
鳜Shh基因分子特征和时空表达规律及其对肌肉生长的调控
2024-02-15 18:14孟杨杨鑫成嘉陈琳褚武英宾石玉张建社
水生生物学报 2024年1期
孟杨杨 朱 鑫成 嘉陈 琳褚武英宾石玉张建社*
(1.广西师范大学生命科学学院,珍稀濒危动植物生态与环境保护教育部重点实验室,桂林 541006;2.长沙学院生物与化学工程学院,水生动物营养与品质调控湖南省重点实验室,长沙 410022)
鱼类肌肉组织由明显分层的红肌和白肌组成,又可称为慢肌和快肌,它们既是鱼类躯干的结构组织和运动器官,也是人类重要的蛋白源[1]。已有研究证实,多种信号分子参与了慢肌和快肌发生分化的调节过程。这些信号分子主要包括三种类型,即Hedgehog[2]、Wnt[3]和TGF-β家族基因[4]。有关调节肌纤维决定分化的研究主要集Hedgehog和TGFβ家族基因,其研究对象也主要采用如斑马鱼等模式生物。Echidna hedgeho仅在脊索(Notochord)中表达,Currie等[5]研究发现过表达Echidna hedgehog,具有挽救无脊索突变体的肌肉先驱细胞(Muscle pioneer cells)分化的能力。此外,研究还发现,在野生型胚胎中联合异位表达Echidna hedgehog和sonic hedgehog能够诱导额外的肌肉先驱细胞的生成,这表明这两种信号依次作用于体节发育。Lewis等[6—8]采用Hedgehog家族蛋白及Hedgehog家族蛋白的负性调节因子Patched1蛋白处理斑马鱼早期胚胎,研究证实了Hedgehog家族蛋白在诱导斑马鱼慢肌形成中起关键的作用。进而探究发现在野生型斑马鱼胚胎过表达Shh或者显性失活的PKA(Dominant negative PKA)可导致所有的体节细胞发育为慢肌纤维[9,10]。Du等[11]结合Hedgehog和TGF-β家族成员基因以斑马鱼为研究对象开展了慢肌分化的机理研究。该研究揭示,在野生型斑马鱼胚胎过表达Hedgehog同源基因或抑制PKA可诱导慢肌肌源细胞的定向分化,而采用TGF-β家族成员基因Dorsalin-1处理斑马鱼胚胎可抑制肌原细胞的产生,说明Hedgehog和TGF-β家族成员基因在调控慢肌分化中是拮抗性的,即Hedgehog可诱导慢肌的形成,而TGF-β成员,如BMP4抑制慢肌发育。然而,目前有关Hedgehog信号分子在养殖鱼类特别是鳜诱导肌细胞或肌纤维类型定向分化的研究尚未有报道,有待深入研究。
Sonic hedgehog(Shh)信号通路内存在众多家族成员基因,目前所熟知的蛋白主要包括分泌性糖蛋白配体Shh、Patched(Ptch)与Smoothened(Smo)膜受体蛋白,以及核转录因子Gli等。Shh是一种分泌型蛋白,可经过细胞间的互作将Shh传递到受体细胞中与受体结合。Ptch属于12次跨膜蛋白,既是Shh信号通路中的受体,同时也是Shh信号的直接靶基因,并通过负反馈抑制Shh信号的活性[12]。在正常情况下,Ptch抑制Smo蛋白活性,从而抑制下游通路,随后下游的Gli蛋白在蛋白酶体(Proteasome)内被截断,并以羧基端被截断的形式进入细胞核内,抑制下游靶基因的转录。Smo属于7次跨膜结构蛋白,同时也是Shh信号分子传递所必须的受体,当Shh信号分子与Ptch结合时,解除Ptch对Smo的抑制作用,促使Gli蛋白进入核内激活下游靶基因转录。Gli作为Shh信号通路中发挥功能作用的核转录因子,在脊椎动物中有3种类型,即Gli1、Gli2和Gli3。其中Gli1只具有转录激活作用,转录激活Gli1后会正反馈调控更进一步增加Gli的活化。而Gli2与Gli3不仅具有转录激活作用,还同时具有转录抑制作用[13]。
鳜肌肉为高蛋白的优质肉类,较常见经济鱼类总氨基酸含量更高,肉质鲜美,深受广大消费者的喜爱,具有重要的经济价值,是我国淡水养殖的重要名贵特色优良品种[14—17]。本研究对鳜Shh基因进行了生物信息学分析,实时荧光定量PCR检测了Shh基因的表达特性。随后通过环巴胺处理鳜胚胎抑制Shh信号传导,分析了与肌细胞增殖和分化相关转录因子及肌球蛋白重、轻链等基因的表达水平。结果揭示,Shh信号分子被抑制后,所检测的相关基因表达均显著降低,表明Shh参与调控鳜肌细胞的早期分化和融合。该研究有助于我们从分子水平了解Shh信号分子的生物信息学特征,以及鱼类肌细胞分化的潜在机理,将丰富鱼类早期肌肉发生分化的基础理论,为健康养殖提供参考依据。
1 材料与方法
1.1 试验材料
用于实验的鳜胚胎及用于基因组织表达检测的鳜幼鱼均来自于实验室长期合作单位湖南省水产科学研究所。
1.2 实验方法
鳜不同发育阶段胚胎的收集胚胎孵育方法参考刘希良等[18]技术,收集未受精卵、卵裂期、囊胚期、原肠早期、神经胚期、视泡期、尾芽期、肌肉效应期、心脏搏动期、血液循环期、胸鳍原基期和出膜期胚胎材料,分别取40—50枚卵或胚胎保存于装有1 mL RNA保护液(Sample Protector for RNA/DNA,9750 宝日医生物,北京)的离心管中,并用液氮快速冷冻,最后保存于-80℃超低温冰箱待用。
环巴胺溶液浸泡胚胎样品收集称量10 mg环巴胺(Cyclopamine,HY-17024,MedChemExpress,美国)粉末溶于2.43 mL DMSO溶液中,得到10 mmol/L环巴胺母液。将环巴胺母液与曝气水按 1∶500 的比例稀释至环巴胺终浓度20 μmol/L (DMSO终浓度为0.2%)。待胚胎发育到囊胚期时,分别取100枚胚胎至环巴胺处理组和 DMSO 对照组 (浓度为 0.2%,不含环巴胺) 进行浸泡处理,统计两组死亡率并记录,待胚胎发育到出膜期时收集胚胎样品于2 mL离心管中,液氮速冻后,至-80℃储存备用。
鳜组织样品的收集将实验室养殖缸内暂养的鳜随机选取规格大小相一致且健康幼鱼3尾,体重为(250±10) g。参考吴萍等[19]安乐处理鱼类方式,处理后放置冰上解剖。分别取红肌、白肌、肝脏、心脏、脾脏、肾脏、脑和肠道放入2 mL EP管中,在液氮速冻后,迅速放入-80℃超低温冰箱储存备用。
RNA提取及cDNA模板合成鳜各时期胚胎及幼体组织总RNA提取参照RNAiso Plus (9108,宝日医生物,北京) 试剂盒说明书进行。提取胚胎总RNA后使用1.5%琼脂糖凝胶电泳及微量核酸蛋白测定仪(NanoPhotometer NP80,Implen GmbH,Germany)对RNA的完整性及浓度进行检测,核酸A260/A280比值在1.8—2.0确保核酸质量。并用逆转录试剂[MonScriptTMRTⅢ Super Mix with dsDNase(Two-Step),Monad,中国],按照试剂说明书进行RNA逆转录,获得的cDNA保存于-80℃冰箱备用。
引物设计与合成通过UCSC及NCBI网站查询实验所需鳜基因序列,利用NCBI网页中Primer-blast工具进行引物设计,引物序列由北京擎科生物科技有限公司合成 (表1)。

表1 荧光定量引物Tab.1 The Primers for RT-qPCR
实时荧光定量PCR以鳜胚胎及幼鳜组织样品的cDNA为模板,进行实时荧光定量PCR检测,每个样品进行4次技术重复检验。实验反应体系:SYBR Premix ExTaqTMⅡ 6 μL、正反向引物各0.5 μL、DNA模板1 μL、Nuclease-free water 4.5 μL。反应条件为: 95℃ 预变性1min;95℃ 变性5s;58℃ 退火25s;40个循环;绘制融解曲线的温度为65—95℃,每0.5℃ 读板1次。组织表达实验用Rpl13作为内参,胚胎时期表达实验用Gapdh作为内参。
数据处理及分析将实验所得数据利用Excel软件进行分析,各基因相对表达量使用2-ΔΔCt法计算,并采用SPSS 20软件对各组基因表达值进行统计分析。其中时序及组织表达结果使用单因素方差分析,环巴胺浸泡实验数据使用独立样本t检验。最终所得结果均以平均值±标准误表示,当显著性P<0.05 认为两组数据之间差异显著。
2 结果
2.1 鳜Shh基因生物信息学特征
Shh基因序列鉴定分析鳜Shh基因mRNA序列全长2407 bp,5′UTR长度为160 bp,3′UTR长度为1005 bp。起始密码子ATG在161—163 bp处,终止密码子TGA在1400—1403 bp处。开放阅读框长度为1242 bp,编码413个氨基酸。
Shh蛋白特性和结构预测通过在线Expasy-ProtParam软件工具预测鳜Shh蛋白,结果显示其由413个氨基酸组成,分子量为46.01 kD,等电点为6.57,蛋白偏弱酸性。预测不稳定指数(Ⅱ)计算能35.27,为稳定性蛋白。Shh蛋白中含有47个酸性氨基酸(Asp+Glu),44个碱性氨基酸(Arg+Lys)。该蛋白质预估半衰期为30h,脂溶系数为82.83,亲水性平均系数是-0.292,属亲水蛋白,疏水性不强。编码鳜Shh蛋白的20种氨基酸中,亮氨酸(Leu)最多,占比8.7%,半胱氨酸(Cys)最少,占比1.5%。通过TMHMM-2.0对蛋白质跨膜域进行分析,显示Shh蛋白存在一处跨膜域结构,该蛋白质属于膜结合蛋白。
采用NCBI保守结构数据库(CDD) 的CD-searcher在线工具预测,Shh蛋白含有2个功能结构域,即Hh-N和Hh-C结构域。Hh-N结构域中拥有Hh_signal模块,模块结构位于39—184位氨基酸处,编码146个氨基酸残基。Hh-C结构域的Hint模块位于186—399位氨基酸处,编码214个氨基酸残基。
鳜Shh同源蛋白比对及生物进化树分析在NCBI数据库中检索包括鱼类、两栖类、爬行类、鸟类和哺乳类等20个物种的Shh氨基酸序列,采用NCBI上Protein BLAST对蛋白同源性进行比对,结果显示,鳜Shh蛋白与鱼类Shh蛋白具有较高的同源性,其与长吻钻嘴鱼(Chelmon rostratus)和大黄鱼(Larimichthys crocea)Shh蛋白相似度最高,为97.58%;与其他物种相比,鳜Shh蛋白与家猫(Felis catus)Shh蛋白相似度最低,仅为61.17%。使用DNAMAN软件对不同物种间Shh蛋白Hh-N结构域的氨基酸序列比对,结果显示,其在鳜、大口黑鲈及白梭吻鲈等鲈形目鱼类中具有较高的相似性,说明Shh蛋白中组成Hh-N结构域的氨基酸序列在不同物种之间较为保守。使用MEGA11软件对Shh蛋白进行系统进化分析,结果显示(图1),鳜Shh与鱼类Shh聚为一支,其中,鳜和鲈形目白梭吻鲈的遗传距离最近。
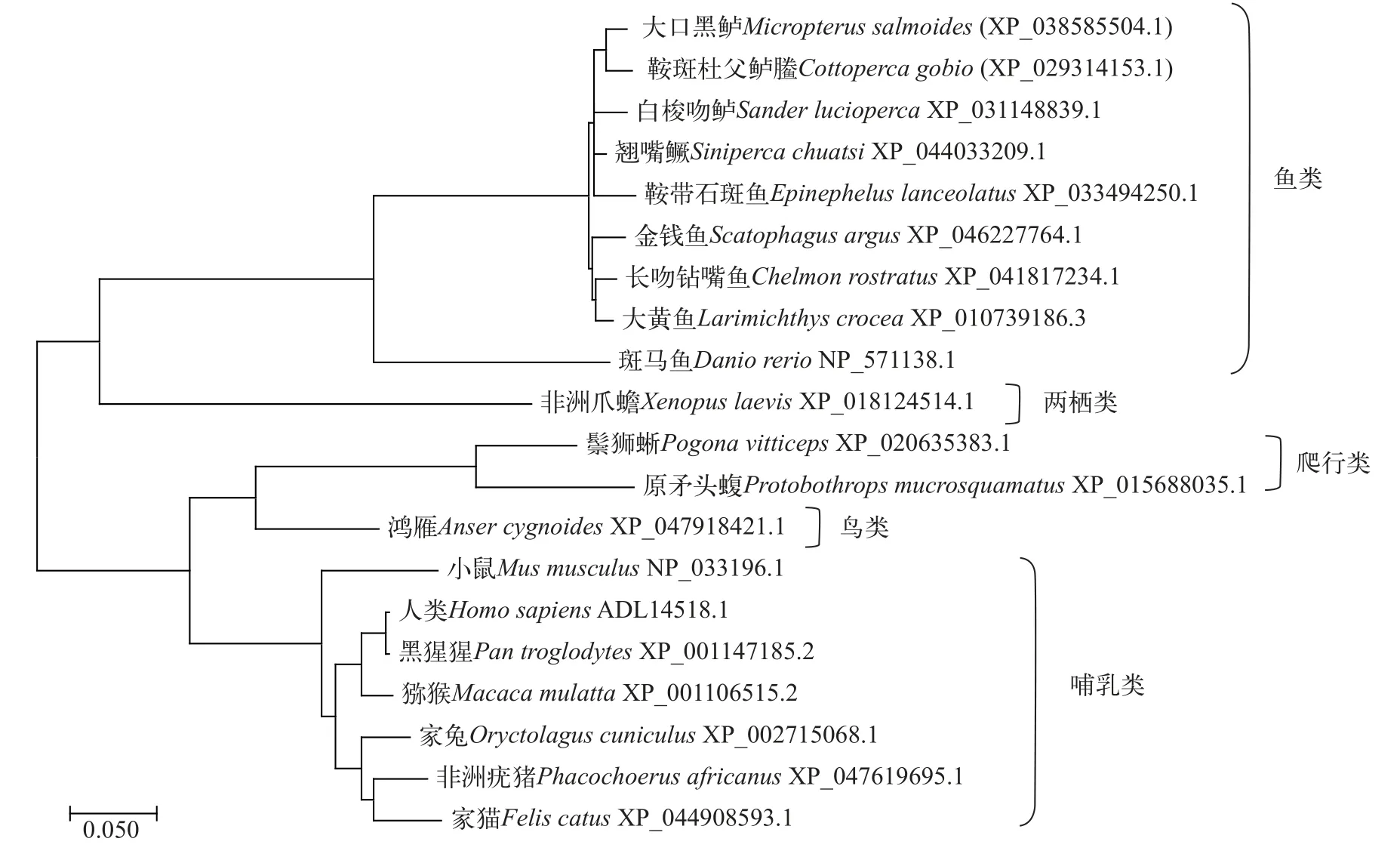
图1 Shh蛋白系统进化分析Fig.1 Phylogenetic analysis of the Shh protein
2.2 鳜Shh基因时空表达特征
Shh在鳜胚胎不同发育阶段的表达特征通过RT-qPCR检测Shh基因在鳜胚胎不同发育阶段的表达量,如图2所示,Shh在整个胚胎发育阶段均有不同程度表达,胚胎发育初期表达水平相对较低,随着胚胎发育进程推进,其在神经胚期表达显著上调(P<0.05)。Shh在胚胎发育中后期表达水平较高,并在出膜期达到峰值。
Shh在鳜不同组织中的表达特征通过RTqPCR检测分析Shh基因在鳜不同组织内的表达差异(图3),结果表明,在脑和肠道中显著高于在白肌、红肌、心脏、肾脏、肝脏和脾脏组织中的表达(P<0.05)。
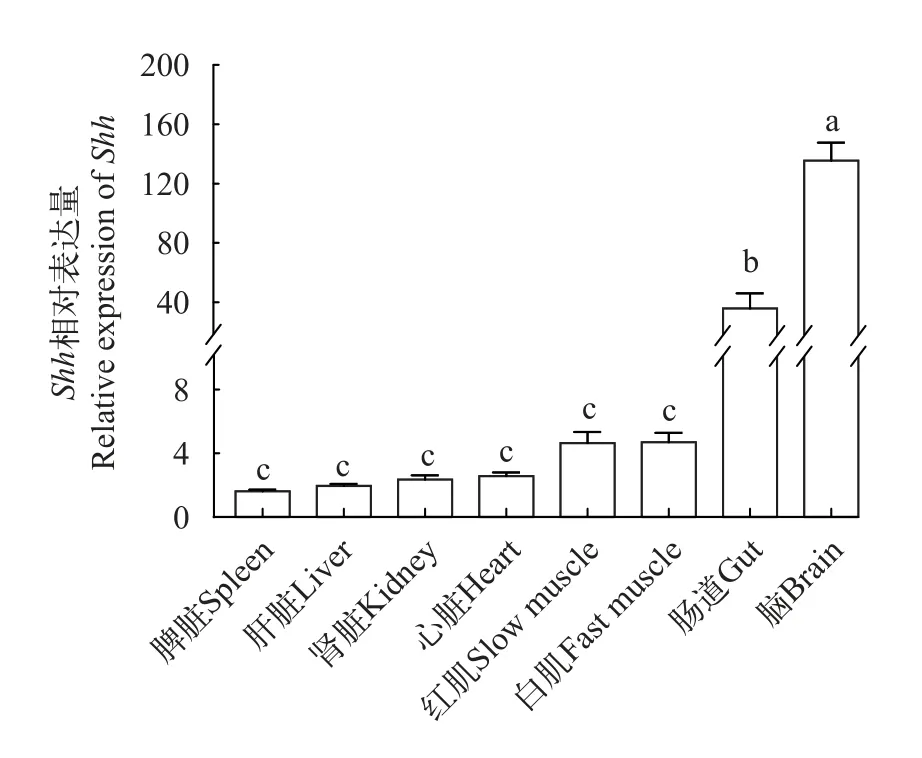
图3 鳜不同组织中Shh的相对表达丰度Fig.3 Relative expression abundance ofShhin different tissues ofSiniperca chuatsi
2.3 环巴胺抑制Shh信号通路对鳜胚胎发育及肌肉发育相关调控因子的影响
环巴胺浸泡鳜胚胎对发育的影响环巴胺作为一类Shh信号分子的特异抑制剂,实验采用浸泡方式处理鳜胚胎,并采用DMSO浸泡作为对照处理。实验期间死亡率统计结果为,实验组死亡23枚,对照组死亡18枚,并利用SPSS 20进行卡方检验,无显著性差异(P>0.05)。如图4所示,对照组存活的82枚胚胎躯干全部正常,实验组存活的77枚胚胎躯干尾端全部呈弯曲状,致畸率100%,推测可能是由于其尾部肌肉组织发育异常,导致胚胎畸形。
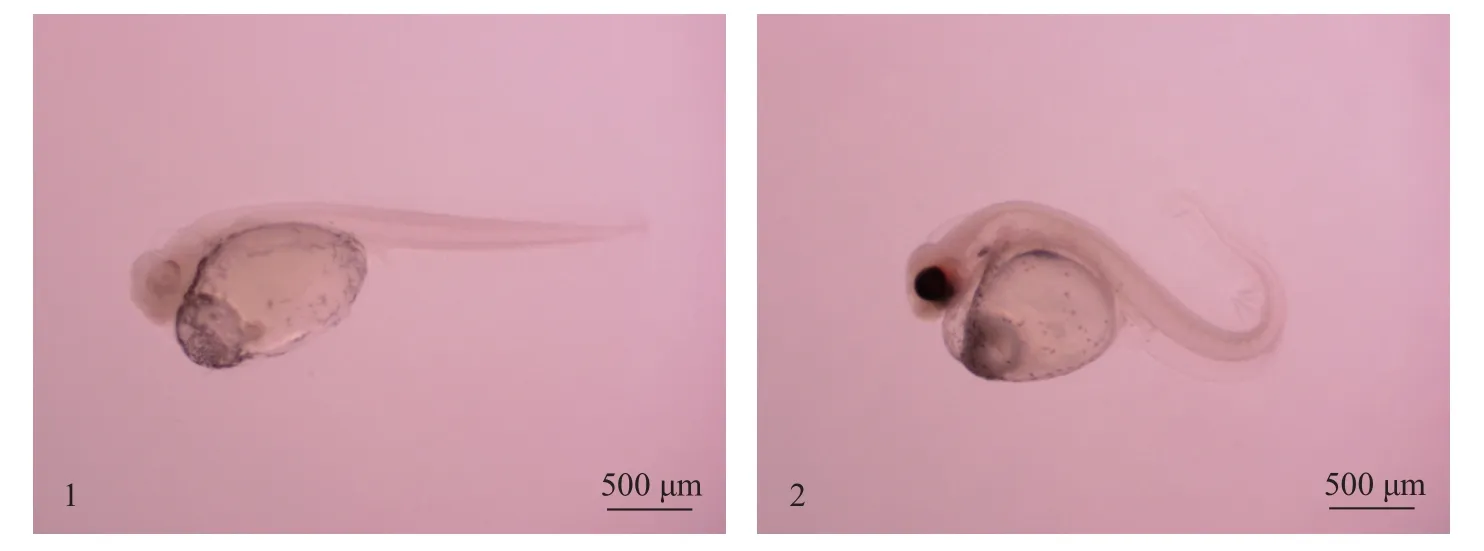
图4 浸泡处理后鳜胚胎呈现的发育表型Fig.4 Developmental phenotype ofSiniperca chuatsiembryos after immersion treatment
环巴胺处理对Shh通路下游基因及生肌调节基因表达影响通过RT-qPCR检测Shh信号通路下游靶基因Ptch1、Gli1、Gli2和Gli3的表达量,评判Shh信号传递受阻情况。随后检测与肌细胞增殖和融合相关基因(Pax3、Pax7和Myomaker)及生肌调节因子(MRFs)家族成员,如Myod、Myog、Mrf4和Myf5基因的表达量。如图5所示,经环巴胺浸泡处理的鳜胚胎,所检测基因表达量较对照组均显著下降(P<0.05)。
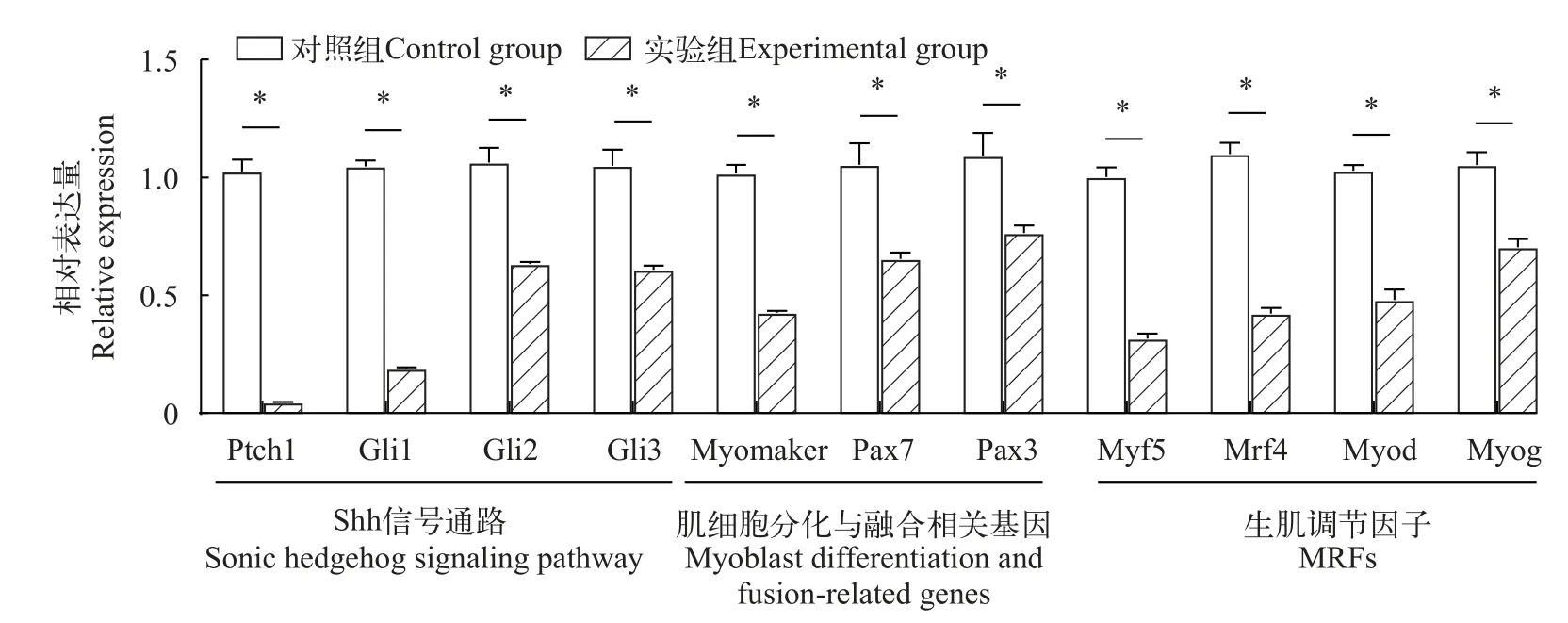
图5 鳜出膜期Shh通路下游靶基因、肌细胞分化与融合相关基因、生肌调节基因MRFs家族成员基因的相对表达丰度Fig.5 Relative expression abundance of downstream target genes of the Shh pathway,genes related to myoblast differentiation and fusion,and genes of members of the MRFs family of myogenic regulatory genes inSiniperca chuatsiat the emergence stage
环巴胺处理对鳜胚胎肌球蛋白编码基因表达影响为探讨环巴胺处理对鳜肌肉生长的影响,通过RT-qPCR检测分析肌球蛋白轻链及重链基因在鳜出膜期表达量。如图6所示,与对照相比,肌球蛋白轻链及轻链激酶基因的同分异构体包括Myl2、Myl3、Myl6、Mylk和Mylk4基因的表达量均显著降低,肌球蛋白重链的7个同分异构体如Myh1、Myh2、Myh3、Myh7、Myh9、Myh10和Myh11基因的表达量同样显著下降(P<0.05)。
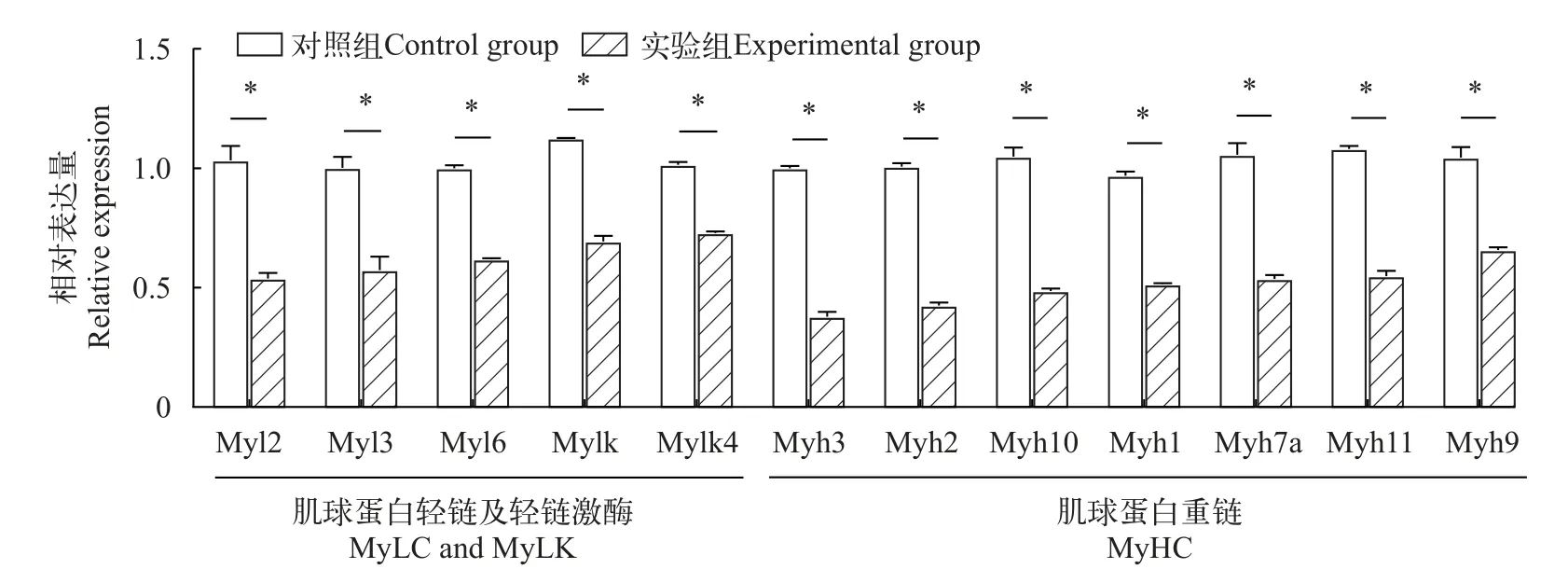
图6 鳜出膜期肌球蛋白及激酶基因表达丰度Fig.6 Myosin and kinase gene expression abundance in the emergence phase ofSiniperca chuatsi
3 讨论
3.1 鳜Shh基因编码蛋白的结构特征分析
本研究对鳜Shh蛋白结构和同源进化特征进行生物信息学分析,结果显示,鳜Shh开放阅读框为1242 bp,编码413个氨基酸,分子量为46.01 kD,等电点为6.57,蛋白偏弱酸性;鳜Shh蛋白脂溶系数为82.83,为脂溶性蛋白,亲水性平均系数是-0.292,有一处跨膜结构,推定为亲水性膜结合蛋白。该基因编码的蛋白含有两个功能结构域,即Hh-N与Hh-C。预测得出的Hh_signal模块与Hint模块分别属于氨基端结构域和羧基端结构域。有研究报道,果蝇与脊椎动物Hh前体蛋白通过C-末端部分催化并自我分割后变为N端与C端,果蝇Hh-N过表达会导致胚胎表皮特征发生改变,Hh-C过表达则无任何变化[20—22]。在脊椎动物中Shh-N可诱导运动神经元和基底细胞分化[23]。以上说明Hh-N结构域具有Hh蛋白信号活性。Hh-C的功能则是起到胆固醇转移酶类作用,影响信号分子的分布[24,25]。
同源进化特征分析显示,鳜Shh蛋白与长吻钻嘴鱼和大黄鱼的Shh蛋白相似度最高,与家猫Shh蛋白相似度最低,且物种间Shh蛋白Hh-N结构域的氨基酸序列较为保守。鳜Shh蛋白系统进化分析显示,鳜Shh蛋白与鱼类Shh蛋白聚为一支,并与鲈形目白梭吻鲈遗传距离最短,亲缘关系最近,说明Shh在鱼类进化过程中进化较为保守。
3.2 鳜Shh基因时空表达特征分析
有研究表明,Shh基因在动物胚胎发育过程中,对神经、消化道及肢芽等组织发育起到重要调控作用[26]。尤其在脊椎动物早期胚胎发育过程中,脊索或神经底板分泌的Shh对背侧肌节部位的肌细胞形成和存活起着至关重要的作用[27—29]。本研究采用RT-qPCR对翘嘴鳜12个胚胎发育时期Shh的表达进行分析,结果表明Shh在胚胎不同发育阶段都有表达且具有显著性差异,并随胚胎发育进程的推进,表达量在神经胚期显著增高,且整个胚胎发育中后阶段表达量保持较高水平,最终在出膜期表达量达到峰值。因此,我们推测Shh可能对鳜胚胎神经的发育发挥作用。
有研究报道,Shh基因在不同动物的多组织中表达[30]。本次研究对鳜不同组织表达分析揭示,Shh在检测的所有组织中均有表达,脑和肠道中表达较高,在白肌和红肌等组织中表达相对较低。有研究表明,Shh作为一种分泌蛋白,在海马体神经元回路的形成和可塑性方面发挥重要作用,其中海马神经元树突中的Shh受体激活后参与跨神经元信号通路,该通路加速了神经轴突生长[31]。因此,我们推测Shh可能在幼体鳜的脑神经发育中也有着重要的调控作用。有研究发现,Shh在脊椎动物体内是通过作用于肌肉干细胞直接调控肌肉的生长发育,以及促进新的肌纤维数量增加[32]。Koleva等[33]对小鼠肌肉干细胞进行体外培养,发现Shh能够促进肌肉干细胞增殖。因而推测Shh可能在鳜肌细胞生长过程中起到调控作用。
3.3 鳜Shh基因对肌肉发育相关调控因子的表达分析
本研究为探究Shh在鳜肌肉发育中的作用,采用Shh信号通路特异性拮抗剂环巴胺对鳜鱼胚胎进行浸泡处理。环巴胺在Shh信号通路中的作用机制是抑制Smo的活性,阻断Shh信号的传递[34]。本次研究检测Shh通路内基因(Ptch1、Gli1、Gli2和Gli3)、生肌调节因子家族基因、调控肌肉发育及相关肌球蛋白重链、轻链及轻链激酶基因的表达。结果显示,Ptch1、Gli1、Gli2和Gli3表达量都显著降低,说明环巴胺处理后,该通路内的基因都受到不同程度的抑制。
Pax3、Pax7、Myf5、Myod、Myog和Mrf4等转录因子,它们协同诱导肌源性干细胞的激活、增殖和定向分化直至形成成熟肌纤维[35]。其中Pax3和Pax7在肌肉干细胞中特异表达,并在调控肌肉干细胞增殖和分化中起重要作用[36]。Pax3、Pax7、Myf5及Myod的表达是控制肌肉干细胞激活、增殖与分化的关键要素[37]。另外,Myomaker是一个近期新发现的促使肌细胞融合的重要基因,属于tmem 8c家族成员,是一种跨膜蛋白[38]。Myomaker体内表达实验证实,该基因在肌细胞融合过程发挥关键作用[38],在斑马鱼中,Myomaker基因在快肌中表达,敲除Myomaker后导致肌细胞融合缺陷[39—41]。同时,过表达Myomaker足以诱导快肌细胞之间的过度融合,同时还促使通常无法融合的慢肌细胞融合,说明该基因在鱼类肌细胞融合中有重要作用[42]。通过环巴胺处理鳜胚胎抑制Shh信号通路后,对参与调控肌细胞分化的转录因子及肌细胞膜融合基因进行RT-qPCR检测,发现Pax3、Pax7、Myf5、Myod、Myog、Mrf4和Myomaker的表达均显著下降,推测Shh可能通过直接或间接调控生肌调节因子等基因表达,影响鳜胚胎肌肉细胞的发生与分化。
我们先前在鳜肌纤维中鉴定了多种肌球蛋白重链和轻链亚型,其中,肌球蛋白是构成鱼类肌肉的主要结构蛋白,是一种高度不对称且分子量约500 kD的六聚体蛋白,由两条分子量约230 kD的重链和四条分子量约16—20 kD的轻链组成[43,44]。另外,研究表明,肌球蛋白重链亚型如Myh1、Myh7a、Myh9、Myh10和Myh11基因在慢肌中表达成倍高于快肌,而Myh2和Myh3的表达量在快肌中的表达水平是慢肌的数倍;肌球蛋白轻链及轻链激酶亚型如Myl2、Myl3、Myl6、Mylk和Mylk4等基因在慢肌中的表达是快肌的数倍[19]。本实验结果表明,当Shh信号传递受阻后,肌球蛋白重链、轻链及轻链激酶亚型基因表达量均显著降低表达。由此推测,鳜胚胎发育畸型的原因是环巴胺抑制剂处理导致Shh信号分子传递阻断,继而抑制肌细胞分化相关基因的表达。因此说明,Shh对鳜肌肉细胞分化和生长发挥重要调控作用,但其调控的分子机制仍有待进一步深入研究。
猜你喜欢
食品科学(2023年4期)2023-03-06
广州大学学报(自然科学版)(2019年1期)2019-05-07
天津科技大学学报(2016年1期)2016-02-28
湖北师范大学学报(自然科学版)(2015年2期)2016-01-10
中国当代医药(2015年33期)2015-03-01
郑州大学学报(医学版)(2015年2期)2015-02-27
心血管病学进展(2015年4期)2015-02-22
现代检验医学杂志(2015年2期)2015-02-06
重庆医学(2014年36期)2014-03-04
中国病理生理杂志(2012年8期)2012-03-17
