创新与品牌(2019年12期)2019-02-06
