
草乌头光照适应性的转录组分析
2024-12-31 00:00:00史佳怡吕丽娟穆赢通路景诗张晓明王俊杰
草地学报 2024年7期
关键词:转录组












摘要:研究通过分析遮荫处理对草乌头(Aconitum kusnezoffii L.)叶片中差异表达基因的影响,揭示其响应遮荫的分子机制。通过对遮荫处理后草乌头叶片的转录组测序,发现数据质量较高(Q30≥94.37%),并通过拼接和组装得到59 082个Unigenes(N50为1 826 bp)。功能注释表明,有27 668个Unigenes(46.83%)被注释到多个重要数据库。差异表达基因分析筛选出483个差异基因,其中421个获得注释。研究发现,光照条件显著影响草乌头的光合作用、激素调控和能量代谢等生物过程。关键基因分析显示,LHCB17,ATPβ,PetA等在遮荫下表达下调,LHCB1,GA2ox等表达上调。这些基因的调控作用揭示了光照对草乌头生长和代谢适应性的影响。实时荧光定量分析验证了转录组数据的可靠性。研究结果不仅增进了对草乌头适应光照环境的理解,还为其遗传调控网络和功能基因的研究提供了线索,对草乌头的合理栽培和利用具有重要意义。
关键词:草乌头;转录组;遮荫;qRT-PCR
中图分类号:R282.2""" 文献标识码:A""""" 文章编号:1007-0435(2024)07-2018-10
doi:10.11733/j.issn.1007-0435.2024.07.003
引用格式:
史佳怡, 吕丽娟, 穆赢通,等.草乌头光照适应性的转录组分析[J].草地学报,2024,32(7):2018-2027
SHI Jia-yi, LYU Li-juan, MU Ying-tong,et al.Transcriptome Analysis of Light Adaptation of Aconitum kusenezoffii L.[J].Acta Agrestia Sinica,2024,32(7):2018-2027
收稿日期:2024-04-15;修回日期:2024-05-10
基金项目:草地多功能兼用型草种高效繁育与加工利用技术集成(2022YFDZ0025);六种特色蒙药材原生态种植关键技术研究与示范(2021GG0327)资助
作者简介:
史佳怡(1997-),女,汉族,内蒙古呼和浩特人,硕士研究生,主要从事牧草、药用植物种质资源与栽培育种方向研究,E-mail:1697720262@qq.com;*通信作者Author for correspondence,E-mail:jjw62@163.com
Transcriptome Analysis of Light Adaptation of Aconitum kusenezoffii L.
SHI Jia-yi1, LYU Li-juan2, MU Ying-tong1, LU Jing-shi1, ZHANG Xiao-ming1, WANG Jun-jie1*
(1.Inner Mongolia Autonomous Region Sino-Mongolia Medicinal Materials Breeding Engineering Technology Research Center,
Key Laboratory of Grassland Resources College of Grassland, Resources and Environment, Inner Mongolia Agricultural
University, Huhhot, Inner Mongolia 010018, China; 2.Hohhot Forestry and Grassland Bureau, Hohhot,
Inner Mongolia 010019, China)
Abstract:The effects of shading treatment on differentially expressed genes in leaves of Aconitum kusnezoffii were analyzed to reveal the molecular mechanism of its responses to shading. Through the transcriptome sequencing of the leaves of A. kusnezoffii after shading treatment,it was found that the data quality was high (Q30 ≥ 94.37 %),and 59 082 Unigenes (N50 = 1 826 bp) were obtained by splicing and assembly. Functional annotation showed that 27 668 Unigenes (46.83 %) were annotated to multiple important databases. Analysis of differentially expressed genes found 483 differentially expressed genes,of which 421 were annotated. It was found that light conditions significantly affected the biological processes such as photosynthesis,hormone regulation and energy metabolism of A. kusnezoffii. Key gene analysis showed that LHCB17,ATPβ and PetA were down-regulated under shading,LHCB1 and GA2ox were up-regulated. The regulation of these genes revealed the effect of light on the growth and metabolic adaptability of A. kusnezoffii. Real-time fluorescence quantitative analysis verified the reliability of transcriptome data.The results of this study not only enhance the understanding of the adaptation of A. kusnezoffii to low light environment,but also provide clues for the study of its genetic regulatory network and functional genes,which is of great significance for the rational cultivation and utilization of A. kusnezoffii.
Key words:Aconitum kusenezoffii L.;Transcriptome;Shade;qRT-PCR
草乌头(Aconitum kusnezoffii L.)为多年生草本植物,在我国东北和华北地区广泛分布[1]。草乌头的块根、叶片、花朵和芽部分都被用作药材。其中,干燥的块根是蒙药材的重要及常用的中药材之一。其具有祛风除湿、温经止痛的功效,常用于治疗关节疼痛、心腹冷痛以及麻醉止痛等症状[2-6]。
药用植物与其周围环境的相互作用对品质具有重要影响。光照作为影响药用植物生长、形态以及次生代谢的关键生态因素之一,在植物的生态适应过程中扮演着至关重要的角色。不同地区的光照条件受到多种因素影响,包括海拔、经纬度等,导致药用植物对光照的适应性和特征表现出明显的差异[7-8]。植物对光照的适应性不仅影响植物的生长发育,而且对于药用植物药材的品质和药效成分有一定影响作用[9]。光照作为调节植物生长和发育的关键因素,光量和光质的变化均对植物的光合作用产生影响,尤其在遮荫条件下,光照强度和光谱成分都会降低[10],包括降低红光与远红光的比例以及光合有效辐射等,这些因素均会限制植物的正常生长和发育过程[11]。因此,光照条件的变化对药用植物的品质具有直接的影响。目前,关于光照强度对阴生药用植物的影响已经进行了较多研究,但主要集中在最适光强和光质选择以及光照对药用植物活性成分诱导性效应等方面[12-13]。然而,光照如何调控药用植物的发育并影响其次生代谢合成的具体机制,目前仍不十分清晰。遮荫处理是模拟自然生长环境中光照条件变化的一种手段,可以有效模拟草乌头在不同生长环境下的生理生态响应。本研究通过对草乌头进行遮荫处理,可以探究草乌头对光照条件的适应机制,同时用转录组测序技术解析不同光照强度对草乌头叶片在光调节下的生长发育规律和内在代谢机制影响,以期为阐明光照对药用植物代谢积累机制奠定基础,同时为草乌头的精准栽培和合理利用提供科学依据和理论指导。
1" 材料与方法
1.1" 试验材料与处理
试验选用实验室低温贮藏两年以上的草乌头种子,将种子外表的翅搓掉后,用10%的NaClO溶液进行消毒,在1 800 mg·L-1的赤霉素中浸泡48 h。在培养皿中铺两层滤纸后将种子均匀摊放在上面,喷洒适量的蒸馏水,在10℃~20℃的变温培养箱中进行催芽。种子干燥时适时喷水,待种子发芽或长出子叶后点播入花盆中,胚根朝下,覆土3 cm左右。待长出真叶后,选取生长状况良好的实生苗,用相同材质及孔径的网袋进行遮荫处理。设置三个处理组:无遮荫,光照强度为790 lx(A组);覆盖一层遮阳网袋,光照强度为620 lx(B组)覆盖两层遮阳网袋,光照强度为450 lx(C组)。实验期间进行统一的水肥管理。
1.2" 转录组学测序及分析
1.2.1" RNA提取及质量控制" 据以上试验的研究结果,选取A组(无遮荫)、B组(覆盖一层遮阳网袋)和C组(覆盖两层遮阳网袋)生长的草乌头叶片,各称取新鲜叶片0.20 g,设置3个生物学重复,将样品用锡纸包好后,液氮速冻,经干冰保存送至百迈客生物科技有限公司进行测序。
1.2.2" 转录组分析流程" 对原始数据(Raw Data)进行质控,得到高质量Clean Data,使用Trinity软件采用paired-end方法对cleandata进行无参转录组从头组装(de novo)得到转录本,为了减少冗余信息,挑选最长的转录本作为Unigene,并使用CD-HIT[14]软件去冗余得到最终的Unigene,作为后续分析的参考序列。使用Diamond[15]软件将Unigene比对Nr,Nt,KOG,COG,Swiss-Prot,KO和GO数据库,以及使用HMMER[14]软件对Pfam数据库进行Unigene的功能分析。使用Express[17]软件对Unigene进行定量分析,筛选差异的条件为qlt;0.05且差异倍数(Fold Change)gt;2定为差异表达基因[18]。
1.3" 差异表达基因qRT-PCR验证
根据叶片转录组数据选取LHCB17,LHCB1,ATPβ,PetA,PFK,NCED,PP2C,GA2ox,DELLA和AMY验证差异表达基因结果(表1)。
2" 结果与分析
2.1" 测序数据质量检测与优化
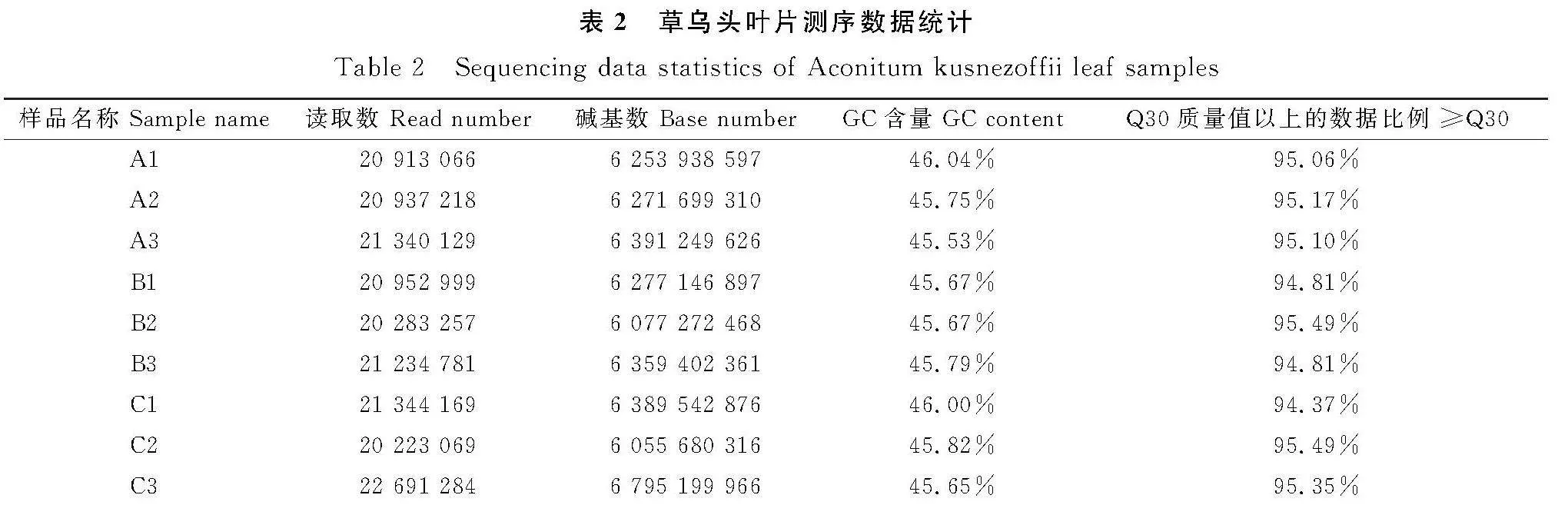
对草乌头叶片进行测序,获得9个样品的转录组原始数据,并通过对原始数据进行过滤筛选,获得56.78 Gb的高质量序列。经分析,所有产品的Q30≥94.37%,这表明样本测序数据和从头组装的质量很高,后续试验可以基于高质量的序列进行分析(表2)。
2.2" 无参转录组拼接结果统计
使用Trinity软件对样品的有效reads进行de novo拼接后,得到59 082个Unigenes,其中200~500 bp有28 681条,占比48.54%;大于1 000 bp序列有18 829条,占比31.87%;大于2 000 bp序列有8 472条,占比14.34%。拼接后总长度为58 562 027 bp,平均长度为991.20 bp,N50长度为1 826 bp(表3)。
2.3" Unigene的功能注释
草乌头叶片转录组测序得到Unigene功能注释见表4及图1。将样品59 082条Unigene序列分别与Nr,eggNOG,GO,Pfam,KEGG,SwissPort,KOG和COG数据库进行相似性比较,从而获得全面的基因功能信息。最终获得27 668个Unigenes,占总Unigenes的46.83%,长度大于1 000的序列有16 330条,占比59.02%。与Nr数据库比对,26 522条(44.89%)Unigenes获得注释;与eggNOG数据库比对,22 446条(37.99%)Unigenes获得注释;与GO数据库比对,21 638条(36.62%)Unigenes获得注释;与Pfam数据库比对,18 913条(32.01%)Unigenes获得注释;与KEGG数据库比对,17 099条(28.94%)Unigenes获得注释;与SwissPort数据库比对,16 819条(28.47%)Unigenes获得注释;与KOG数据库比对,14 353条(24.29%)Unigenes获得注释;与COG数据库比对,6 464条(10.94%)Unigenes获得注释。未被注释到的Unigenes有32 674条,占比55.30%。
2.4" 差异表达基因鉴定
对草乌头不同处理的样本进行差异表达基因筛选,将Fold Change≥2且FDRlt;0.01 作为筛选标准,对不同处理样本进行比较(图2),A组与B组比较共有219个差异基因(DEGs),其中55个DEGs上调表达,164个DEGs下调表达;A组与C组比较共有297个DEGs,其中58个DEGs上调表达,239个DEGs下调表达;B组与C组比较共有73个DEGs,其中37个DEGs上调表达,36个DEGs下调表达。
为了更直观区分DEGs的共同性和差异性,本研究将不同比较组的DEGs进行归类汇总(图3)。结果表明A组与B组比较特有DEGs有129个;A组与C组比较特有的DEGs有201个;B组与C组比较特有DEGs有47个。以A组为对照,B组和C组之间的重复DEGs有80个;以B组对照,A组和C组之间重复DEGs有10个;以C组为对照,A组和B组之间重复为16个。三者共有重复DEGs为0,三组共有483个DEGs。
2.5 "差异表达基因功能注释
对草乌头不同处理叶片转录组的差异表达基因进行数据库功能注释(表5),共注释到差异表达基因421个,占比87.16%。其中A组与B组比较的差异表达基因得到注释信息的共有190个,占A组与B组比较差异基因比为86.76%,Nr数据库注释最多,为189个;其次为Pfam数据库注释167个;KOG与COG数据库注释最少,分别为100个、66个;A组与C组比较的差异表达基因得到注释信息的共有265个,占比89.23%,其中Nr数据库注释最多为260个、其次为eggNOG与GO数据库均注释217个、Pfam数据库注释215个,而KOG和COG注释最少,分别为131个、65个;B组与C组比较的差异表达基因得到注释信息的共有59个,占比80.82%,其中Nr数据库注释最多,为55个,其次为eggNOG数据库注释46个,而KOG和COG数据注释最少,分别为27个、10个。
2.6" 差异表达基因功能富集分析
将所有草乌头叶片的差异基因同Nr数据库进行比对,显示数量分布最多的物种为变色耧斗菜(Aquilegia coerulea),有193个,占比46%;其次为唐松莲花(Thalictrum thalictroides),有99个,占比24%;其余物种占比均相对较低,博落回(Macleaya cordata),有17个,占比4%;罂粟(Papaver somniferum),有8个,占比2%;莲(Nelumbo nucifera)、葡萄(Vitis vinifera)及沉水樟(Cinnamomum micranthum f. kanehirae),分别有6,5,4个,占比均约为1%,其他共有89个,共占比21%(图4)。
为进一步了解差异基因表达基因生物学功能,本研究采用GO富集对DEGs进行功能富集分类。GO富集分析的结果有由生物过程、细胞组成和分子功能三个部分组成。对A组与B组比较进行富集分析,GO数据库注释到的DEGs有163个,其中,生物过程(biological process)中的最显著富集的是代谢过程(metabolic process)和碳水化合物代谢过程(cellular process);在分子功能(molecular function)中,富集最显著的分子功能是催化活性(catalytic activity)和结合(binding)。对A组与C组比较进行富集分析,GO数据库注释到的DEGs有217个,其中富集到biological process中的最显著的是metabolic process;在molecular function中,富集最多的是binding。在B组与C组比较的GO富集分析中,GO数据库注释到的DEGs有42个,其中富集到biological process中的最显著的是metabolic process以及cellular process;在molecular function中,富集最多的是transcription regulator activity(图5)。
将筛选出来草乌头转录组差异基因与KEGG数据库进行比对分析代谢途径,结果表明差异基因成功富集于83个代谢途径通路,主要集中在植物激素信号转导(Plant hormone signal transduction)、植物-病原体相互作用(Plant-pathogen interaction)以及MAPK信号通路-植物(MAPK signaling pathway-plant),分别为17,14,13个差异基因。说明遮荫处理可能对草乌植物的生长和应答产生显著影响,特别是对于激素信号传导、病原体相互作用和MAPK信号途径等生物学过程(表6)。
2.7" 转录因子分析
通过与植物转录因子数据库比对,仅鉴定出44个转录因子,分布于14个转录因子家族,其中随着遮荫程度的加大,NAC,GRAS,C2H2,Tify,WRKY,bZIP,C2C2-GATA,C3H转录因子仅表现为显著下调;FAR1,MYB-related仅表现为显著上调;AP2/ERF-ERF,bHLH,HB-other则均为混合调控。
2.8" 关键基因相对表达量分析
在其显著富集的通路中,依据基因的表达量和差异情况(q-valuelt;0.01,FCgt;2)随机选择差异基因,分别为光合作用相关基因(LHCB17,LHCB1,PetA,ATPβ)、植物生长发育和激素调控相关基因(NCED,GA2ox,DELLA)、能量代谢和代谢途径相关基因(AMY,PFK)以及信号转导和调控途径相关基因(PP2C),共10个基因。其相对表达量和转录组FPKM的线性回归的R2为0.856 9,证明了转录组数据的可靠性(图6)。
为验证草乌头叶片转录组测序结果的准确性,随机选择10个差异基因设计引物,其中所选差异基因中上调基因有GA2ox,LHCB1,下调基因有LHCB17,PFK,NCED,PP2C,DELLA,AMY,ATPβ和PetA。经实时荧光定量表达分析,结果表明10个差异基因表达量与转录组数据显示趋势一致(图7)。
3" 讨论
光照强度显著影响药用植物的生长和活性成分的合成,进而影响药材的品质。研究表明,不同植物对光照条件的需求各不相同,光照对药用植物的影响也因此各异。例如,延胡索[19]、金莲花[20]、丹参[21]和三七[22]等植物在不同光照条件下表现出不同的生长和活性成分积累模式。这些差异提示了光照管理在药用植物栽培中的重要性,为优化栽培条件以提高药材品质提供了依据。遮荫处理作为模拟自然生长环境中光照条件变化的手段,在研究药用植物光照适应性和次生代谢调控机制方面具有重要意义。随着高通量测序技术的快速发展,遮荫对植物发育调控的分子机制研究逐渐成为热点。本研究对遮荫处理的草乌头叶片进行了转录组测序,获得了56.78 Gb的高质量序列,所有样品的Q30值均≥94.37%,显示出良好的测序质量。拼接组装后得到59 082个Unigenes,其中27 668个Unigenes获得了功能注释,占总Unigenes的46.83%。在差异表达基因筛选中,采用Fold Change≥2、FDRlt;0.01作为筛选标准,对不同处理样本进行了比较分析。结果显示,AvsC比较组具有较多的差异基因,而BvsC比较组的差异基因数量最少,共筛选出483个差异基因。这一数量远低于草乌头种子休眠转录组的差异基因数量[1],以及其他物种在遮荫处理下的差异基因数量[23-24]。这种差异可能与不同物种、不同部位的特性以及测序数据的可靠性有关。差异表达基因共注释421个,占比87.16%。差异基因同Nr数据库进行比对,显示数量分布最多的物种为毛茛科耧斗菜属变色耧斗菜(Aquilegia coerulea),占比46%;其次为毛茛科小银莲属唐松莲花(Thalictrum thalictroides),占比24%,两者均与草乌头同属于一科,反映其物种相似性。GO富集分析的结果由生物过程、细胞组成和分子功能三个部分组成,注释到的DEGs有163个,并且由于差异基因不仅仅只与其中某一个过程相关,也可分类于其他组别,三个比较组中存在一定相似性,如:AvsB比较组与A vs C的biological process中的最显著富集的均为metabolic process。用KEGG数据库进行比对分析代谢途径表明差异基因富集于83个代谢途径通路,主要集中在植物激素信号转导、植物-病原体相互作用以及MAPK信号通路-植物,分别为17,14,13个差异基因。说明遮荫处理可能对草乌头植物的生长和应答产生显著影响,特别是对于激素信号传导、病原体相互作用和MAPK信号途径等生物学过程。对草乌头差异基因所在转录因子家族进行分析可知,其主要分布在AP2/ERF-ERF家族,表达情况分别为NAC,GRAS,C2H2,Tify,WRKY,bZIP,C2C2-GATA,C3H转录因子仅表现为显著下调;FAR1,MYB-related仅表现为显著上调;AP2/ERF-ERF,bHLH,HB-other为混合调控。发现为了解草乌头基因调控网络提供了重要线索,同时也表明不同转录因子家族在草乌头中具有多样化的调控模式。草乌头叶片在遮荫处理下表达了一系列与光信号传导密切相关的基因,如PsbA,PsaO,PetB,PetE。PetA是参与光合作用中电子传递链的一部分,其下调可能意味着光合电子传递活性的降低或调节;LHCB17是光合作用中叶绿体膜上的一个叶绿素a/b结合蛋白基因,其下调可能反映了叶绿体中光合色素复合物的变化,可能与光合作用效率或光能捕获能力的调节有关;LHCB1也是叶绿素a/b结合蛋白基因,其上调可能表示植物对于光合色素复合物的重新组装或调整,以适应遮荫条件下的光能利用;ATPβ是F-ATP酶复合物的一个亚基,其下调可能反映了ATP合成酶活性的调节或ATP能量代谢的变化。此外还发现了与激素调控、能量代谢等多个生物学过程相关的基因(如NCED,PP2C,GA2ox,AMY,PFK,DELLA等),它们的差异表达与光照条件密切相关。这些基因的变化揭示了草乌头叶片在不同光照强度下的转录组调控机制。这一发现不仅拓展了我们对草乌头光调控生物学过程的认识,也为进一步探究其适应性和生长调控提供了更深入的视角。
本研究对草乌头叶片在光照条件下的分子响应进行了深入探究,揭示了其对光照变化的适应机制。光信号传导相关基因的差异表达反映了草乌头叶片在光照调节方面的复杂性和精细调控能力。这些发现不仅丰富了对草乌头的生理生化机制的理解,还为研究其在不同环境下的生长和发育提供了科学依据。进一步研究将有助于优化草乌头的种植管理策略,提高其药用成分的产量和品质,促进其在药用和保健领域的应用。
4" 结论
通过对遮荫处理后草乌头叶片的转录组测序和分析,我们揭示了光照对草乌头栽培的重要影响。遮荫处理显著改变了多个基因的表达,特别是与光合作用、能量代谢和生长发育相关的PsaO,PetB,LHCB17,ATPβ等基因。这表明光照对草乌头的光合能力和生长发育具有重要调节作用。研究这些基因的调控机制,为草乌头的合理栽培和利用提供了科学依据,有助于提高其有效成分产量和药用品质。高通量测序和生物信息学的发展,为我们深入理解草乌头及其他药用植物的分子调控机制提供了新方法。
参考文献
[1]" 樊东昌. 草乌头种子休眠解除过程生理生化变化及转录组分析[D]. 呼和浩特:内蒙古农业大学,2022:1-5
[2]" 李谦,过立农,郑健,等. 乌头属药用植物的研究进展[J]. 药物分析杂志,2016,36(7):1129-1149
[3]" 赵翡翠,李杰. 我国乌头属药用植物中生物碱分析方法的研究进展[J]. 中药材,2010,33(8):1351-1354
[4]" 郑权发,张宇,陈芳,等. 乌头汤、四妙散对胶原诱导性关节炎DBA/1小鼠的作用研究[J]. 湖南中医药大学学报,2022,42(4):564-570
[5]" 荣宝山,黄凯丽,袁琳嫣,等. 乌头类药材化学成分和药理作用研究进展[J]. 中国药事 2021,35(8):932-947
[6]" 贾春赫,吕丽娟,李晓杰,等. 草乌幼苗形成期叶绿体发育的转录组分析[J]. 草地学报,2024,32(6):1779-1788
[7]" 郭巧生.药用植物栽培学[M]. 北京:高等教育出版社,2010:28-38
[8]" 朱萱,高康,滕文军,等. 遮荫对5种观赏草表型的影响及耐荫性评价[J]. 草地学报,2023,31(7):2096-2106
[9]" 李振陆.植物生产环境[M]. 北京:中国农业出版社,2006:131-152
[10]YANF F,HUANG S,GAO R,et al. Growth of soybean seedlings in relay strip intercropping systems in relation to light quantity and red:far-red ratio[J]. Field Crops Research,2014,155:245-253
[11]ZHEN S,IERSEL V W M. Far-red light is needed for efficient photochemistry and photosynthesis[J]. Journal of Plant Physiology,2017,209:115-122
[12]张伟,孟祥庆,苏晓荟,等. 光调控对药用植物次生代谢成分合成的影响[J]. 药学实践与服务,2024,42(2):50-59
[13]刘紫祺,王仪,王秀,等. 不同光照强度对西洋参生长、皂苷含量及基因表达的影响[J]. 中国中药杂志,2022,47(18):4877-4885
[14]WANG Y,HTWE Y M,LI J,et al. Integrative omics analysis on phytohormones involved in oil palm seed germination[J]. BMC Plant Biology,2019,19(1):363-377
[15]KHAN A. Primary preventive and permissive roles of hormones in plant systems[J]. Botanical Review,1975,41(4):391-420
[16]樊东昌,穆赢通,张晓明,等. 北乌头种子休眠解除过程中营养物质的代谢变化研究[J]. 耕作与栽培,2022,42(1):48-50,56
[17]HAN C,ZHEN S M,ZHU G G,et al. Comparative metabolome analysis of wheat embryo and endosperm reveals the dynamic changes of metabolites during seed germination[J]. Plant Physiology and Biochemistry,2017,115:320-327
[18]ZHAO M,ZHANG H X,YAN H,et al. Mobilization and role of starch,protein,and fat reserves during seed germination of six wild grassland species[J]. Frontiers in Plant Science,2018,9:234
[19]ZAFAR M S,MUHAMMAD F,JAVED I,et al. White mulberry (Morus alba):A brief phytochemical and pharmacological evaluations account[J]. International Journal of Agriculture amp; Biology,2013,15(3):1560-8530
[20]牛欢,韦坤华,徐倩,等. 不同光照度对金线莲生长、生理特性和药用成分的影响[J]. 植物资源与环境学报,2020,29(1):26-36,43
[21]冯思念,王瑞,顾雯,等. 不同强度的红蓝光质对丹参根系形态和有效成分积累的影响[J]. 中草药,2019,50(21):5313-5318
[22]陈黎明,罗美佳,夏鹏国,等. 光强对三七生长、光合特性及有效成分积累的影响[J]. 时珍国医国药,2016,27(12):3004-3006
[23]张鑫鑫. 不同光照条件下胡桃楸幼苗生长、生理及分子响应机制研究[D]. 哈尔滨:东北林业大学,2024:35-47
[24]杨永庆,马晓蕾,李玉荣,等. 不同光源条件下与花生株高变化相关的转录组分析[J]. 四川农业大学学报,2024,42(1):19-27
(责任编辑" 闵芝智)
猜你喜欢
中国中药杂志(2017年4期)2017-03-28 17:06:08
中国中药杂志(2017年2期)2017-03-25 17:23:23
中国中药杂志(2017年1期)2017-03-06 21:20:38
中国中药杂志(2016年22期)2017-02-13 17:06:29
安徽农学通报(2015年8期)2015-07-02 01:36:11
江苏农业科学(2014年1期)2014-07-18 21:05:24
