
苏云金芽胞杆菌cry1Aa19基因的克隆、表达及杀虫活性分析
2012-08-08 12:23李海涛刘东明高继国
东北农业大学学报 2012年1期
罗 玲,李海涛,刘东明,高继国
(东北农业大学生命科学学院,哈尔滨 150030)
苏云金芽胞杆菌(Bacillus thuringiensis,Bt)是一种分布极其广泛且在其生活史中能形成芽胞和产生多种蛋白晶体的革兰氏阳性细菌[1]。Bt是当今微生物农药中研究最为深刻、应用最为广泛、对环境最为友善、商业化程度最高的微生物杀虫剂[2-3]。因此,利用Bt基因可以大量减少化学杀虫剂的使用[4-5]。目前已报道的杀虫晶体蛋白(Insecticidal crystal protein,ICPs),也称δ-内毒素(delta-endotoxin),有564种,分为68类Cry蛋白和3类Cyt蛋白,分属229种模式蛋白[6]。它的形状、结构和大小均与其毒力有着密切关系[7]。Cry晶体蛋白被敏感幼虫吞食后,在幼虫肠道碱性环境和蛋白酶的作用下释放出活性毒素,可与昆虫中肠上皮细胞的特异受体结合并形成孔道,破坏细胞渗透压平衡,最终导致昆虫死亡[8]。因此,对杀虫晶体蛋白的挖掘及研究具有非常重要的实际价值。目前已有多种cry基因用于高效广谱杀虫工程菌的构建和杀虫转基因植物的培育,部分商品化的工程菌和转基因植物已带来明显的经济效益和生态效益。由于Bt基因在构建杀虫工程菌株和转基因植物应用中的巨大潜力,有关高毒力和新型杀虫活性的cry基因的分离、克隆及开发研究已成为国内外Bt研究和利用的一大热点。
最近几年,已有一些Bt菌株从东北地区的多个自然保护区中被分离出来,东北农业大学与中国农业科学院植物保护研究所合作,从黑龙江省五常市拉林镇附近分离到一株Bt菌株LS-R-21,并从该菌株中分离出基因cry1Aa,进行了序列比对分析,在大肠杆菌中进行了基因的表达,表达产物的生物活性测定结果表明该基因对鳞翅目菜蛾科小菜蛾(Plutella xylostella)具有较高毒力。本研究通过对LS-R-21分离株进行形态和遗传特性分析,为LS-R-21分离株作为防治鳞翅目害虫的生物农药菌株,及培育转基因抗虫作物奠定良好的基础。
1 材料与方法
1.1 材料
1.1.1 菌株与质粒
大肠杆菌JM109及Rosetta(DE3)由本实验室保存;表达载体pEB由中国农业科学院植物保护研究所惠赠。
1.1.2 培养基
LB液体培养基:胰蛋白胨10 g,酵母提取物5 g,氯化钠10 g,加水定容至1 000 mL,pH 7.2,15磅灭菌20 min。
LB固体培养基:LB液体培养基加入1.3%琼脂粉。
1.1.3 主要试剂
TaqDNA聚合酶、限制性核酸内切酶、dNTP购自TaKaRa公司,KOD高保真酶购自TOYOBO公司,PCR引物由上海生工公司合成,DNA凝胶回收试剂盒和DNA连接酶购自上海生工公司。其他药品均为国产分析纯化以上产品。
1.1.4 供试昆虫
小菜蛾(Plutella xylostella)是由中国农业科学院植物保护研究所提供的标准化试虫。
1.2 方法
1.2.1 Bt LS-R-21菌株晶体形态的观察
将Bt LS-R-21菌株接种于1/2 LB固体培养基上,30℃培养48 h后,进行涂片、烘干固定,石炭酸复红染液染色3 min,清水冲洗,100倍油镜进行镜检。
1.2.2 PCR-RFLP体系鉴定Btcry基因
参照文献[9-12]的方法进行。
1.2.3cry1Aa全长基因的PCR扩增
根据已公布的cry1Aa类基因的全长序列,通过其同源性的比较,设计出用于扩增全长序列的引物:
上游引物(1-5):5′ATGGATAACAATCCGAAC ATC 3′
下游引物(1-3):5′TTCCTCCATAAGGAGTAA TTCC 3′
PCR热循环参数为:94℃预变性5 min;94℃变性1 min,50℃退火1 min,72℃聚合4 min,循环30次;72℃聚合10 min。取5 μL PCR产物于0.7%琼脂糖凝胶上电泳,检测PCR产物。
1.2.4 原核表达载体的构建
回收KOD plus扩增所得cry1Aa全长基因的PCR产物,及pEB载体用EcoⅠ30Ⅰ进行单酶切的产物,两者4℃连接过夜。将连接产物转入CaCl2法转化制备的感受态E.coliJM109中,涂布在氨苄抗性的LB平板上,37℃、12 h,PCR鉴定及酶切分析,筛选出含cry1Aa基因的阳性重组质粒。
1.2.5 序列测定及分析
选取阳性重组克隆由大连宝生物工程有限公司完成序列测定,用NCBI BLAST、DNAMAN等软件分析序列。
1.2.6cry1Aa基因在大肠杆菌中的表达
将获得的阳性重组质粒转化到感受态E.coliRosetta(DE3)中,230 r·min-1、37 ℃培养。加入0.5 mmol·L-1IPTG 进行诱导表达,20 mmol·L-1Tris-HCL悬浮后超声破碎,离心收集上清和沉淀分别进行SDS-PAGE检测分析[13]。同时用0.5 mmol·L-1IPTG诱导含pEB空载体转化的Rosetta(DE3)作为阴性对照。
1.2.7 Cry1Aa蛋白杀虫生物活性测定
将甘蓝叶片用清水洗净晾干,选取鲜嫩一致的甘蓝叶片,在稀释好的待测样品中(浓度梯度为 0.5、0.25、0.125、0.0625、0.03125、0.015625 μg·mL-1)浸泡10 s,晾干,放入生测瓶中,每瓶接3龄幼虫20头,每个处理重复3次,放置25℃养虫室中,48 h后检查幼虫死亡情况,所得数据用POLO软件分析,计算LC50。以Cry1Ac蛋白为阳性对照,灭菌水为阴性对照。
2 结果与分析
2.1 Bt LS-R-21菌株生物学特性的分析
Bt LS-R-21菌株半胞晶体形态的观察见图1。
由图1可知,Bt LS-R-21在LB培养基中30℃培养48 h可见菌落呈乳白色,表面粗糙不透明,边缘不整齐。挑取单菌落少量,制片,油镜下观察,可见伴饱晶体形态为大小不一的菱形。
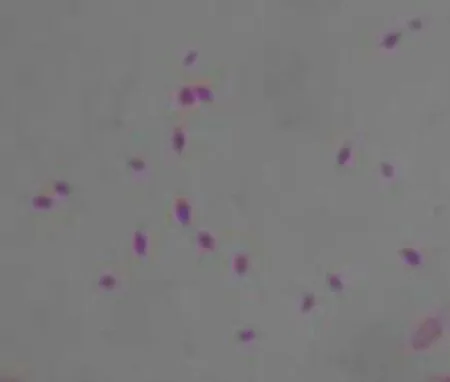
图1 Bt LS-R-21菌株的晶体形态Fig.1 Shape of crystal of Bt LS-R-21
2.2 Bt LS-R-21菌株cry基因类型的鉴定
对菌株LS-R-21进行基因型的PCR-RFLP鉴定,用cry1类基因鉴定引物K5un2/K3un2获得了大小为1.6 kb的PCR产物,将其进行PstⅠ/XbaⅠ双酶切分析,获得了RFLP图谱(见图2),从此图可以看出,扩增获得的1.6 kb PCR产物经酶切后产生了2个酶切片段,大小分别为1 117和518 bp,根据Kuo等表明[10],LS-R-21中含有cry1Aa基因。
2.3 cry1Aa全长基因的克隆及原核表达载体的构建
设计cry1Aa类基因的全长引物L5unx/L3unx,扩增得到cry1Aa3.6 kb的全长基因,将其与载体pEB进行连接,转化入感受态JM109中,经抗性筛选、PCR鉴定及SmaⅠ和SalⅠ双酶切分析,筛选出含有cry1Aa基因的阳性重组质粒。图3为阳性重组质粒酶切分析及PCR鉴定结果,经SmaⅠ和SalⅠ酶切后,得到大小为5.7 kb的载体条带和3.6 kb全长基因条带,说明该表达载体构建正确,将其命名为pEB-cry1Aa。

图2 Bt LS-R-21菌株cry1类基因的PCR鉴定与PCR-RFLP酶切检测电泳Fig.2 PCR detection and PCR-RFLP patterns of cry1 from Bt LS-R-21

图3 pEB-cry1Aa的酶切分析与PCR鉴定Fig.3 Restriction analysis and PCR detection of pEB-cry1Aa
2.4 cry1Aa基因的序列测定及分析
通过DNAMAN软件分析cry1Aa测序结果,推导的氨基酸结果如图4所示。
由DNA序列推导的氨基酸为1 175个,由该蛋白质的氨基酸组成分析表明亮氨酸(Leu)、谷氨酸(Glu)、丝氨酸(Ser)三种氨基酸含量最高,分别占总数的8.68%、7.74%、7.32%,。蛋白质的分子质量为132.9964 ku,等电点为4.805,此蛋白为弱酸性蛋白质。利用ANTHEPROT6.0软件,采用Garnier方法,由此氨基酸组成推导出Cry1Aa19的二级结构,H代表螺旋,S代表折叠,T代表转角,C代表其他松散结构,它们所占比例分别为25.7%、11.6%、36.10%和26.56%。该基因已在国际基因库GenBank中注册,登记号为HQ685121,并经Btδ-内毒素基因国际命名委员会正式命名为cry1Aa19。


图4 通过DANMAN软件分析推导的氨基酸结果Fig.4 Amino acid results by DNAMAN software
NCBI Blast比对该蛋白氨基酸序列与其他cry1Aa基因氨基酸的差异,通过vector NTI软件分析,结果表明,该蛋白与已公布的18种Cry1Aa蛋白的氨基酸序列多数存在差异,与Cry1Aa7差异最大,有33个氨基酸残基不同,与其同源性仅为97.19%。2010年国际Bt杀虫晶体蛋白命名委员会将其命名为Cry1Aa19。cry1Aa19与其他cry1Aa基因比较,在结构域Ⅰ、结构域Ⅱ存在一些差异,结构域III与大部分cry1Aa基因相同。说明来自不同国家和地区的菌株所含的同一亚类基因保守性较强。同时NCBI Conserved Domain Summary分析结果表明该蛋白的Domain I由N端第36~254位,共219氨基酸组成,Domain II由第259~460位,共202个氨基酸组成,Domain III由第470~607位,共138个氨基酸组成。因此,Cry1Aa蛋白的活性区估计在N-端的36~607个氨基酸附近。因此菌株LS-R-21中含有的Cry1Aa蛋白具有进一步研究和开发的价值。
2.5 cry1Aa19基因在E.coli Rosetta(DE3)中的表达
将重组质粒pEB-cry1Aa转化到E.coliRosetta(DE3)中,IPTG诱导表达,SDS-PAGE(8%)凝胶电泳。结果表明,无论是在可溶还是在非可溶组分中cry1Aa19基因都能通过表达载体pEB在大肠杆菌中高效表达蛋白,而经IPTG诱导的转入Rosetta(DE3)的pEB空载体没有特异的目的条带产生(见图5)。
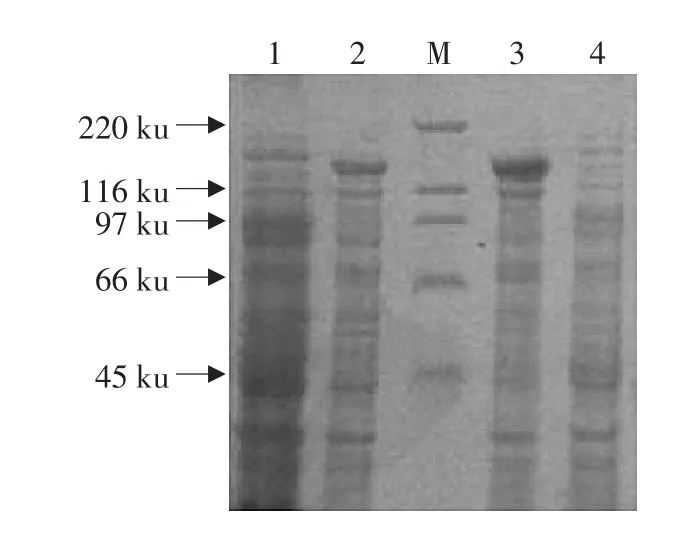
图5 cry1Aa19在Rosetta(DE3)中表达蛋白的SDS-PAGE分析Fig.5 Protein SDS-PAGE which cry1Aa19 expressed in Rosetta(DE3)
2.6 杀虫生物活性测定
Cry1Aa19 对小菜蛾的 LC50为 1.18 μg·mL-1,95%置信区间为0.97~1.42。阳性对照Cry1Ac对小菜蛾的LC50为4.92 μg·mL-1,95%置信区间为3.80~7.00。表明Cry1Aa19对小菜蛾有较强的杀虫活性。
3 讨论与结论
本实验室从黑龙江省五常市拉林镇附近分离到一株Bt菌株LS-R-21,通过显微镜下观察到LS-R-21的晶体呈现大量的大小不一的菱形,经PCR-RFLP体系对LS-R-21菌株进行基因型的鉴定,结果发现该菌株含有cry1Aa基因,其与已知的cry1Aa基因的相似性最高达99%。该蛋白与已公布的18种Cry1Aa蛋白的氨基酸序列多数存在差异,与Cry1Aa7差异最大,有33个氨基酸残基不同。蛋白质的分子质量为132.9964 ku,等电点为4.805,为弱酸性蛋白质。由氨基酸组成推导出Cry1Aa的二级结构表明,螺旋、折叠、转角及其他松散结构所占比例分别为25.7%、11.6%、36.10%和26.56%。cry1Aa19与其他cry1Aa基因比较,在结构域Ⅰ、结构域Ⅱ中存在一些差异,结构域Ⅲ与大部分cry1Aa基因相同。说明来自不同国家和地区的菌株所含的同一亚类基因保守性较强。
cry1Aa19基因在大肠杆菌中成功的表达了132 ku的重组蛋白。Cry1Aa蛋白对重要的农业害虫小菜蛾有较高的毒力,LC50为 1.18 μg·mL-1,95%置信区间为0.97~1.42。
本研究发现的cry1Aa19为我国抗虫转基因作物和工程菌的研制提供了新的基因来源。自转Bt基因作物产业化以来,其高效的杀虫性能和高度的特异性对其靶标害虫具有强大的选择压力,害虫Bt抗性的产生可能对这项高效无公害技术的长期效益构成威胁[14]。为延缓抗性的产生,分离出新的作用机制不同的杀虫蛋白是保持转基因作物成功的关键。因而开发cry1Ac类以外的基因受到人们的普遍关注。cry1Aa19基因的克隆和杀虫活性研究,将为害虫防治提供新的基因选择和研究思路。为筛选、延缓和克服昆虫产生抗性的基因组合提供了重要依据,具有重要的理论和实践意义。
[1] 戴莲韵,王学聘.苏云金芽胞杆菌研究进展[M].北京:科学出版社,1997.
[2] 中国植物保护学会,植物保护学学科发展报告,2007-2008[R].北京:中国科学技术出版社,2008:138-142.
[3] 李先军,高继国,齐东来,等.十株Bt野生菌杀虫晶体蛋白及基因型分析[J].东北农业大学学报,2009,40(9):57-61.
[4] Van Frankenhuyzen K.Application of Bacillus thuringiensis in forestry[C]//Charles J F,Dele'cluse A,Nielsen L R,Entomopathogenic bacteria:From laboratoryto field application.Dordecht:Kluwer Academic Publishers,2000:371.
[5] Bauce E,Carisey N,Dupont,A,et al.Bacillus thuringiensissubspkustakiaerial spray prescriptions for balsam fir stand protection against spruce budworm(Lepidoptera:Torticidae).[J]Econ.Entomol,2004,97:1624-1634.
[6] Crickmore N.Bacillus thuringiensistoxin nomenclature[2008-08-20].http://www.lifesc.i sussex.ac.uk/home/Neil_Crickmore/Bt.
[7] 邓淑,束长龙,林毅,等.新型cry7Ab基因的鉴定、克隆、表达与杀虫活性[J].农业生物技术学报,2009,17(5):908-913.
[8] 吴洪福,郭淑元,李海涛,等.苏云金芽孢杆菌杀虫晶体蛋白结构和功能研究进展[J].东北农业大学学报,2009,40(2):118-122.
[9] 宋福平,张杰,陈中义,等.苏云金芽孢杆菌cry基因PCR-RFLP鉴定体系的建立[J].中国农业科学,1998,31(3):8-13.
[10] Kuo W S,Chak K F.Identification of novel cry-type genes fromBacillus thuringiensisstrains on the basis of restriction fragment length polymorphism of the PCR-amplified DNA[J].Appl Environ Microbiol,1996,62(4):1369-1377.
[11] Song F,Zhang J,Gu A,et al.Identification of cry1I-type genes fromBacillus thuringiensisstrains and characterization of novel cry1I-type gene[J].Appl Environ Microbiol,2003,69(9):5207-5211.
[12] Yu H,Zhang J,Huang D,et al.Characterization ofBacillus thuringiensisstrain Bt185 toxic to the Asian cockchafer:Holotrichia parallela[J].Curr Microbiol,2006,53(1):13-17.
[13] Sambrook J,Russell D W.Molecular cloning[M].A Laboratory Manual,3nd.Cold Spring Harbor NY:Cold Spring Harbor Laboratory,2001.
[14] Andow D A,Hilbeck A.Science-based risk assessment for nontarget effects of transgenic crops[J].Bioscience,2004,54(7):637-649.
猜你喜欢
今日农业(2019年12期)2019-08-15
中国洗涤用品工业(2019年4期)2019-05-11
广州大学学报(自然科学版)(2019年1期)2019-05-07
中成药(2018年1期)2018-02-02
中成药(2017年3期)2017-05-17
中国中医药信息杂志(2016年7期)2016-12-01
天津科技大学学报(2016年1期)2016-02-28
湖北师范大学学报(自然科学版)(2015年2期)2016-01-10
动物医学进展(2015年10期)2015-12-07
川北医学院学报(2015年5期)2015-12-05
