
粟米草皂甙对血管平滑肌细胞增殖的影响及机制探讨
2010-05-22 12:05熊建军吴周环甘丽君黄爱君李雪芹
山东医药 2010年18期
熊建军,龚 帧,吴周环,林 玲,甘丽君,黄爱君,李雪芹
(九江学院基础医学院,江西九江 332000)
研究证实,血管平滑肌细胞(VSMCs)异常增殖是高血压的重要病理变化。VSMCs异常增殖使血管壁增厚,顺应性下降,管腔狭窄,同时分泌众多活性物质及细胞外基质成分,促进高血压的病理过程。因此,抑制 VSMCs增殖是防治高血压的重要途径之一[1]。粟米草为番杏科植物,性味淡平,有清热解毒、利湿功能,主治腹痛泻泄、疮疥肿毒等[2]。文献报道,粟米草中提取的粟米草皂甙,具有抗心律失常、增加冠状动脉血流量、降低血压等药理作用[3],但其降压机制尚不明确。2008年 10月 ~2009年 7月,我们观察了粟米草皂甙对 VSMCs增殖的影响,并初步探讨其作用机制。
1 材料与方法
1.1 材料 粟米草皂甙,原代培养大鼠胸主动脉VSMCs(第 4~8代细胞),DMEM干粉,新生小牛血清,胰酶,青霉素,链霉素,四甲基偶氮唑蓝(MTT),总 RNA提取试剂盒,一步法 RT-PCR试剂盒,β-actin及线粒体转录因子 A(mtTFA)引物,CO2细胞培养箱,酶标仪,普通台式离心机,电热恒温水浴箱,PCR仪,凝胶成像系统,超净工作台,普通及低温冰箱,普通及电子天平。
1.2 方法
1.2.1 VSMCs增殖情况观察 采用 MTT法。将VSMCs悬液以每孔 1×104个细胞接种于 96孔细胞培养板,待细胞贴壁生长良好后吸去原培养液,换含血清培养液继续培养 24 h。实验组加入粟米草皂甙,使其终浓度分别为 1、3、9、27μmol/L(其中 DMSO终浓度不超过 0.1%);对照组加等体积稀释后的 DMSO。每组均设 4个复孔。常规培养不换液,分别培养 24、48、72 h。实验终止前加 MTT(5 mg/ml)20μl,继续培养 4 h后离心弃上清液,加 DMSO 150μl,振荡 10 min,溶解结晶物。酶联免疫检测仪上波长 490 nm处测各孔吸光度值(A值),以空白对照调零,计算对细胞增殖抑制率。细胞增殖抑制率(%)=(1-观察组 A值 /空白组 A值)×100%。
1.2.2 VSMCs中 mtTFA mRNA表达检测 采用RT-PCR法。Trizol试剂提取 VSMCs总 RNA,分光光度计测总 RNA纯度。采用一步法 RT-PCR试剂盒,按照说明书操作,内参照为 β-actin(475 bp)。反应结束后,取 PCR反应液 8μl进行 1.4%的琼脂糖凝胶电泳,确认 PCR扩增产物。以目的条带值与β-actin条带的平均光密度值比值表示 mtTFA mRNA表达量。
1.2.3 统计学方法 采用 SPSS15.0软件处理。组间显著性检验采用 t检验。P≤0.05为差异有统计学意义。
2 结果
2.1 两组细胞增殖抑制率比较 见表1。由表可见,在粟米草皂甙浓度为 3~27μmol/L范围内,细胞增殖抑制率与药物浓度呈明显依赖关系;且在一定浓度范围内,粟米草皂甙对细胞增殖的抑制作用呈时间依赖关系。
2.2 两组 mtTFA mRNA表达比较 见表2。由表可见,不同浓度粟米草皂甙作用后,VSMCs中 mt-TFA mRNA表达水平下降,随浓度递增下降更为明显。
表1 两组不同时间点细胞增殖抑制率比较(%,±s)
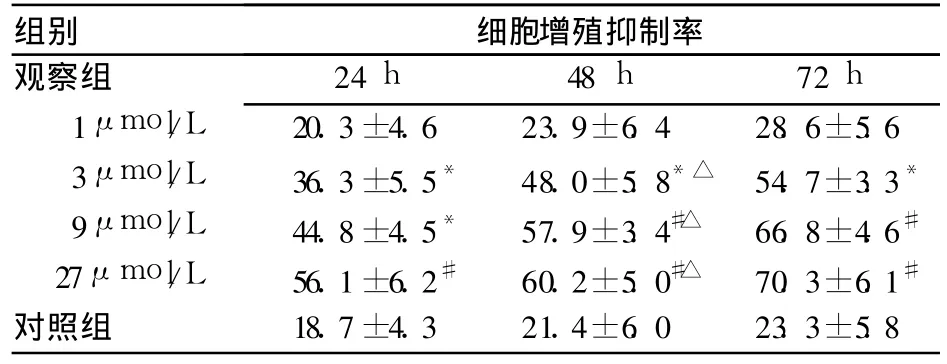
表1 两组不同时间点细胞增殖抑制率比较(%,±s)
注:与对照组比较,*P<0.05,#P<0.01;与同组 24 h、72 h比较,△P<0.05
组别 细胞增殖抑制率观察组24 h 48 h 72 h 1μmol/L 20.3±4.6 23.9±6.4 28.6±5.6 3μmol/L 36.3±5.5* 48.0±5.8*△ 54.7±3.3*9μmol/L 44.8±4.5* 57.9±3.4#△ 66.8±4.6#27μmol/L 56.1±6.2# 60.2±5.0#△ 70.3±6.1#对照组 18.7±4.3 21.4±6.0 23.3±5.8
表2 两组 mtTFA mRNA表达比较(±s)
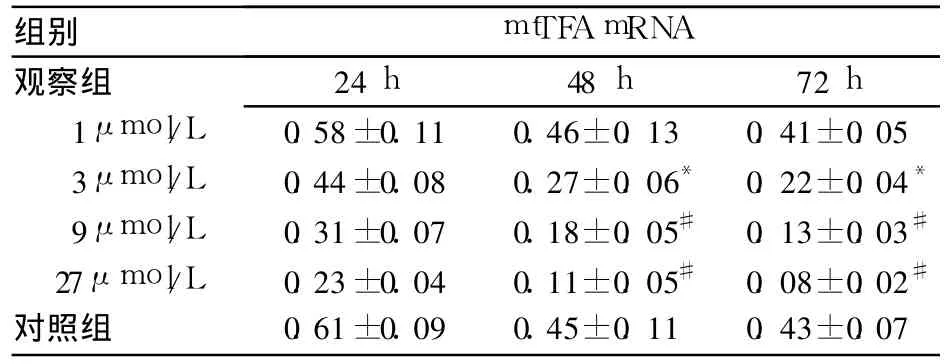
表2 两组 mtTFA mRNA表达比较(±s)
注:与对照组比较,*P<0.05,#P<0.01
组别 mtTFA mRNA观察组24 h 48 h 72 h 1μmol/L 0.58±0.11 0.46±0.13 0.41±0.05 3μmol/L 0.44±0.08 0.27±0.06* 0.22±0.04*9μmol/L 0.31±0.07 0.18±0.05# 0.13±0.03#27μmol/L 0.23±0.04 0.11±0.05# 0.08±0.02#对照组 0.61±0.09 0.45±0.11 0.43±0.07
3 讨论
粟米草为番杏科植物,包括星粟米草属和粟米草属,有清热解毒、利湿等功能,其药效物质基础是黄酮类和甾体类化学成分。我们的前期研究表明,非细胞毒剂量的粟米草提取物对高血压大鼠有明显的降压效果,但是其作用机制尚不明确。
在动脉粥样硬化、高血压等疾病中,VSMCs异常增殖是重要的病理变化,抑制 VSMCs增殖与迁移可逆转血管构型,提高动脉粥样硬化、高血压等患者的生存率及生存质量。目前,临床上只有少数药物如血管紧张素转化酶抑制剂、血管紧张素Ⅱ受体拮抗剂等有类似疗效。本研究结果显示,不同浓度粟米草皂甙作用后均可抑制 VSMCs增殖,并且这种抑制作用随着粟米草皂甙浓度增高而增强。这表明粟米草皂甙可有效抑制 VSMCs增殖,为阐明粟米草抗高血压的分子机制提供了理论基础。
在体外培养中,血清对 VSMC增殖受细胞外和细胞内信号网络的调控,其中涉及多条通路。我们研究发现,粟米草皂甙抑制 VSMCs的增殖与 mtTFA的转录表达存在关联。已知 mtTFA是由核基因组编码,在胞质内合成后再进入线粒体的蛋白分子,对线粒体增殖和功能起重要调控作用[4]。mtTFA从胞质转位到线粒体,作用于 mtDNA启动基因,在线粒体 RNA聚合酶作用下,通过调节 mtDNA轻链和重链的转录而控制 mtDNA的转录和复制。因此,mtTFA是决定线粒体数量和转录活性的关键分子,对细胞增殖产生影响[5]。国外研究已证明,通过反义技术干预 mtTFA能显著抑制平滑肌细胞的增殖[6]。本研究结果显示,粟米草皂甙能抑制血清对mtTFA mRNA的表达,提示粟米草皂甙可能通过抑制 mtTFA mRNA高表达发挥抑制VSMCs增殖作用。已知 mtTFA表达受控于转录水平的调控,多种胞外信号如葡萄糖、LPS等可导致其启动子的激活[7,8],引起蛋白表达升高。mtTFA启动子的激活依赖于核呼吸因子 1和 2(NRF-1、NRF-2)[9],同时过氧化物酶体增殖物激活受体 γ辅激活因子-1(PGC-1)对NRF-1、NRF-2和 mtTFA等基因表达产生强大的诱导作用[10]。粟米草皂甙中的黄酮类和甾体类化学成分有可能在多个环节抑制 mtTFA的转录,从而抑制了 mtTFA蛋白的表达,最终导致线粒体功能改变,细胞增殖减少,其具体机制有待深入研究。
[1]Voelkel NF,Tuder RM.Cellular and molecular biology of vascular smooth muscle cells in pulmonary hypertension[J].Pulm Pharmacol Ther,1997,10(5-6):231-241.
[2]姚果原,周凯旋,卢琦华,等.粟米草总皂甙的心血管药理作用[J].九江医学,1994,9(1):22-23.
[3]李雪芹,高春华,刘建云,等.粟米草提取物对自发性高血压大鼠的降压作用[J].山东医药,2005,45(17):8-9.
[4]Reyes A,Mezzina M,Gadaleta G.Human mitochondrial transcription factor A(mt TFA):gene structure and characterization of related pseudogenes[J].Gene,2002,291(1-2):223-232.
[5]Freyssent D,Dicarlo M,Escobar P,et al.Zidovudine(AZT)induced alterations in mitochondrial biogenesis in rat striated muscles[J].Can JPhysiol Pharmacol,1999,77(1):29-35.
[6]Inagaki H,Kitano S,Lin KH,et al.Inhibition of mitochondrial gene expression byantisense RNA of mitochondrial transcription factor A(mtTFA)[J].Biochem Mol Biol Int,1998,45(3):567-573.
[7]Kain KH,Popov VL,Herzog NK.Alterations in mitochondria and mtTFA in response to LPS-induced differentiation of B-cells[J].Biochim Biophys Acta,2000,1494(1-2):91-103.
[8]Kanazawa A,Nishio Y,Kashiwagi A,et al.Reduced activity of mt-TFA decreases the transcription in mitochondria isolated from diabetic rat heart[J].Am J Physiol Endocrinol Metab,2002,282(4):778-785.
[9]Kraft CS,Lemoine CM,Lyons CN,et al.Control of mitochondrial biogenesis during myogenesis[J].Am JPhysiol Cell Physiol,2006,290(4):1119-1127.
[10]Anedersson U,Scarpulla RC.Pgc-1-related coactivator,a novel,serum-inducible coactivator of nuclear respiratory factor 1-dependent transcription in mammalian cells[J].Mol Cell Biol,2001,21(11):3738-3749.
猜你喜欢
现代农村科技(2022年1期)2022-01-21
中国循证心血管医学杂志(2021年10期)2021-11-05
情感读本·道德篇(2019年8期)2019-10-21
食品工业科技(2019年3期)2019-03-28
台州学院学报(2018年6期)2018-02-26
医学研究杂志(2015年5期)2015-06-10
资源节约与环保(2014年12期)2014-02-01
读者·校园版(2013年14期)2013-05-14
天然产物研究与开发(2010年5期)2010-10-09
食品工业科技(2010年8期)2010-09-12
