
混交对杉木和山杜英混交林生长、结构和生产力的影响
2024-03-19 08:14周浩然孙洪刚韩媛媛李文华
植物研究 2024年2期
周浩然 孙洪刚 张 鹏 韩媛媛 李文华,3
(1.东北林业大学林学院,森林生态系统可持续经营教育部重点实验室,哈尔滨 150040; 2.中国林业科学研究院亚热带林业研究所,杭州 311400; 3.北华大学林学院,吉林 132013)
森林的成分和结构多样化不仅能提升森林生态系统稳定性,同时可持续提供木材和不同类型的生态系统产品[1]。近年来,人工混交林的研究表明:混交林因其成分和结构多样化程度远高于人工纯林,有利于人工林的高产和稳产[2],所以混交林培育为实现木材可持续生产提供了一种可能。较纯林而言,混交林的生态过程相对复杂,目前仍不确定混交林产量是否一定高于纯林。混交林内如果出现种间促进或竞争减缓等正向混交效应,将利于降低树种间的生境资源竞争强度,增加资源利用效率,从而提高混交林生产力[3]。例如,在欧洲山毛榉(Fagus sylvatica)-花旗松(Pseudotsuga menziesii)混交林中,欧洲山毛榉通过根系二次生长增强水分获取能力,降低与花旗松间的水分竞争强度,并可在长期干旱时将土壤深层水分提升至土壤表层,缓解花旗松根系层的土壤水分亏缺,混交林整体生产力远高于欧洲山毛榉和花旗松各自纯林的生产力[4]。相反,具有相似生物学特性或生境需求趋近的树种间混交,可能会加剧混交林内树种间的生境资源竞争强度,进而导致混交林生产力下降。例如,因对光照需求较为接近,导致杉木(Cunninghamia lanceolata)-檫木(Sassafras tsumu)混交林的蓄积量远低于杉木纯林[5]。此外,树种间相互作用会随生境条件变化而发生改变,导致树种混交对林分生产力的影响出现空间动态变化[6]。在巨桉(Eucalyptus grandis)-马占相思(Acacia mangium)混交林中,二者间的养分互补性会随土壤总氮浓度增加而降低,导致混交林增产程度随土壤氮素含量增加逐渐降低,甚至增产变成减产[7]。而且,树种的生物学特性和培育措施对混交林分产量具有耦合效应[8]。例如,在亮果桉(Eucalyptus nitens)+银荆树(Acacia dealbata)的幼龄混交林中,银荆树的固氮作用会促进亮果桉生长,使混交林蓄积表现增产,但混交林冠郁闭程度随林分生长发育逐渐增加后,种间的光竞争强度会超过种间促进作用(固氮作用),导致混交林蓄积低于亮果桉纯林[9]。可见,混交林能否增产,主要受混交树种的内在生物学特性、外在生境资源条件和人工培育措施等因素的综合影响。
杉木纯林广泛分布于我国南方18 个省(自治区),因其生长快、产量高的特性,在缓解木材供给压力、增加林业生产经营收益等方面发挥了重要作用[10]。但长期多代连栽导致杉木人工纯林地力衰退,林分生产力逐代下降[11]。以往造林试验发现,在幼龄杉木-山杜英(Elaeocarpus sylvestris)混交林中,由于山杜英不仅满足杉木幼苗侧方庇荫需求,还因其凋落物量大且易于分解而培肥土壤,导致混交林内杉木生长量远高于杉木纯林的生长量[12]。但对于完整培育周期内,杉木-山杜英混交林内种间关系对生长、林分结构和生产力的影响及其机制还鲜见报道,这限制了杉木-山杜英混交模式在生产中的推广利用。本研究以20年生杉木-山杜英混交林及纯林为研究对象,从内在树种生物学特性和外界生境资源角度,探究杉木-山杜英混交林在生长、林分结构和生产力形成等方面的表现,从而丰富树种混交理论,并指导杉阔混交林的生产实践。
1 研究区概况
试验样地位于江西省景德镇市枫树山林场(29°32′~29°33′ N,117°36′~117°36′ E)。该区域属中亚热带季风气候,年日照时间1 800~2 000 h,年降水量1 650~1 850 mm,年平均相对湿度79%,年均温17.8 ℃。试验样地海拔150 m,地貌类型为低山丘陵。土壤类型是板岩和花岗岩风化形成的红壤,土层深度可达60 cm。天然植被类型以常绿阔叶林为主,物种种类主要有杉木、山杜英、枫香(Liquidambar formosana)、拟赤杨(Alniphyllum fortunei)、青冈(Cyclobalanopsis chungii)、茅栗(Castanea seguinii)、白栎(Quercus fabri)、乌饭树(Vaccinium bracteatum)、杜鹃(Rhododendron simsii)、胡枝子(Lespedeza bicolor)、芭茅(Miscanthus floridulus)、芒箕(Gleichenia linearis)等。
2 研究方法
2.1 样地设置和调查
2002 年冬季,在坡度20°~25°、坡长230 m 左右的杉木皆伐迹地采用1年生容器苗造林,营造杉木纯林(Pure C)、山杜英纯林(Pure E)以及杉木-山杜英混交林(Mix C-mix E)3种林分,造林株行距均为2 m×2 m,混交林混交比例为1∶1,混交方式为行状混交。造林后的3 年内,每年进行2 次幼林抚育,主要伐除对造林幼树有竞争的杂灌和高大草本。2021年12月,分别在3种林分的上坡、中坡和下坡位设置样地,共计27块样地,每块样地大小为600 m2(30 m×20 m),对上述27 块样地进行每木调查,测量并记录树种种类、胸径、树高、枝下高和冠幅等指标。在晴朗无云时,用照度计(DLY 1802,中国)在每块混交林样地内测定林下光照强度。
2.2 样品采集和处理
2022 年2~3 月,依据生长调查数据,分别在杉木和山杜英纯林样地内各选取3株标准木,每个混交林样地内选取杉木和山杜英各3株标准木,在标准木胸径处分别沿东西和南北方向,用生长锥各取木芯1根,保证钻头穿过髓心且尽量完整保留树皮,将木芯装入PVC 管并标记,共计获取36 根木芯。同时,将标准木树根部分全部挖出,为减少粗根年轮变异现象,选取2 个最粗的地下粗根,沿地面水平方向,在距离树干基部30 cm处截取每一选中粗根2 cm 厚的根盘[13],分别标记后装入带标签的密封袋,共获取72 个根盘。将木芯和根盘带回实验室,常温下干燥、打磨后,用年轮分析系统Win-DENDRO(STD4800,加拿大)测量年轮宽度[14]。
在混交林样地内按梅花点法选择5个采样点,去除地表凋落物后,用土钻(内径5 cm)分别钻取0~5 cm 的土壤样品,分别装入带标签的密封袋,用便携式土壤水分/温度/电导率测定仪(TDR 150,中国)同步测量各样点土壤体积含水率和温度。将样品装入干冰密封箱保存带回实验室,挑出树根和杂质后,土壤样品过筛风干,并测定土壤酸碱度、土壤有机碳、土壤水解性氮和土壤有效磷含量[15-16]。
2.3 林分蓄积量和地位指数计算
采用二元材积经验式测算杉木单株材积[17],采用平均实验形数法测算山杜英单株材积[18],林分蓄积量是样地内所有活立木材积的加和。计算公式如下:
式中:VC为杉木单株材积;D为胸径;H为树高;VE为山杜英单株材积;g1.3为胸高断面积;f为山杜英平均形数是0.40。
由于杉木-山杜英混交林中,杉木处于优势地位,为评价混交林分立地质量,在9 块杉木纯林样地内选取5 株优势木,计算其树高平均值,并依据《全国杉木地位指数表》进行地位指数(SI)划分[19]。
2.4 混交效应评估
混交效应采用混交林相对生产力和混交林绝对生产力进行评估[20]。相对生产力计算公式如下:
式中:PR(C,E)为混交林的相对生产力;PC,E为混交林生产力的观测值;PC,E为混交林生产力的预期值,根据两树种纯林生产力加权平均所得(PC,E=PC×mC+PE×mE),PC为纯林中杉木生产力,mC为杉木的混交比例,PE为纯林中山杜英生产力,mE为山杜英的混交比例;PR[C,(E)]为混交林中的杉木相对生产力;PC,(E)为混交林中的杉木生产力;PR[(C),E]为混交林中的山杜英相对生产力;PC,(E)为混交林中山杜英生产力。若相对生产力>1,为混交林或混交林中某一树种产量增加;若相对生产力<1,为混交林或混交林中某一树种产量减少;若相对生产力=1,为混交林或混交林中某一树种产量不变[20]。当相对生产力与1 不同时,表示存在混交效应,采用单样本t检验,确定相对生产力与1 是否存在显著差异(P<0.05)[21]。
绝对生产力计算公式如下:
式中:PA(C,E)为混交林的绝对生产力、PA[C,(E)]为混交林中的杉木绝对生产力、PA[(C),E]为混交林中的山杜英绝对生产力。
2.5 林分直径分布分析
对样地内所有活立木,以2 cm 径阶距划分径阶,并统计各径阶株数。用2 参数Weibull 模型拟合杉木-山杜英混交林、杉木和山杜英纯林的直径分布情况,并计算偏度(SK)和峰度(K)来描述混交林及其纯林的直径分布特征[18],计算公式如下:
式中:n为林分内活立木株数;xi为林分活立木直径;-x为林分算术平均直径;σ为标准差。
2.6 异速生长分析
利用高径比、冠长率和冠径比分析林分地上生长分配特征。其中,高径比是树高与胸径的比值,冠长率是冠长与树高的比值,冠径比是冠幅与胸径的比值。
使用胸径和粗根直径指标,描述林分地上和地下生长分配特征,并用幂指数方程的对数形式进行拟合:
式中:y为粗根直径;β为常数项;x为胸径;α为异速生长指数;所有数据经对数转换为线性关系:
2.7 混交林混交效应与立地条件相关分析
为分析混交效应与立地条件之间的因果关系,本研究以相对生产力作为因变量,以杉木-山杜英混交林、混交林中杉木和混交林中山杜英的地位指数(SI)作为自变量进行线性回归,用拟合优度检验回归模型精度。
2.8 数据处理与分析
利用单因素方差分析(One-way ANOVA)和多重比较(Duncan 检验),比较不同林型和不同树种的生长差异及生长分配差异和不同坡位混交林的生境因子差异(P<0.05)。用单样本t检验分析树种混交对林分产量和结构的影响。用标准化主轴法完成异速生长分析[22]。排序轴长度为0.1 而采用冗余分析解析混交林生长状况与环境因子的耦合关系,用前向选择的逐步回归选取显著影响混交林生长状况的环境因子,并采用蒙特卡洛检验对环境因子进行排序。
单因素方差分析、单样本t检验以及林分直径分布拟合均在IBM SPSS Statistics 19 软件中完成,标准化主轴分析在R 4.2.0 软件中完成(R 包:smatr),冗余分析在Canoco 5 软件中完成。用Microsoft Excel 2019和Canoco 5作图。
3 结果与分析
3.1 杉木-山杜英混交林及其纯林的生长分析
杉木和山杜英的生长对混交的响应不同。除冠长外,混交林中杉木的胸径、树高、枝下高和冠幅均大于杉木纯林,尤其是胸径比纯林高出1.84 cm(P<0.05)。而混交林中山杜英的胸径、树高、枝下高、冠长和冠幅均小于山杜英纯林,其中胸径和树高分别比山杜英纯林低2.63 cm、1.81 m(P<0.05)(见表1)。

表1 杉木-山杜英混交林及其纯林生长情况Table 1 Growth characteristics of C. lanceolata and E. sylvestris in the pure and mixed stands
比较混交林内杉木和山杜英的生长指标发现,杉木的胸径、树高、枝下高和冠高均显著大于山杜英(见表1),其中,杉木的胸径和树高分别比山杜英大5.84 cm、3.62 m(P<0.05)。但山杜英冠幅比杉木大0.74 m,且差异显著(P<0.05)。
杉木-山杜英混交林蓄积观测值(165.64 m3·hm-2)比混交林蓄积预期值(158.94 m3·hm-2)高6.7 m3·hm-2,混交林蓄积观测值高于山杜英纯林蓄积量(137.88 m3·hm-2),但低于杉木纯林蓄积量(179.99 m3·hm-2),且杉木-山杜英混交林蓄积相对生产力为1.05,无显著差异(P>0.05)(见表2,图1),这表明杉木-山杜英混交林蓄积量只是相对增产,无显著增产表现。混交林中杉木的单株材积相对生产力和蓄积相对生产力分别是1.13 和1.15,均显著增产(P<0.05)。而混交林中山杜英的单株材积相对生产力和蓄积相对生产力分别是0.91 和0.87,没有增产(P<0.05)。由此表明,主要是混交林中杉木蓄积量显著高于纯林,导致混交林增产。此外,在混交林内,杉木的单株材积和蓄积量分别比山杜英高0.051 m3和48.80 m3·hm-2(见图1)。
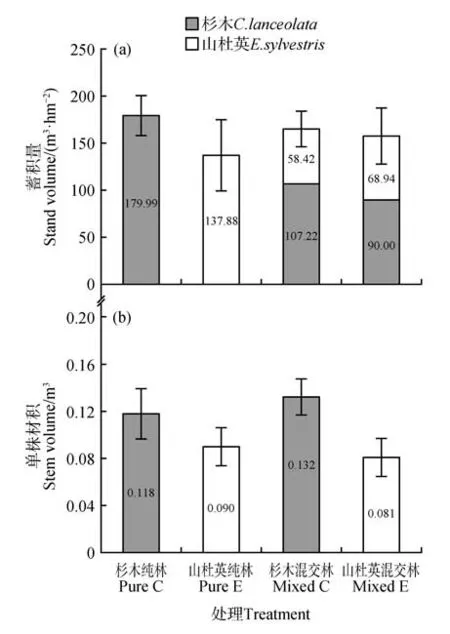
图1 杉木-山杜英混交林及其纯林蓄积量和单株材积Fig.1 The standing volume and individual volume of C.lanceolata and E. sylvestris in the pure and mixed stands

表2 杉木-山杜英混交林单株材积和蓄积的混交效应Table 2 Mixed effect of stem volume and stand volume in mixed stands of C. lanceolata and E. sylvestris
3.2 杉木-山杜英混交林结构特征
混交林中两树种结构均与纯林存在明显差异(见图2)。无论混交林还是纯林,杉木的直径分布均呈单峰分布。与杉木纯林相比较,混交林中杉木直径分布曲线的峰值呈现向高阶移动趋势,曲线峰值比纯林高0.013。相对于山杜英纯林,混交林中山杜英直径分布曲线的峰值呈现向低阶移动趋势,且混交林曲线峰值比纯林低0.012。
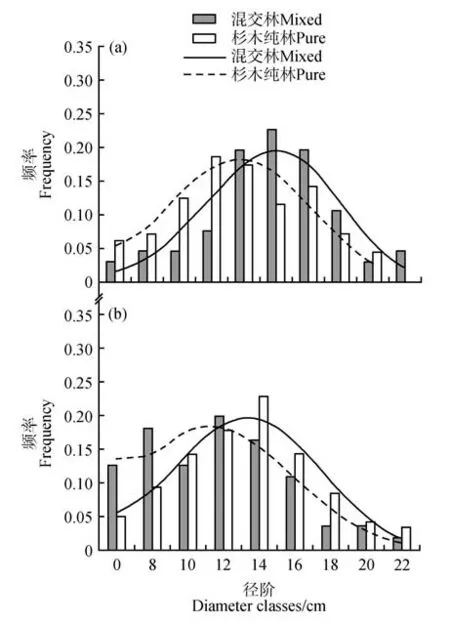
图2 杉木(a)、山杜英(b)在不同林型的直径径阶分布差异Fig.2 The difference distribution of C. lanceolata(a) and E. sylvestris(b) diameter class under different stand type
采用偏度和峰度指标分析林分的直径分布特征(见表3)。杉木在混交林和纯林中的直径分布偏度分别为-0.58 和0.08,峰度分别为0.13 和-0.87,其中峰度的差异达到显著水平(P<0.05)。可见,混交林直径分布曲线左偏,而纯林直径呈正态分布,且混交林中杉木的直径分化程度显著小于纯林。与山杜英纯林相比,混交林中山杜英直径分布的偏度和峰度均存在显著差异(P<0.05)。山杜英在纯林中直径分布偏度和峰度分别为-2.64 和5.29,在混交林中直径分布偏度和峰度分别为0.54 和-0.28,这表明混交林直径分布曲线右偏,纯林直径分布曲线左偏,混交林中山杜英的直径分化程度显著大于纯林。

表3 杉木-山杜英混交林及其纯林胸径径阶分布特征值Table 3 Characters distribution of C. lanceolata and E. sylvestris diameter class in the pure and mixed stands
3.3 杉木-山杜英混交林异速生长分析
采用高径比、冠长率和冠径比描述林分的地上生长分配特征(见表4)。山杜英纯林的高径比、冠长率和冠径比均低于其混交林,分别低0.11、0.06 和0.04,其中高径比差异达到显著水平(P<0.05)。混交林中杉木的高径比和冠长率分别比杉木纯林低0.08 和0.03,其中高径比差异显著(P<0.05),冠径比虽然比纯林高0.01,但差异并不显著。比较混交林中杉木与山杜英生长发现,山杜英的高径比、冠长率和冠径比均大于杉木,且差异皆达到显著水平(P<0.05)。

表4 杉木-山杜英混交林及其纯林的形态特性Table 4 Morphological traits of C. lanceolata and E. sylvestris in the pure and mixed stand
杉木-山杜英混交会改变两树种的地上(树干)和地下(粗根)生长分配特征(见表5)。纯林中杉木的粗根直径-胸径的主轴回归(SMA)斜率为1.166,显著高于混交林中杉木的0.770(P<0.05)。纯林中山杜英的粗根直径-胸径的SMA斜率为1.305,显著低于混交林中山杜英的1.689(P<0.05)。就两树种而言,纯林中山杜英的粗根直径-胸径的SMA 斜率显著高于纯林中杉木(P<0.05),这表明杉木和山杜英混交进一步加剧林木异速生长分配的种间差异(见表5)。
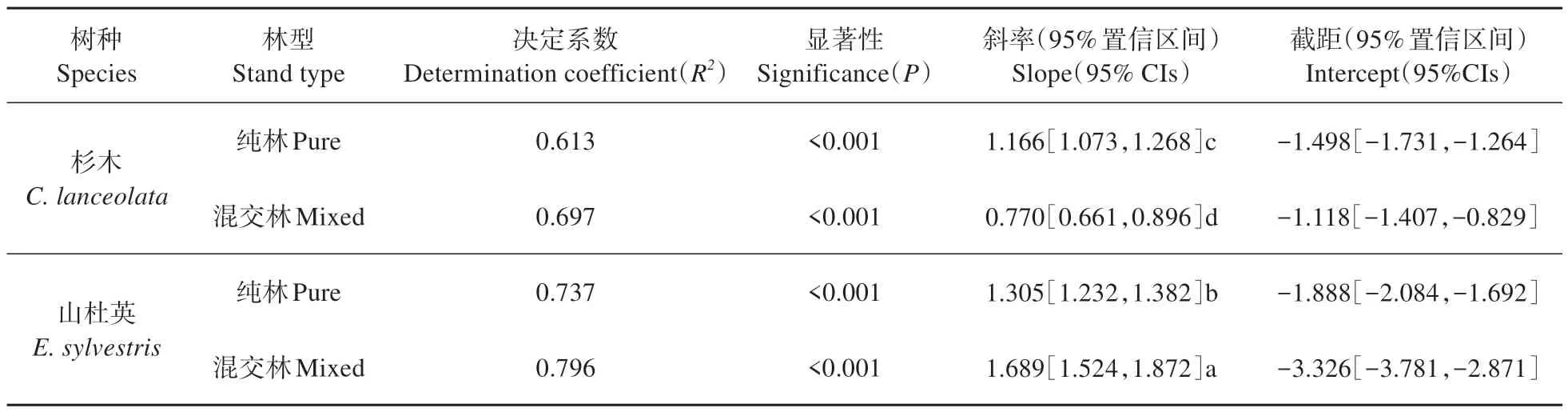
表5 杉木-山杜英混交林及相应纯林林木的粗根直径-胸径标准化主轴回归参数Table 5 Standardized major axis(SMA) regression parameters of root-stem of trees of C. lanceolata and E. sylvestris in the pure and mixed stands
坡位对杉木和山杜英粗根直径-胸径异速生长关系具有显著影响(见表6)。无论林型,杉木和山杜英的粗根直径-胸径的SMA 斜率在不同坡位间均存在显著差异(P<0.05),且表现出斜率随坡位由上到下而减少的趋势。混交林中杉木的SMA斜率在各坡位为0.275~0.795,其中上坡位(0.795)和中坡位(0.617)都显著高于下坡位(0.275),上坡位的SMA 斜率比下坡位高0.520。纯林中杉木的SMA 斜率在各坡位为0.465~1.453,其中上坡位(1.453)、中坡位(0.911)以及下坡位(0.465)由上到下依次显著降低,上坡位的SMA 斜率比下坡位高0.988。混交林中山杜英的SMA 斜率在各坡位为1.205~1.855,其各坡位的斜率差异趋势与混交林中杉木一致,但上坡位和下坡位的SMA 斜率差值(0.650)比混交林中杉木高。纯林中山杜英的SMA 斜率在各坡位为0.987~1.445,其各坡位的斜率差异趋势与混交林中两树种一致,而上坡位到下坡位的SMA 斜率差值(0.458)比混交林中两树种都低(见表6)。

表6 不同坡位的杉木-山杜英混交林及相应纯林林木的粗根直径-胸径标准化主轴回归参数Table 6 Standardized major axis(SMA) regression parameters of root-stem of trees of C. lanceolata and E. sylvestris in the pure and mixed stands in different slope position
3.4 杉木-山杜英混交林生产力与立地条件的关系
采用相对生产力和绝对生产力评估杉木-山杜英混交林分蓄积的混交效应(见图3~4)。以混交林预期蓄积量为参照,有6块混交林的相对生产力>1,3 块混交林的相对生产力<1,混交林的绝对生产力为-19.31~28.56 m3·hm-2。这表明绝大多数杉木-山杜英混交林表现增产。与相应纯林蓄积量相比较,所有混交林中杉木的相对生产力>1,其绝对生产力为8.02~56.86 m3·hm-2,杉木表现混交受益。除2 块混交林中山杜英的相对生产力>1外,其余混交林中山杜英的相对生产力<1,其绝对生产力为-54.39~-7.26 m3·hm-2。由此表明,绝大多数混交林中山杜英表现生长受限,仅在上坡位的混交林和混交林中存在正效应。
立地条件降低会增加杉木-山杜英的混交效应(见图5)。以20 年生杉木纯林的优势木平均高指示混交林立地条件,混交林在上坡位的地位指数为12,在中坡位的地位指数为14,在下坡位的地位指数为16。此外,在地位指数与相对生产力的相关关系中,除混交林中杉木的相对生产力与地位指数无显著关系外,混交林和混交林中山杜英均与地位指数呈显著负相关(P<0.05)。由此表明,主要因地位指数对混交效应的负效应,导致上坡位混交林和混交林中存在正效应。
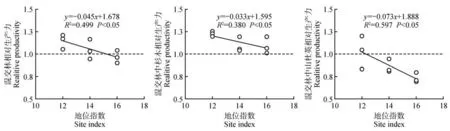
图5 混交林相对生产力与地位指数的关系Fig.5 Relationship between relative productivity and site index in C. lanceolata and E. sylvestris mixed stands
坡位显著影响土壤理化因子,对光照强度却无显著影响(见表7)。土壤含水率、土壤有机质、土壤水解性氮和土壤有效磷的测定值均随坡位由上到下增加,其中土壤有机质、土壤水解性氮和土壤有效磷的测定值,在上、中、下3 个坡位间均存在显著差异(P<0.05),而土壤含水率表现为下坡位显著大于中坡位和上坡位,上坡位和中坡位间差异不显著。此外,土壤酸碱度和土壤温度的测定值随坡位由上到下减少,其中土壤酸碱度在上、中、下3 个坡位间均存在显著差异,而土壤温度仅表现为上坡位和下坡位间存在显著差异。
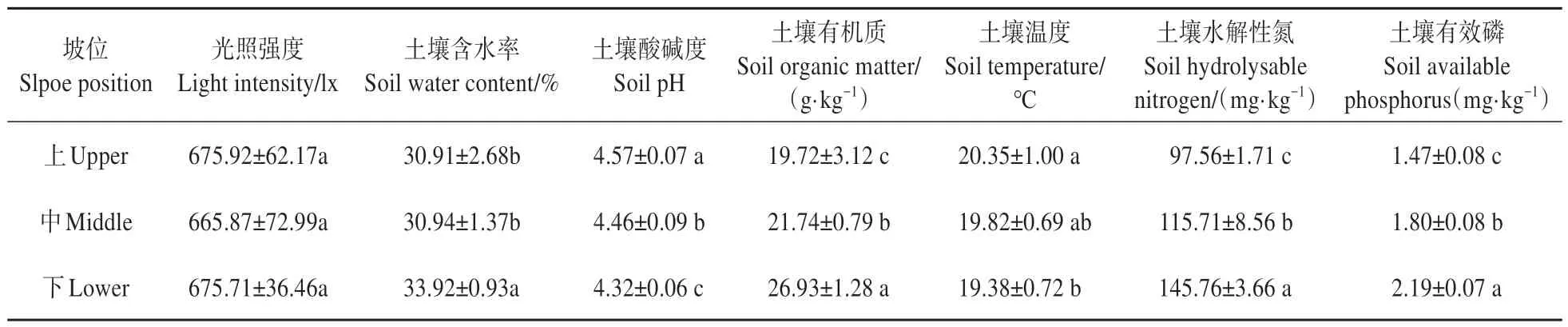
表7 不同坡位杉木-山杜英混交林的生境因子Table 7 Habitat factors of C. lanceolata and E. sylvestris mixed stands under different slope positions
为定量分析杉木-山杜英混交效应的直接影响因子,以混交林生长状况作为响应变量,光照强度和土壤理化因子作为环境解释变量进行冗余分析。前向选择逐步回归结果表明,土壤有效磷、光照强度和土壤酸碱度3 个环境因子与混交林生长状况呈显著相关关系(P<0.05)(见表8)。

表8 杉木-山杜英混交林生长状况与环境因子的前向选择结果Table 8 Results of forward selection on growth characters and environmental factors of C. lanceolata and E. sylvestris mixed stands
根据前向选择结果进行冗余分析,结果表明第一轴和第二轴分别解释混交林生长状况变异程度的46.31%和12.50%,即前两轴累积解释混交林生长状况的58.81%(见图6)。其中,土壤有效磷与相对生产力呈负相关关系,与混交林的其余生长状况指标呈正相关关系。而光照强度与冠长率和高径比呈负相关关系,与相对生产力、蓄积量、冠径比和胸径呈正相关关系,但与树高的相关性不明显。土壤酸碱度仅与相对生产力呈正相关关系,与混交林其他生长状况指标呈负相关关系。此外,通过对环境因子进行蒙特卡洛检验排序,发现对混交林生长状况影响最大的因子是土壤有效磷,解释量达到45.8%,是第二个关键环境因子光照强度的3.9倍,最后是土壤酸碱度(3.9%)。
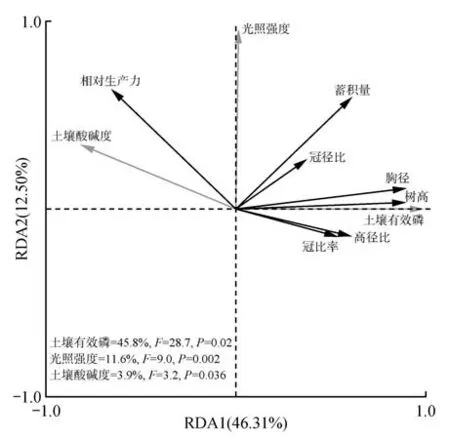
图6 杉木-山杜英混交林生长状况与环境因子的冗余分析Fig.6 Redundancy analysis on growth characters and environmental factors in C. lanceolata and E.sylvestris mixed stands
4 讨论
4.1 混交对林分生长的影响
生物学特性的种间差异可改变生境资源可利用性,进而影响林分生长[23]。有研究表明,杉木成熟林树高可达15~20 m,大于山杜英成熟林树高(9~12 m)[24]。这与本研究杉木纯林树高显著大于山杜英纯林的研究结果相似,即高生长存在种间差异,并且发现混交进一步加剧了高生长的种间差异,即混交促进杉木高生长,但显著降低山杜英高生长。混交加剧种间生长差异可能是耐荫性和高生长的生物学特性差异导致林冠垂直分层所致[4,25]。喜光杉木树冠位居较耐荫山杜英之上,相对于纯林中杉木,混交林中杉木可截获更多有效光并减缓光的个体间竞争而促进杉木生长,且混交成熟林中可能发生非对称竞争[26],大树获取大部分可利用资源,导致混交林中山杜英生长低于纯林。此外,不同林型下直径分布和高径比的种内差异,也解释了本研究中混交加剧种间生长差异的结果。已有研究表明,随林分密度降低杉木直径分布向高阶移动[27]。这与本研究中相对于纯林,混交杉木直径分布峰值向高阶移动的结果相似,表明混交林中杉木竞争减缓。其次,本研究发现混交林中山杜英高径比显著大于纯林。林木地上竞争激烈时,为获取有效光,林木会调整自身资源分配,以径向生长为代价促进高生长,导致高径比增加[28],这意味混交加剧了山杜英个体竞争状况。然而,以往研究发现,与杉木混交可减缓山杜英地上竞争而促进其生长[29]。这可能与混交效应存在时间动态变化有关[6],以往研究调查的幼龄混交林虽无明显林冠分层,但树冠构型的差异可能使山杜英占据更多生长空间而促进其生长,但随林分生长发育,林中非对称竞争程度可能逐渐增加,会抵消甚至超过对山杜英的互补作用,导致混交减缓山杜英生长。Harper[30]将混交林显著增产定义为混交林生产力超过最高产纯林。本研究在林分小尺度上发现杉木-山杜英混交林整体略微增产,并无显著增产。混交林产量较纯林无显著差异,并不代表混交树种的产量较该树种纯林无显著增产,可能因混交促进一树种生长但抑制另一树种生长,相互影响导致混交林产量无显著增产[31]。本研究中,对于纯林产量,混交林中杉木产量显著提升,而混交林中山杜英产量显著降低,即杉木-山杜英混交林增产主要因杉木增产导致。这表明杉木与山杜英混交改善了杉木的生境资源状况。
4.2 混交对林木异速生长的影响
异速生长关系反映林木不同器官间资源分配格局,为适应树种混交对生境条件的改变,林木会逐渐调整自身资源分配导致形态可塑性变化,进而影响林分结构和林分生长[32]。已有研究表明,树种混交因提升林木资源可利用性而增加林木拥挤程度[33]。这与本研究中混交林株数密度大于纯林的研究结果类似,且本研究发现混交仅增加杉木树干资源分配。这表明杉木-山杜英混交林分拥挤度增加,可能是树种混交增加杉木资源可利用性进而改变其自身资源分配造成的。然而,通常密度增加会导致植株根冠比增加,即密度加剧生长空间限制使植株以减少地上资源分配为代价,倾向把资源分配于根系[34]。这与本研究中混交山杜英根系部分资源分配更多的研究结果相似,表明混交加剧了山杜英生长空间限制。本研究中,纯林的粗根直径-胸径异速生长关系表明,当林木胸径一定时,山杜英根系资源投入显著大于杉木,这可能与两树种根型不同有关[24,35]。杉木属浅根系树种[35],而山杜英主根或斜生根较长,具有将细根位移至深层土壤的潜力[24]。已有研究表明,根系成层性会减缓种间地下竞争且增加地下资源可利用性[36]。这表示除了林冠成层性,杉木-山杜英混交林的根系成层性也可增加杉木生境资源可利用性。但林木根系倾向生长于养分富集的土壤表层[37],根系成层性更可能发生在养分贫瘠的林地。
植株资源分配格局反映植株适应生境的生长策略。最优分配理论认为植株优先向获取生长限制因子的器官分配资源[38]。本研究发现无论林型和树种,随坡位由上到下,林木个体倾向树干分配更多资源,以此保证地上部分生长和光合作用进行,即两树种采取最优分配策略适应生境变化。这与Nikolova 等[13]和Thurm 等[39]的研究结论相似,适宜生境中的林木生长侧重于树干生长,贫瘠林地中的林木生长侧重于根系生长。这表明适宜生境下杉木-山杜英混交林木的地上生长空间限制程度较大。
4.3 立地对混交效应的影响
混交效应取决于不同类型种间相互作用(如林冠成层性和根系成层性)对林分生产的综合影响,若既定生境资源可利用性在空间梯度上存在由高到低的变化,当树种间相互作用可增加既定生境资源时,混交效应会出现从负到正的空间动态变化[6]。胁迫梯度假说认为,恶劣生境条件下种间促进作用显著发生,而种间竞争作用不显著[40]。本研究发现随地位指数减少,混交林增产程度增加,这与Pretzsch 等[41]的研究结果类似,在一定程度上支持胁迫梯度的假说。地位指数反映生境因子对林分生长的综合影响[18]。本研究中,各坡位林地的土壤有效磷含量属低水平范围[42],且土壤有效磷含量随坡位由上到下呈递增趋势。进一步分析发现,土壤有效磷是解释混交林木生长的主要环境因子。这可能是因为亚热带酸性红壤磷素的固定吸附作用强,林木主要通过根系接触土壤面积获取土壤磷素,常使土壤磷素成为林木主要生长限制因子[43]。已有研究表明,随磷胁迫程度增加会影响植物根系资源分配进而使根系产生形态可塑性变化,以此保障植物磷素吸收[44]。这与本研究中两树种随坡位由下到上逐渐增加根系资源分配的研究结果相似。这表明杉木-山杜英混交效应出现空间动态变化可能与根系成层性有关,根系成层性提升土壤磷素可利用性会显著促进上坡位的杉木-山杜英混交林分生产,但下坡位林地表层土壤养分充足,杉木-山杜英混交林根系成层性不易发生,且本研究发现两树种随坡位由上到下更倾向于树干分配资源,也表明下坡位根系成层性不易发生。尽管下坡位的混交林冠成层性可增加光的可利用性,但林木的主要生长限制因子是土壤磷素,林冠成层性不能显著影响混交效应,且下坡位的混交林木的地上生长空间限制程度较大,所以下坡位的杉木-山杜英混交林表现减产。此外,本研究发现地位指数对混交林中杉木的混交效应无显著影响,这可能与杉木根系采取主动觅养策略获取土壤磷素有关[45],具体影响原因还需进一步深入研究。
5 结论
杉木与山杜英混交可改变生境资源可利用性进而影响林木资源分配、林分直径分布和林分生长。由于两树种存在耐荫性和根型等方面的差异,混交改变地上和地下的生境资源可利用性,相对于纯林,使混交林中杉木倾向地上部分分配资源,使山杜英倾向地下部分分配资源,导致混交林中杉木直径分布向高阶移动,混交林中山杜英高径比显著增加,即混交减缓杉木地上竞争但增加山杜英地上竞争,进而促进混交杉木生长15%,减少混交山杜英生长13%,混交林蓄积整体表现略微增产。土壤有效磷是影响杉木-山杜英混交林木生长的主要生境因子,杉木-山杜英混交效应随地位指数增加而减少,可能与根系成层性提升土壤磷素可利用性有关。研究结果为培育高产杉木-山杜英混交林的造林地选择奠定理论基础。
猜你喜欢
今日中国·西班牙文版(2021年4期)2021-04-09
农民致富之友(2020年8期)2020-05-11
森林工程(2018年4期)2018-08-04
现代园艺(2018年2期)2018-03-15
现代园艺(2017年23期)2018-01-18
现代园艺(2017年23期)2018-01-18
方圆(2018年23期)2018-01-07
广西林业科学(2016年2期)2016-03-20
广西林业科学(2016年1期)2016-03-20
广西林业科学(2016年4期)2016-03-16
