
温度和降水对宁夏六盘山自然保护区蒙古栎年内木质部生长的影响
2024-03-19 08:14:20李乾林郭明钢李佳音郭霞丽黄建国李学斌
植物研究 2024年2期
李乾林 郭明钢 李佳音 郭霞丽* 黄建国 陈 林 李学斌
(1.广西大学林学院,广西森林生态与保育重点实验室,南宁 530004; 2.广西大学林学院,亚热带农业生物资源保护与利用国家重点实验室,南宁 530004; 3.浙江大学生命科学学院,杭州 310058; 4.宁夏大学生态与环境学院,银川 750021)
根据联合国政府间气候变化专门委员会(IPCC)第六次评估报告指出,预计21 世纪末全球平均温度将上升1.5 ℃[1]。随之引发的干旱等极端事件将对树木生长造成严重干扰,进一步改变森林生态系统的结构和功能[2]。因此,揭示树木生长对气候变化的响应及适应机制,对预测未来的森林生长和制定科学管理政策具有重要意义。
木质部不仅发挥着关键的支撑作用[3],同时是重要的碳储存器官,其细胞壁大量储存着由树冠进行光合作用的产物[4]。因此,了解木质部生长动态有助于预测树木的生长模式和森林的固碳潜力。利用微树芯采样技术对木质部进行高频率的周期性采样,通过生物切片技术,可以监测形成层活动的关键物候期以及木质部生长[5]。目前,相关研究取得了一系列进展[6-7],且主要集中在温带和北寒带地区[8],在热带和亚热带地区以及干旱区研究相对较少。研究表明,形成层活动的物候以及木质部年内生长动态受到气候因子的控制[9-10]。在寒冷的湿润地区,温度主要决定了针叶树木质部生长的开始时间[11]。在干旱地区,木质部的生长则主要受水分限制[12]。相比于针叶树种,对阔叶树木质部生长动态及其与气候因子之间关系的研究较为不足。
宁夏回族自治区位于我国西北部,是保护黄河上中游及华北、西北地区的重要生态屏障。六盘山是宁夏回族自治区最重要的山脉之一,处于半湿润向半干旱过渡带上,该地区的树木生长对于气候变化尤为敏感。长久以来,中国西北部都是一个干旱少雨的地区,而自1980年以后,该地区逐渐趋于温暖湿润[13]。蒙古栎(Quercus mongolica)天然次生林是该区域主要地带性植被与顶级群落[14]。但是目前对蒙古栎木质部生长与气候因子之间关系的研究较少,这极大限制了对蒙古栎天然林木质部生长趋势的了解和固碳潜力的评估。
本研究通过在六盘山自然保护区设置监测点,使用微树芯采样技术对蒙古栎木质部进行为期2 a 的连续采样,定量(1)年内木质部动态关键物候期,包括开始、结束生长的日期和生长季长度;(2)年内木质部生长动态与气候因子(温度和降水)之间的关系。本研究结果为预测未来气候变化下蒙古栎木质部的生长提供了依据,为实现我国干旱半干旱区生态保护的可持续发展政策提供参考。
1 材料与方法
1.1 研究区概况
研究区位于宁夏六盘山自然保护区(35°15′~35°41′N,106°9′~106°30′E),海拔1 700~2 931 m。该区处于暖温带半湿润区向半干旱区过渡带上,属于大陆性季风气候[14]。根据靠近研究站点的靖远气象站1970—2010 年的记录,该地区年均温为6.0 ℃,年总降水量为632 mm[15]。蒙古栎林为主要的天然次生林,主要分布于海拔2 200~2 900 m的区域。2019—2020 年在六盘山秋千架林场设置样地,选择4 株树龄相近、健康、直立、长势一致的蒙古栎标准木进行年内木质部生长动态的监测。
1.2 气象因子
为了评估该地区2019 和2020 年的气候条件,从Copernicus气候变化服务网(C3S:https://cds.climate.copernicus.eu/)获取当地每小时气象数据,通过整理和计算得到每日最高温、最低温(℃)和总降水量(mm)。结果显示,该地区2 a 的平均温和最高温较为接近,然而2020 年最低温度比2019 年低6.68 ℃(见表1)。该地区总降水量2020 年高于2019 年,其中,2020 年集中在6—9 月的降水占年总降水量的70.74%(见图1),而2019 年同期降水量占全年总降水量的56.30%。
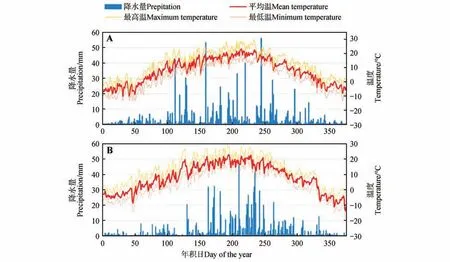
图1 研究区2019年(A)和2020年(B)每日最高温、平均温、最低温及总降水量Fig.1 Daily maximum,mean,minimum temperature and total precipitation in the study area in 2019(A) and 2020(B)

表1 研究区气候特征Table 1 Climatic characteristics of the study area
1.3 微树芯试验
为了更精确地观察到蒙古栎木质部生长的起始日期,采样频率在不同季节有所变化。在生长季前期(3—5月),每3 d使用微生长锥(Trephor,意大利)[16]在样树胸径处(1.3 m)采集1~2 个微树芯,6—11 月每7 d 取1 次样。为了避免相互影响,采集时以“Z”字形从上往下依次进行,相邻2 次采样相隔3~5 cm。所有样品立即放入装有50%乙醇溶液的离心管中固定,带回实验室后放置于冰箱中4 ℃保存[17]。
在实验室内,使用软化剂(V(甘油)∶V(无水乙醇)=1∶2)软化微树芯,软化时间为一个月,然后分别使用不同体积分数的乙醇溶液逐级脱水。之后将样品置于低、中和高浓度的柠檬烯中分别透明,再经过熔点为58~60 ℃的低、中、高浓度石蜡溶液中浸蜡后进行包埋,包埋后的石蜡块修正后用滚动切片机(Leica RM 2235,德国)制备厚度为8 µm 的木质部切片。切片用烤片机烤干后经柠檬烯和无水乙醇溶液按顺序分别脱蜡2 次。切片制备成功后,经0.06%的焦油紫染色1 min 后使用Leica 显微镜(DM500 LED,德国)观察木质部的细胞生长[17-18],并分别置于4 倍和10 倍物镜下拍照观察。利用LAS v4.12 软件测量每棵树当年生的木质部总长度,每个切片沿木质部径向生长方向随机选择3 列木质部生长长度作为重复测量求取平均值,并同时监测导管出现以及木质化的日期。日期格式采用年积日(Day of the year,DOY)来表示。
1.4 数据分析
1.4.1 木质部生长动态模拟
采用Gompertz 函数[19-20]进行拟合。Gompertz函数公式如下:
式中:y为木质部长度,A为上渐近线,β为x轴位置参数,κ为变化率参数,t为时间。
通过Gompertz 函数分析蒙古栎的相关参数:生长起始日期,生长结束日期,生长季长度,年总生长量和生长季内木质部日生长速率的拟合值。当达到年总生长量的5%和95%时,确定木质部生长开始和停止的日期[21]。运用方差分析(ANOVA)对比2年间木质部生长开始、结束和生长季长度的差异是否存在显著差异。
1.4.2 木质部生长与气候因子的关系
运用混合线性模型分析影响蒙古栎年内木质部生长的主要气候因子。以每株树采样日当天的拟合生长速率为因变量,分别以温度或者降水为固定效应因子,嵌套年份和不同树木个体为随机效应因子,探究温度、降水与木质部生长速率之间的相关性。为了更精确评估温度和降水对树木生长的影响,计算采样日前7、10、15 天内的最高、平均、最低温度和总降水量与采样日当天拟合生长速率的相关关系[22]。采用“lubridate”包分别计算采样日前7、10、15 天内的最高、平均、最低温的平均值和总降水量。
所有数据分析均使用R 统计软件[23],图表采用R统计软件中的“ggplot2”包。
2 结果与分析
2.1 木质部分化阶段的开始和结束
显微镜下分别观察第1 个导管出现以及成熟的日期(如表2 所示)。2019 年观测导管出现的日期为4 月9 日(DOY 99),成熟日期为4 月28 日(DOY 118);2020 年 导 管 出 现 日 期 为4 月6 日(DOY 97),成熟日期为4月29日(DOY 120)。
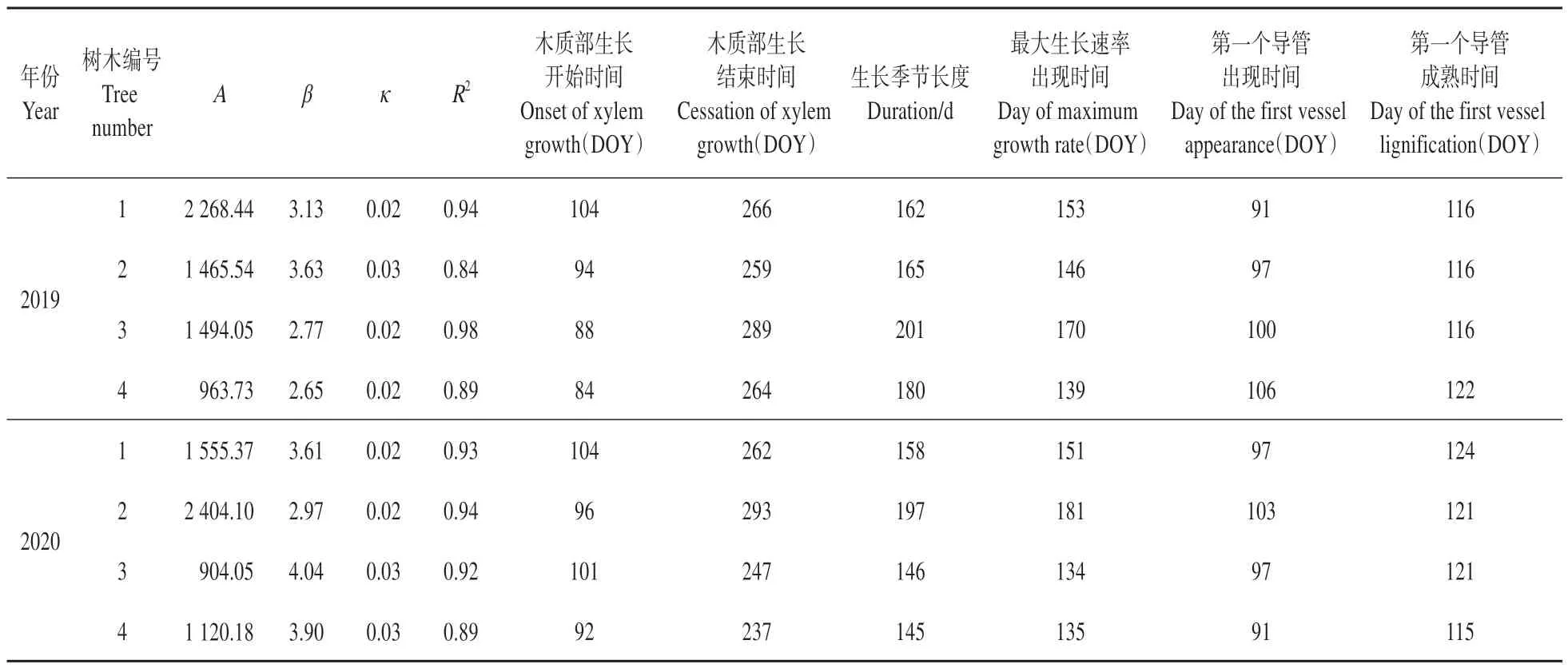
表2 2019和2020年蒙古栎年内木质部生长动态的拟合参数Table 2 Gompertz fitting parameters of intra-annual xylem growth dynamics of Q. mongolica in 2019 and 2020
使用Gompertz 函数对蒙古栎年内木质部生长动态进行拟合发现(见表2),在总体趋势上,2019年木质部开始生长的日期为4月2日(DOY 92),结束日期为9 月27 日(DOY 270);2020 年木质部开始生长日期为4月7日(DOY 98),结束生长日期为9月17日(DOY 260)。结果表明,2019年木质部生长开始日期比2020年早6 d,生长结束日期晚10 d,所以2019 年比2020 年生长季持续时间长16 d。对于树木个体而言,开始生长的时间几乎一致,但结束的日期却有较大差异,所以导致个体之间生长季长度也存在较大差异,尤其是2 号树,相比之下,在2019 年生长季最短,但在2020 年最长。最后,2 年间导管出现、成熟的日期及木质部生长开始、结束和生长季长度差异都不显著。
2.2 木质部生长动态及其发育模式
六盘山地区蒙古栎木质部的生长在2 a 中表现出相似的生长模式(见图2)。在2019 年,几乎每棵树的生长趋势都呈“S”形,生长速率曲线基本符合“倒钟”形,使用Gompertz 模型拟合解释量达84%~98%(见表3)。经过连续2 a 的监测发现木质部细胞均在4—7 月生长迅速,2019 年该阶段生长量达总生长量的76.05%,2020 同期生长量占总生长量的68.19%。而在8—10 月生长速率减小,2020 年4 棵树木质部总拟合生长量小于2019 年。2 a最大拟合生长速率分别出现在5月31日(D0Y=152)和6月9日(DOY=161),2019年平均生长速率为8.74 µm·d-1,2020年为9.26 µm·d-1。
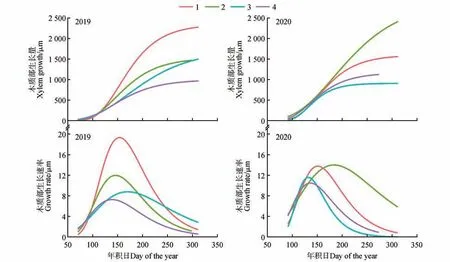
图2 2019年和2020年蒙古栎年内木质部生长动态及生长速率Fig.2 The intra-annual xylem growth dynamics and growth rate of Q. mongolica in 2019 and 2020

表3 2019与2020年蒙古栎木质部生长参数Table 3 Parameters of xylem growth of Q. mongolica.in 2019 and 2020
2.3 气象因子与木质部发育的关系
混合线性模型结果表明,温度和降水共同调节蒙古栎木质部生长速率(见表4)。前7、10、15日的最高、最低、平均温及总降水量与生长速率呈极显著相关关系(P<0.01),与最高温和平均温相比,最低温在影响木质部的生长方面发挥了更重要的作用;与采样日前7、10 日的最低温和降水量相比,前15 日的最低气温与总降水量对木质部生长的影响更显著。
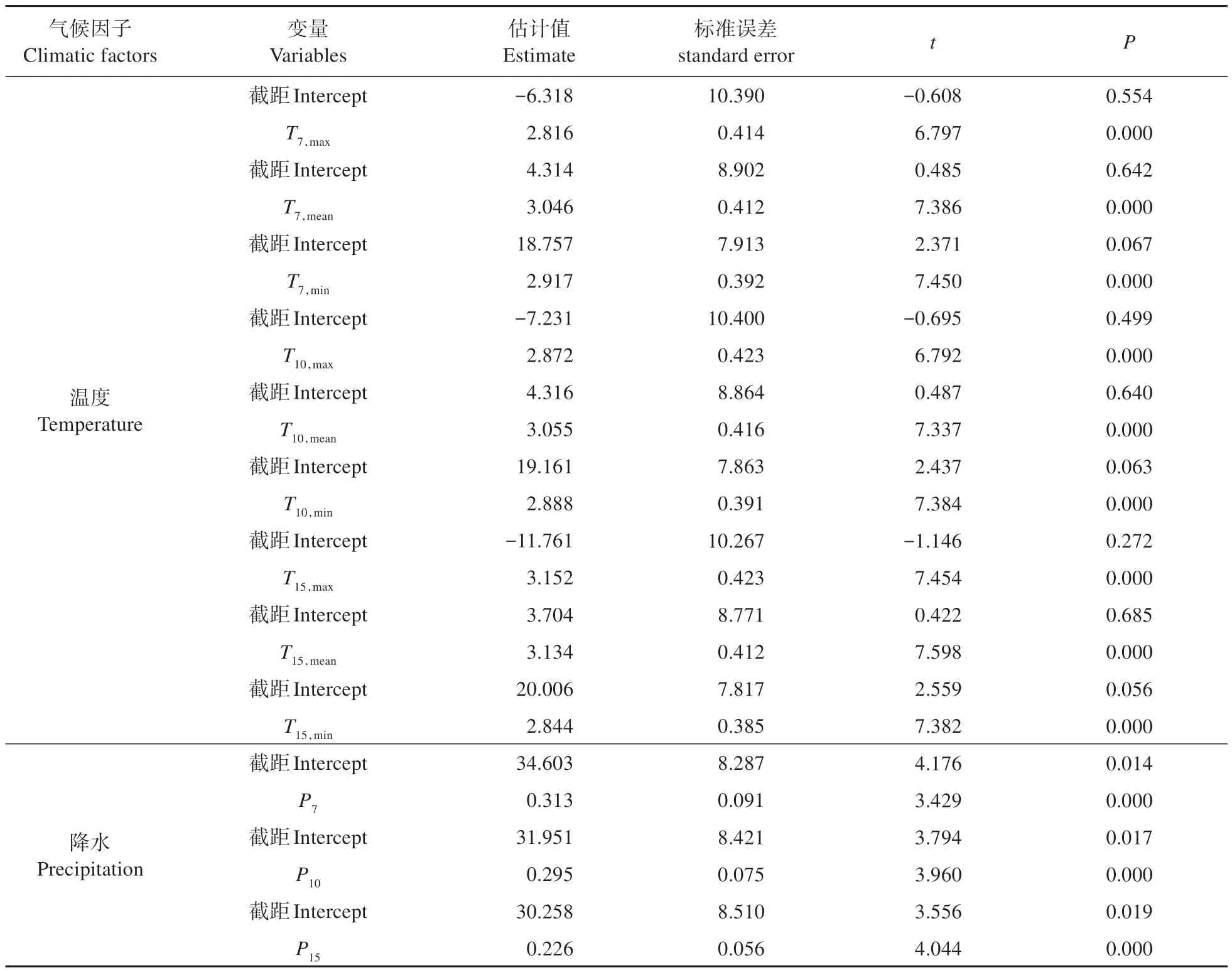
表4 气候因子对蒙古栎木质部生长速率的影响Table 4 Effects of climatic factors on xylem growth rate of Q. mongolica
3 讨论
3.1 年内木质部生长动态
通过对六盘山地区蒙古栎进行连续2 a 的监测,观察到木质部细胞生长从4 月初开始逐步加快,分别在DOY 152 和DOY 161 左右达到顶峰,之后于DOY 270 和DOY 260 停止生长。该木质部年内生长动态模式与温带和北寒带地区的研究结果[24-26]类似。对于温带和北寒带针叶树而言,温度的季节性差异性导致规律的生长模式,即木质部生长速率呈“倒钟”形,即在春季和初夏表现出快速的生长速率,最大生长速率出现在夏至日前后[27]。本研究中最大木质部生长速率的时间在5月底6 月初,比夏至日提前20 d 左右。夏至日过后,日照时间慢慢变短,蒙古栎生长速率逐步降低,确保进入休眠之前完成当年的木质部生长。此外,本研究发现2 号树在2019 年生长季最短且生长量最低,但在随后的1 年中生长季最长,生长量达到最大。黄土高原中部蒙古栎年轮宽度每年的波动差异也较大[28],这可能与气候变化和个体差异有关,需进一步研究。
3.2 温度对木质部生长的影响
本研究发现六盘山蒙古栎木质部生长速率与温度呈极显著正相关关系(P<0.01),在温带和北寒带森林研究中也观察到了类似的结果。树木在春季生长发育之前,需要经历生理休眠期和生态休眠期2个阶段[29]。当树木处于生理休眠期,无论外界环境条件如何变化,树木均不会打破休眠。而当树木进入生态休眠时,在外界环境适宜的情况下,树木就会打破休眠开始生长。具体来说,树木需要足够的冷激打破生理休眠期,也需要一定的积温结束生态休眠期[30-31],两者均与温度密切相关。研究表明,春季温度是引发北半球中高纬度地区针叶树种木材形成开始时间的主要因素[11,32]。相较于高纬度和高海拔寒冷地区,处于较温暖地区的木质部生长开始时间比较早[24],这是由于较高温度促使树木积累了足够的有效积温,从而诱发生长。另外,生长季内较高的温度可以促进光合作用,从而积累更多的碳水化合物用于树木的生长。Huang 等[33]对我国亚热带地区马尾松(Pinus massoniana)木质部的研究表明,温度升高促进形成层细胞加速分裂,进而有利于树木木质部生长。同时,对干旱和半干旱地区的阿尔泰山脉南部西伯利亚落叶松的研究中[34]也得到类似的结果。李明明等[35]在贺兰山地区通过VS 模型模拟树木木质部形成的动态,并发现温度是生长季结束时的主要限制因子。与前人的研究结果一致,处于暖温带半湿润区向半干旱区过渡带上六盘山地区蒙古栎木质部的生长也受到温度的调控。
3.3 降水对木质部生长的调控作用
本研究中,蒙古栎木质部生长与总降水量相关性极显著(P<0.01),这与阿尔泰山南部的西伯利亚落叶松木质部生长受到降水量调节的规律[34]一致。研究表明,在我国半干旱、干旱甚至半湿润地区,水分是限制生长季节内木质部生长速率的主要因素[15]。在贺兰山中部地区,6 月降水量显著影响油松(P.tabuliformis)木质部生长,各海拔高度日径向增量与降水量呈显著正相关[36]。对于生长在青藏高原地区的祁连圆柏(Juniperus przewalskii),春季降水是其木质部开始发育的主要启动因子[37]。在高山生态系统中,研究发现瑞士五针松(Pinus cembra)的日径向生长量与降水以及相对湿度呈正相关[38]。在西北半湿润气候下,土壤有效含水量同样也会影响树木木质部生长[8]。
形成层细胞分裂、细胞增大、细胞壁增厚以及细胞成熟,是树木径向生长的主要过程[39]。在干旱半干旱地区,树木的径向生长开始时间、结束时间和生长速率主要取决于水分可利用性,这和降水、土壤含水量及相对湿度密切相关[40]。例如,对我国北方干旱地区侧柏(Platycladus orientalis)的研究表明,其木质部生长受到水分可利用性的限制[41]。水分可利用性一般通过2 个途径影响木材产量,一种是通过调节光合速率和碳水化合物储存库来间接影响形成层细胞分裂和木质部发育[42]。作为植物新陈代谢活动过程中不可缺少的介质,水分通过驱动光合作用和同化物的运输,间接影响木质部以及叶片等植物器官的生长[43]。另一种方式是通过维持细胞膨压,从而促进形成层细胞的分裂活动和木质部细胞的增大过程。细胞膨压需要保持足够的水分,这可以进一步促进形成层细胞的分裂活动和细胞的扩张和生长[44-45]。研究表明,生长季早期水分亏缺可能会降低青藏高原地区的祁连圆柏木质部细胞分裂速率[26]。降水量会直接影响土壤含水率和相对湿度,也在一定程度上影响水分可利用率。因此,降水会直接或者间接影响细胞的生长,从而影响蒙古栎木质部的生长量。
4 结论
本研究通过对宁夏六盘山地区天然次生林蒙古栎连续2 a 的年内木质部生长动态监测,发现蒙古栎木质部生长开始于4 月初,结束于9 月中旬,生长季长度为162~177 d。与以往的研究结果不同,本地区蒙古栎木质部的生长受温度和降水的共同调控。随着气温的持续上升和降水模式的改变,蒙古栎木质部的生长可能会受到显著影响。然而温度和降雨之间的交互作用对树木生长的影响,需要进一步研究,从而精准预测树木对全球变化因子的响应。
猜你喜欢
绿色科技(2021年21期)2021-11-26 09:12:06
内蒙古林业调查设计(2021年3期)2021-07-06 14:12:00
植物研究(2021年2期)2021-02-26 08:40:10
阅读与作文(小学高年级版)(2019年8期)2019-10-16 04:46:50
西域历史语言研究集刊(2018年0期)2018-11-09 01:01:40
现代园艺(2018年1期)2018-03-15 07:56:13
草原歌声(2017年1期)2017-04-23 05:08:53
天津农业科学(2015年11期)2015-12-03 00:03:49
江苏农业科学(2015年4期)2015-06-15 21:11:35
交通建设与管理(2015年15期)2015-03-20 15:19:28
