
产磷脂酶 C芽孢杆菌 Z-13产酶条件优化
2010-05-17 00:43詹逸舒孙天玮刘仁萍吴永尧
天然产物研究与开发 2010年3期
詹逸舒,孙天玮,陈 里,刘仁萍,吴永尧
湖南农业大学生物科学技术学院,长沙 410128
磷脂酶 C(phospholipase C,PLC)是一类可催化磷酯 C3上的磷酯酰键的脂类水解酶,在细胞代谢、信息传递和抗血小板功能等方面起着重要作用[1]。其作用底物为甘油磷脂和磷脂酰类,能特异性的水解质膜表面鞘磷脂(sphingomyelin,SM)的磷酸二酯键产生神经酰胺[2,3](ceramide)。神经酰胺作为生物信息传递的“第二信使”,具有调节神经细胞生长,诱导癌细胞程序性凋亡,以及对皮肤的特殊生物高效保湿功能[4]。国外已将其作为重要生理活性成分及医药原料,广泛应用于在药品、化妆品、美容食品等领域[5]。
目前国外主要从动植物原料中提取神经酰胺,植物源比动物源神经酰胺更为安全。神经酰胺在生物材料中含量非常低(万分之一至十万分之一),价格十分昂贵。我国魔芋(Konjac)资源十分丰富,占世界魔芋种数的 22.6%。其中含有丰富的神经酰胺(0.15% ~0.2%),是米糠和小麦中(0.01% ~0.02%)的 10倍[6],但其中的神经酰胺与磷酸胆碱结合以鞘磷脂形式存在于细胞膜中,而呈结合状态的神经酰胺难以直接提取。为使可提取的游离神经酰胺含量提高,积极开发产生特殊活性的 PLC的菌株,具有十分重要的意义。作者对产磷脂酶 C芽孢杆菌 Z-13产酶条件进行了研究,目的在于优化产酶条件,提高产酶能力,为磷酯酶 C的开发利用及魔芋神经酰胺的提取打下基础。
1 材料与方法
1.1 菌种及培养基
产磷脂酶 C芽孢杆菌 Z-13由本实验室分离、筛选并保藏。
基础培养基:蛋白胨 1.0%、牛肉膏 0.3%、NaCl 0.5%、pH 7.2~7.4,1×105 Pa灭菌 30min。
硼砂卵黄培养基:硼砂 0.19% 、硼酸 1.09%、NaCl 0.66% 、卵黄 10%,pH 7.2~7.4。
1.2 产磷脂酶 C菌 Z-13产酶条件的优化
1.2.1 培养基组成的确定
保持基础培养基其它组成不变,依次改变培养基的氮源、碳源和无机盐种类,等量接种 Z-13菌悬液,35℃、装液量为 100mL/250m L、150 r/min摇床培养 12 h,测定发酵液中磷脂酶 C活性及生物量。
确定最佳氮源、碳源和无机盐后,采用均匀设计试验[7],利用 DPS v7.05专业版软件的均匀试验设计表,根据预试验结果及试验可行性确定考察因素的范围,对筛选出的培养基组分进行优化,然后通过二次多项式逐步回归分析实验数据,建立回归方程,通过指标最大预测值确定发酵培养基各组分的适合配比,并进行验证试验。
1.2.2 产酶培养条件的确定
采用优化后的培养基,分别从接种量、摇床转速、培养基初始 pH值、培养温度、培养时间等进行单因素试验。
1.3 粗酶液制备
培养 12 h的发酵液用 60目纱布粗滤,8000 r/min离心 25min,收集上清液即为粗酶液,供酶活测定。
1.4 生物量测定
平板菌落计数法
1.5 酶活测定
卵黄琼脂杯碟法[8,9]:取 20 g卵黄(新鲜的土鸡蛋),加 80mL无菌水,摇匀,得到 20%的卵黄液,再制成硼砂卵黄琼脂培养基。待培养基溶化后,倒入培养皿中冷却凝固,其上平放牛津杯,吸取 100μL粗酶液于杯中,37℃条件下培养 24 h,根据磷脂酶C能水解卵黄琼脂平板中卵磷脂产生乳白色的晕圈,用游标卡尺测定反应后生成的乳白色晕圈(沉淀圈)直径,乳白色晕圈越大,PLC活性越大。
2 结果与讨论
2.1 单因素法优化培养基组成
2.1.1 碳源
基础培养基中分别添加浓度为 1.0%的不同碳源,它们对磷脂酶 C活力及生物量影响见表 1。从表中可以看出,可溶性淀粉和魔芋飞粉为碳源时菌Z-13产酶能力明显优于牛肉膏、葡萄糖、蔗糖、麸皮、玉米粉,说明菌 Z-13利用可溶性淀粉和魔芋飞粉的能力优于其他几种碳源。其中魔芋飞粉作为碳源时,对其产酶影响较大,乳白色沉淀圈直径达到了23mm,生物量为 4.5×107CFU/mL,这可能是由于魔芋飞粉中含有的丰富淀粉和鞘磷脂化合物,及其它有利于其生长的营养成分,促进了其发酵产酶,因此选择魔芋飞粉为碳源。

表 1 不同碳源对酶活和生物量的影响Table 1 Effect of different carbon sources on enzyme activity and biomass
2.1.2 氮源
在基础培养基中分别添加浓度为 1.5%的不同氮源,它们对磷脂酶 C活力及生物量的影响见表 2。从图中可以看出,酵母膏作氮源时发酵后产磷脂酶C活性最高,乳白色沉淀圈直径达到了 24mm。这说明菌 Z-13利用酵母膏的能力优于其他几种氮源。这可能与酵母粉含有的氨基酸、维生素等生长因子的配比更适合其的生长。其中黄豆粉作氮源时,乳白色沉淀圈直径达到了 22mm,略低于酵母膏,但明显高于其他 2种无机氮源,且生物量为 1.6×108CFU/mL,高于其他氮源。由于黄豆粉是发酵工业最常用的有机氮源,价格便宜,而且其中含有的丰富磷脂类化合物能诱导该菌种产酶,综合考虑效果与原料成本,因此选择黄豆粉为氮素营养。
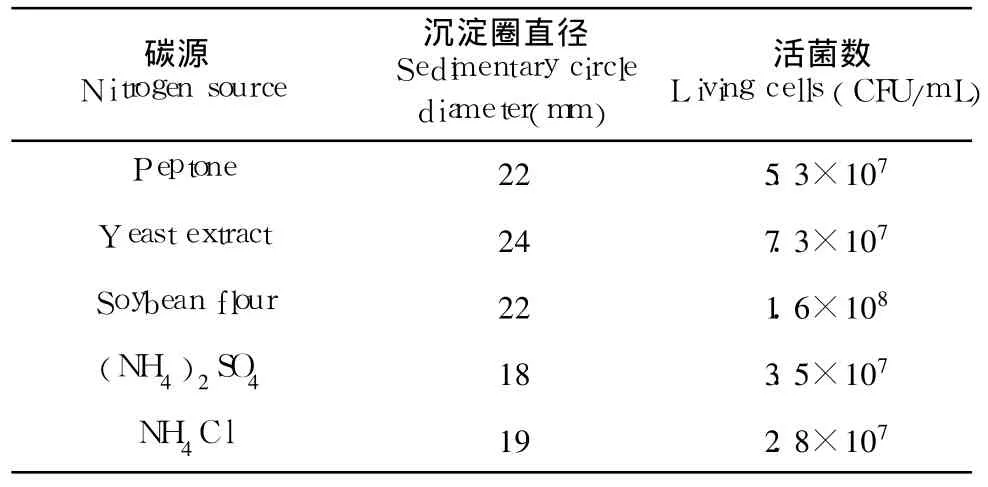
表 2 不同氮源对酶活和生物量的影响Table 2 Effect of different nitrogen sources on enzyme activity and biomass
2.1.3 无机盐
在基础培养基中分别添加浓度为 0.2%的不同无机盐,它们对磷脂酶 C活力及生物量的影响见表3。结果表明,Zn2SO4◦7H2O和 K2HPO4◦3H2O有利于菌种生长,对产酶有一定的促进作用,产生的乳白色沉淀圈直径大于其他无机盐,生物量也高于其他无机盐。可能由于金属离子中 Zn2+是酶的辅酶或激活剂[10];同时磷是氧化磷酸化反应的必需元素,K+能影响细胞膜的通透性,磷酸盐既能促进菌体的基础代谢,又影响许多代谢产物的生物合成,因此选用 Zn2SO4◦7H2O和 K2HPO4◦3H2O为无机盐组合,添加到培养基中进行下一步优化试验。

表3 不同无机盐对酶活和生物量的影响Table 3 Effect of different inorganic salts on enzyme activity and biomass
2.2 均匀设计法优化培养基
培养基中各组分之间有一定的内在联系,在单因素试验的基础上,考察培养基中碳源、氮源及无机盐的含量对菌种生长及产磷脂酶 C的影响,以寻求培养基中各组分的最佳配比。对筛选出的 4种培养基组分进行了优化,采用 4因素 15水平 15组试验的方式,每 1 L培养基内各因素水平为:魔芋飞粉和黄豆粉 0.0~35 g,各水平差为 2.5 g;K2HPO4◦3H2O和 ZnSO4◦7H2O 0.0~2.8 g,各水平差为 0.2 g。均匀设计表及试验结果见表 4。
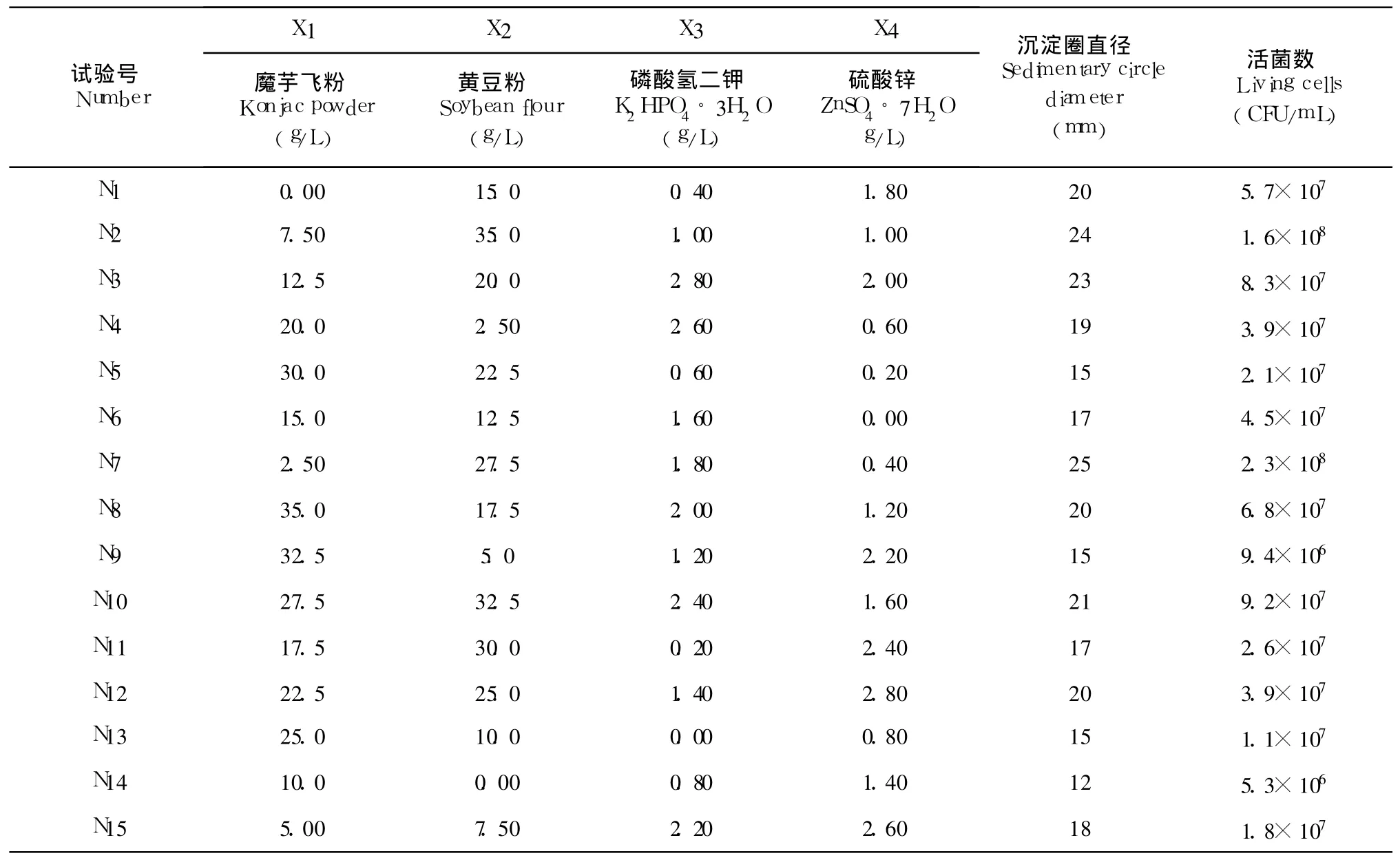
表 4 均匀设计试验优化培养基方案和结果Table 4 The scheme and resultofmedium optimization by uniform design
采用 DPS v7.05专业版软件,将表 4中各因素各水平对酶活的影响结果,进行二次多项式逐步回归分析,最终得到如下方程:
y=13.06-2.59X1+3.92X2+23.03X3+21.42X4+0.39X21-0.50X22-4.52X23-84.94X2
4
相关系数 R=0.9282,检验值 F=4.6703,显著水平 p=0.0382,剩余标准差 S=2.0874,调整后的相关系数 Ra=0.8229查表 F0.05(4,10)=3.48,F>F0.05(4,10),显著性水平检验通过,回归方程显著。
根据回归方程对 y求最大值,当 x1=2.5 g/L,x2=30 g/L,x3=2.8 g/L,x4=1.3 g/L时,Ymax=27.75 mm,其预测区间为:Y=Ymax±Uα◦ S=27.75 ±4.09。
即以魔芋飞粉 2.5 g/L、黄豆粉 30 g/L、K2HPO4◦ 3H2O 2.8 g/L、Zn2SO4◦ 7H2O 1.3 g/L为优化培养基配方时,通过卵黄琼脂杯碟法测得产生的乳白色晕圈(沉淀圈)直径应该在 23.66~31.84 mm范围内。根据优化条件,取魔芋飞粉 2.5 g/L、黄豆粉30 g/L、K2HPO4◦ 3H2O 2.8 g/L、Zn2SO4◦ 7H2O 1.3 g/L,进行 3次摇瓶实验验证,最终乳白色晕圈(沉淀圈)直径的平均结果为 27mm,生物量为 5.7×108CFU/m L,实验结果在预测的范围内,且均有显著性提高。
2.3 产酶条件确定
2.3.1 接种量优化
在相同培养条件前提下,分别按 0.1%、0.5%、1%、4%和 8%的接种量接种菌悬液(菌浓度 3×105 CFU/mL)于培养基中进行培养,它们对磷脂酶 C活力及生物量的影响见图 1。结果表明,接种量为0.5%时,晕圈直径为 26 mm,是 5个不同梯度接种量中晕圈最大的;接种量为 1.0%时生物量达到最大,晕圈直径为 24.5mm,酶活下降不大。当接种量逐渐继续增大时,酶活反而减小,这可能是由于随接种量的增加,菌体繁殖过快、过稠,造成营养基质缺乏或溶氧不足而不利于菌体产 PLC。基于磷脂酶 C活力及生物量综合考虑,选择接种量为 1.0%的为该菌培养的合适接种量。

图1 接种量对酶活和生物量的影响Fig.1 Effects of inoculation size on enzymeactivity and biomass
2.3.2 转速优化
分别在摇床转速为 60 r/min、100 r/min、150 r/min和 200 r/min的条件下进行摇瓶培养,摇床转速与酶活及生物量的关系见图 2。结果表明,在较低的转速下,溶解氧浓度较低,所得到的生物量较小,发酵液的酶活力较低;转速为 150 r/min时,乳白色晕圈直径最大,且生物量也达到最大;摇床转速在200~250 r/min之间,由于转速过快,剪切力过大,不利于菌种生长,发酵产 PLC能力略有下降。基于提高溶氧,菌种的耐受力及节约能源方面的考虑,选择摇床转速150 r/min为该菌培养的合适摇床转速。

图 2 摇床转速对酶活和生物量的影响Fig.2 Effects of rotating speed on enzyme activity and biomass
2.3.3 温度优化

图3 温度对酶活和生物量的影响Fig.3 Effects of temperature on enzyme activity and biomass
温度可通过影响发酵过程中酶反应速率、蛋白质性质及发酵液物理性质,进而影响发酵的动力学特性和产物的合成。分别按 25、28、32、35、40℃摇瓶培养,培养温度与酶活及生物量的关系见图 3。结果表明,在 25~36℃的温度范围内,菌体生物量随温度增加而不断增加,同时酶活力也在 32℃左右达到最大;当温度继续升高,不利于菌 Z-13生长和PLC积累,生物量及酶活力均呈下降趋势。因此,选择温度为 32℃为该菌合适的培养温度。

图 4 pH对酶活和生物量的影响Fig.4 Effects of pH on enzyme activity and biomass
2.3.4 初始 pH优化
分别按培养基初始 pH 5、6、7、8、9、10和 11摇瓶培养,培养基初始 pH与生物量及酶活的关系见图 4。结果表明,当培养基初始 pH小于 10时,酶活随 pH提高而增大,菌体生物量在 pH值为 8时达到最大,且 pH在 8~10范围内,生物量变化不大;当pH值为 11时,酶活开始显著下降,菌体生物量与产PLC酶活的培养基合适初始 pH都在碱性范围内。基于菌种生长和 PLC的积累方面的考虑,选择pH9.0为该菌发酵合适的初始培养基 pH。
2.3.5 培养时间优化

图 5 培养时间对酶活和生物量的影响Fig.5 Effects of time on enzyme activity and biomass
微生物发酵终点的判断,对提高产物的生产能力和经济效益很重要的,因此确定一个合适的培养时间十分必要,培养时间与生物量及酶活的关系见图 5。结果表明,随着培养时间的延长,菌体的生长繁殖,酶活开始有明显的上升,培养 15 h后,酶活达到最大值,18~21 h菌体达到对数生长期。随着时间的推移,营养物质不断减少,组成成分发生变化,菌体的生长繁殖速度下降;同时由于菌体释放体内的分解酶及各种环境因子的改变抑制了酶的活性。因此,发酵培养时间不宜过长,最佳培养时间为15 h。
2.3.6 产酶条件优化前后 PLC酶活定性测定
利用磷脂酶 C可水解卵黄中的卵磷脂,在含有卵黄的琼脂平板上产生乳白色晕圈的特点(如图 6所示),可以根据酶样品在卵黄平板上所产生的乳白色晕圈的大小来定性检测菌种产磷脂酶 C酶活的高低。由于操作方便,原料丰富,广泛的应用于磷脂酶 C菌种的筛选及磷脂酶 C的纯化制备[11,12]。将菌 Z-13接种至产酶优化前后培养基中,分别在优化前后的条件进行培养,卵黄琼脂杯碟法定性测定PLC酶活,结果如图 6所示。结果表明,在原始条件下,菌 Z-13产磷脂酶 C产生的沉淀圈(乳白色晕圈)直径为 22 mm,经过产酶条件优化后,菌 Z-13产磷脂酶 C产生的沉淀圈(乳白色晕圈)直径为 30 mm,增加了 8mm,产磷脂酶 C活性显著性提高。
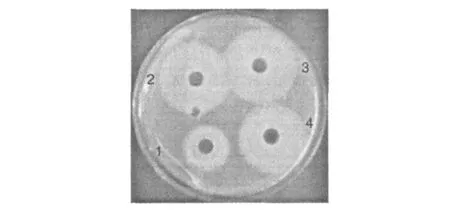
图 6 卵黄琼脂杯碟法酶活定性检测Fig.6 Qualitative detection ofenzyme activity by egg yolk cylinder platemethod
3 小结
目前国内还没有商业化的工业用 PLC制剂生产,对于细菌产磷脂酶 C的产酶条件研究研究报道还较少。本文以生物量和酶活为主要指标,采用单因素优化方法结合均匀设计试验,对芽孢杆菌菌 Z-13产磷脂酶 C(phospholipase C,PLC)条件进行了优化,确定产酶培养基的适宜碳源为魔芋飞粉、适宜氮源为黄豆粉,适宜无机盐为磷酸氢二钾和硫酸锌,采用均匀优化试验设计,确定培养基组成为魔芋飞粉2.5 g/L、黄豆粉 30 g/L、K2HPO4◦ 3H2O 2.8 g/L、Zn2SO4◦7H2O 1.3 g/L;在培养条件为接种量为1.0%、摇床转速为 150 r/min、培养温度 30℃、初始pH值 9.0、培养 15 h,有利于菌 Z-13生长及产磷脂酶 C,活菌总数为 4.5×109CFU/m L,卵黄琼脂杯碟法测定磷脂酶 C产生的沉淀圈(乳白色晕圈)直径可达 30mm左右。本研究所获得芽孢杆菌 Z-13的优化产酶条件,经逐级放大试验,将有可能在解决魔芋鞘磷脂酶法水解产神经酰胺过程中提供生物催化剂,使可提取的游离神经酰胺含量提高,为磷酯酶 C的工业生物技术开发利用及魔芋神经酰胺的生物加工过程打下基础。
1 Chen T(陈涛),Wang CG(王常高),He DP(何东平).The mutation of high phospholipase C productivity strain.Nat Prod Res Dev(天然产物研究与开发)2005,17:712-716.
2 Hou LL(侯丽莉),Chen XH(陈肖华).Property and function of acid sphingomyelinase.Int J Radiat Med Nucl Med(国外医学:放射医学核医学分册),2005,25:44-47.
3 Yang D(杨栋),Yang LX(杨丽霞).The role of ceramide in apoptosis.J Int Pathology Clini Med(国际病理科学与临床杂志),2006,26:166-168.
4 Kong CZ(孔垂泽),Wang X(王侠).神经酰胺对人膀胱癌细胞凋亡诱导作用及其机制.Chin JExp Surgery(中华实验外科杂志),2006,10:1159-1161.
5 Sun QJ(孙庆杰).Research and development of natural ceram ide.China Oils and fats(中国油脂),2003,2:60-61.
6 Pu YF(蒲云峰),Zhang WM(张伟敏),Zhong G(钟耕).Application and function of ceramide.JCereals&Oils(粮食与油脂),2005,7:14-16.
7 Zhen XT(郑晓婷),Zhao XZ(赵新淮).Optimization of fermentation conditions for proteases p roduced by mucor.Inst Microbiology(微生物学通报),2009,36:193-197.
8 Schm iel DH,Miller VL.Bacterial phospholipase and pathagensis.Micral Infect,1999,1:1103-1112.
9 Gao L(高林),Chen T(陈涛),Gao ZM(高智谋).Op timal conditions of Bacilus cereus Shenzhen 754-1 strain with high phospholipase C production.JAnhui Agric Univ(安徽农业大学学报),2007,34:501-504.
10 Kook-Hwa Seo,Jong Il Rhee.H igh-level expression of recombinantphospholipase C from Bacilluscereusin Pichia pastoris and its characterization.Biotechnology Letter,2004,26:1475-1479.
11 Sun CL(孙春来),Gan X(干信).Studies on the purification of phospholipase C.JHubei Univ Tech(湖北工学院学报),2004,19(2):8-10.
12 Wang CG(王常高),Cheng MK(陈明锴),Cheng T(陈涛).Study on selection of strain with high production phospholipase C and its anti-platelet function.Nat Prod Res Dev(天然产物研究与开发),2003,15:345-348.
猜你喜欢
鸭绿江(2022年10期)2022-10-21
医学综述(2021年24期)2022-01-11
飞天(2021年12期)2021-12-24
昆明医科大学学报(2021年2期)2021-03-29
ELLE世界时装之苑(2020年8期)2020-08-03
中国心血管杂志(2020年2期)2020-05-15
安徽医科大学学报(2020年1期)2020-02-14
临床肝胆病杂志(2017年8期)2017-08-30
文艺生活·下旬刊(2017年2期)2017-03-18
天津医科大学学报(2015年2期)2015-12-22
