
基于稳定同位素技术的洈水水库鱼类群落营养结构
2024-02-15 18:14:16姚鸿伟林子木陈敏建钟可儿马吉顺周
水生生物学报 2024年1期
姚鸿伟 林子木 陈敏建 钟可儿马吉顺周 琼
(1.华中农业大学水产学院,农业农村部淡水生物繁育重点实验室,武汉 430070;2.长江经济带大宗水生生物产业绿色发展教育部工程研究中心,武汉 430070)
大水面渔业是我国淡水渔业的重要组成部分,在建设水域生态文明、保障优质水产品供给、推动产业融合、促进渔民增收等方面发挥着举足轻重的作用[1—3]。截至2020年,大水面增养殖的面积和产量分别达到了全国淡水养殖总面积和总产量的42.80%、12.40%,其中水库养殖面积14165.69 km2,大水面养殖面积为21866.62 km2[4],水库养殖面积占大水面增养殖总面积64%以上。然而,很多湖泊和水库的天然饵料资源的利用并不充分[3],熊满堂等[5]、邵旭东等[6]利用浮游生物的生物量分别对丹江口水库、打虎石水库的鲢(Hypophthalmichthys molitrix)、鳙(Hypophthalmichthys nobilis)生物量进行估算,发现水库全年浮游生物可提供的鲢、鳙渔获量远高于实际渔获量,表明丹江口水库和打虎石水库当前的浮游生物利用程度较低,尚有较大的渔业增殖潜力。研究食物网营养结构有助于理解初级食物源传递给高营养级水生动物的物质循环和能量流动过程,为实现湖泊、水库天然饵料的合理利用提供理论依据,这将为处于关键转型期的大水面渔业结构的优化提供建设性意见。
食物网研究能阐明生物群落结构和物种间复杂的营养关系,揭示生态系统中物质循环和能量流动的过程,是淡水生态学和水域生态系统研究的重点方向[7]。鱼类群落营养结构是构建食物网的主要部分,研究鱼类群落营养结构不仅能揭示各生物物种在能量流动过程中较为稳定的摄食与营养关系[8],还能为渔业资源的管理及可持续利用提供依据[9,10]。针对水域生态系统不同生物的食性分析是构建食物网结构的主要途径,食性分析途径主要有传统的肠含物分析法和近年来逐渐兴起的稳定同位素技术[11]。由于肠含物分析只能反映鱼类被捕获前短时间内的食物来源,观察到的肠含物主要是没有消化完全的食物[12],因此具有相对较大的偶然性。相比而言,稳定同位素技术可弥补肠含物分析法的这些不足,反映鱼类长期的摄食组成,而且能够较为准确地评估其营养位置[13],能够对小型鱼类或底栖动物等不容易通过胃含物方法进行分析的物种进行营养级和潜在食物来源分析。目前有关河流的食物网碳源利用情况的研究较多,但水库食物网碳源利用情况的研究报道较少[14]。王晨等[15]利用贝叶斯混合模型分析得出,巴松措水生无脊椎动物和鱼类表现出对河岸植物或陆地树叶的依赖,湖泊水量的增加也有助于推动水体消费者对陆地碳源的利用[16]。碳源作为食物网的基础能量来源,厘清洈水水库的碳源利用情况有助于理解水库的物质循环与能量流动过程[17]。
洈水水库位于湖北省松滋市洈水镇,地处洈水流域的中游,与湖南澧县接壤。水库流域面积为1142 km2,总库容5.11×108m3,是一座以灌溉为主,兼有防洪、发电、养殖、航运、旅游、城乡生产和生活供水等综合利用的大(Ⅱ)型水库。2012年以来,洈水水库以水产业为重点,量身打造了“洈水有机鱼”品牌,其中以鳙和翘嘴鲌(Culter alburnus)尤为著名。目前,洈水水库的网箱养殖已被全面禁止,转而实行更加健康的大水面生态养殖模式。迄今为止,洈水水库渔业结构和鱼类对食物资源利用的研究仍为空白,本研究应用碳(δ13C)、氮(δ15N)稳定同位素技术研究洈水水库夏季与秋季鱼类群落的营养结构特征及其差异,运用贝叶斯混合模型评估主要鱼类对不同碳源的利用情况,旨在为洈水水库渔业结构的调整提供科学依据。
1 材料与方法
1.1 实验设计与数据来源
本研究于2020年6—11月在洈水水库进行了鱼类、水生生物等样品采集,并同时进行了水环境分析。夏季(6—8月)为蓄水期,秋季(9—11月)为排水期。夏季降雨频繁导致水库的水位较高、水体流速快,秋季降雨量大幅度减小,水库排水导致水位较低。根据洈水水库的地理形态特征共设置10个采样站点(图1),对基础食物源和同位素样品进行采集。

图1 洈水水库采样站位图Fig.1 Sampling sites of the Weishui Reservior
1.2 研究方法
稳定同位素样品的采集与处理在洈水水库10个采样点逐月采集周丛藻类、陆生植物、颗粒有机物(Particulate organic matter,POM)及底栖动物和鱼类肌肉样品,用于稳定同位素分析。
由于洈水水库的水位常年保持在较高的水平,库区水生植物稀少,库区岸边的石头、浮木等自然基质也较少,不利于周丛藻类的附着与生长,因此本研究通过放置人工基质来采集周丛藻类。采样前20—30d在水体中提前放置人工基质,每次采完样后将人工基质投放回水中。用硬毛刷将附在基质上的藻类刷入装有蒸馏水的白磁盘后转移至1 L采样瓶内,带回实验室,通过在450℃预烧的Whatman GF/F纤维过滤,滤膜于60℃烘干48h至恒重。POM样品在各采样点采集表、底层水混合,通过预先灼烧的Whatman GF/F纤维滤膜过滤获得,于60℃ 烘干48h至恒重。常见的陆生植物采于洈水水库周围10 m范围内的森林,大型植物仅收集叶片,小型植物全株采集,通过塑料自封袋保存带回实验室,鉴定后用蒸馏水冲洗干净,于60℃ 烘干至恒重,放于干燥器中保存。水生昆虫放置于充分曝气的蒸馏水中过夜,使其肠含物排空,整个个体作为分析样品,螺类去壳仅仅取其足部肌肉,在60℃下烘干至恒重。螺类、摇蚊幼虫等底栖动物清洗后烘干至恒重,干燥保存。鱼类样品通过3层定置刺网捕获后,测量记录体长(mm)和体重(g),取其背部白肌肉,用去离子水冲洗后于60℃烘干48h至恒重,放于干燥器中保存。
生物样品的稳定同位素分析POM与周丛藻类等滤膜样品先通过1 mol/L的盐酸酸化处理24h,去除可能影响δ13C测定的碳酸盐,然后在烘箱中60℃下恒温烘干48h,取出滤膜样品放入研钵中,用研棒和研磨至可以通过60目筛网,将研磨的粉末转移至EP管,储存于干燥器待测。鱼类取其背部白肌肉,用去离子水冲洗后在60℃条件下烘干48h至恒重,置干燥器中保存。处理时取出干燥的鱼类肌肉放入研钵中,用研棒磨至可以通过60目筛网,通过称量纸将研磨的粉末转移至EP管中,置于干燥器中保存待测。螺类和水生昆虫等水生无脊椎动物与有机碎屑采用相同的处理方法。植物样品用剪刀剪成碎屑后与鱼类肌肉样品进行相同的处理。所有样品研磨成粉末后干燥保存。全部样品送往中国科学院水生生物研究所通过Flash EA-1112 HT元素分析仪和Delta V Advantage同位素比质谱仪(Thermo Fisher Scientific,Inc.,美国)联合进行分析。分析碳(δ13C)和氮(δ15N)同位素的参照物质分别是VPDB (Pee Dee Belemnite)和空气中纯净的N2。标准物质分别是国际上通用的IAEA-USGS24和IAEA-USGS25。分析结果表示为δ13C和δ15N:
式中,R代表13C或15N,X代表13C/12C或15N/14N。
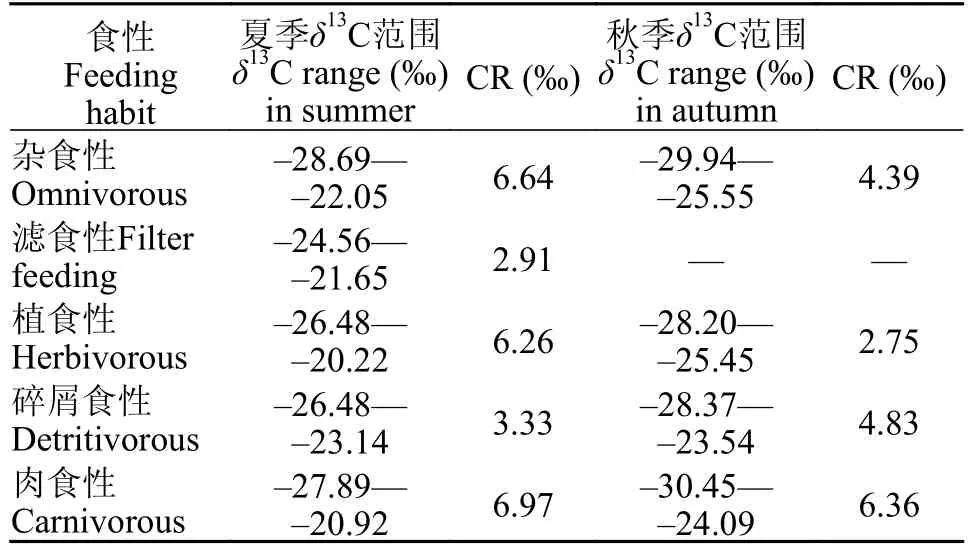
表1 不同食性鱼类在夏季和秋季的稳定碳同位素范围比较Tab.1 Comparison of theδ13C range of fish with different feeding habits in summer and autumn
根据某生物对基线生物氮稳定同位素比值的相对值,计算该生物的营养级(Trophic position,TP)。营养级计算公式如下:
式中,δ15Nsample为测定样品的氮稳定同位素值;δ15Nbaseline为洈水水库生态系统中基准生物的氮稳定同位素值;λ是基准生物的营养级。摇蚊幼虫是洈水水库常年存在的优势初级消费者种类之一,本文以摇蚊幼虫为基准生物(λ=2);Δm为消费者与饵料间的同位素富集度,为 3.4‰[18]。
通过R软件中的SIAR(Stable Isotope Analysis in R)软件包,运用贝叶斯混合模型估算可能的食物源对每种消费者的碳源贡献率[15]。在本研究中,我们使用稳定同位素值(δ13C和δ15N)和siarmcmcdirichletv4指令来估算不同季节食物源对消费者的碳源贡献率(取平均值、最小值和最大值)。
鱼类群落结构分析通过R软件中的SIBER软件包计算鱼类群落营养结构指标,本研究参考Layman等[19]对营养结构进行量化的6个指标: 氮值范围(δ15N range,NR),由水域消费者δ15N富集最高的值减去富集最低的值所得;碳值范围(δ13C range,CR),由水域消费者δ13C富集最高的值减去富集最低的值所得;生态位总面积(Total area,TA),指δ13C—δ15N双标图中所有消费者围成的凸多边形面积,反映食物网中生物对资源的利用情况;平均离心距离(Mean distance to centroid,CD)指各消费者距离中心点(消费者δ13C和δ15N平均值)距离的平均值,反映营养多样性的平均水平;平均最邻近距离(Mean nearest neighbor distance,NND)通过计算δ13C—δ15N双坐标图中每种消费者和距离其最近的消费者之间距离的平均值所得,反映物种聚集的整体密度;平均最邻近距离标准差(Standard deviation of nearest neighbor distance,SDNND)通过计算每种消费者和距离其最近的消费者之间距离的标准差所得,表示营养生态位分布范围水平,反映物种聚集的均匀程度。鱼类食性的确定参照Fishbase鱼类数据库和《四川鱼类志》[20]。
2 结果
2.1 鱼类与可利用食物源的稳定同位素比值(δ13C和δ15N)
夏季洈水水库共采集鱼类13种,各种鱼类δ13C值和δ15N值分别为-28.69‰—20.22‰与6.34‰—12.80‰(图2)。其中δ13C最小值为-28.69‰(黄颡鱼Tachysurus fulvidraco),最大值为-20.22‰(团头鲂Megalobrama amblycephala);δ15N最小值为6.34‰(黄尾鲴Xenocypris davidi),最大值为12.80‰(黄颡鱼)。水生无脊椎动物有摇蚊幼虫和水蚯蚓两种,其δ13C值和δ15N值分别为-37.96‰— -27.29‰与6.11‰—7.70‰。
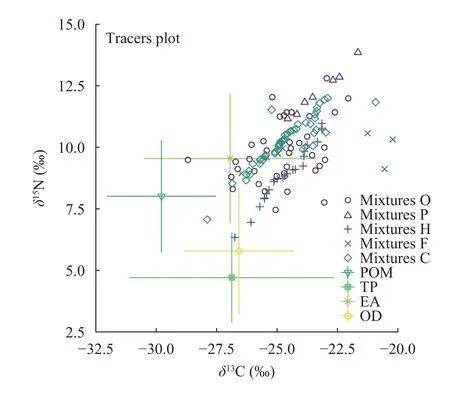
图2 基于碳、氮稳定同位素技术的洈水水库夏季食物网结构Fig.2 The characteristics of food web structure in the Weishui Reservoir in summer based on stable carbon (δ13C) and nitrogen(δ15N) isotopes
秋季洈水水库采集鱼类10种,其δ13C值和δ15N值分别为-30.45‰— -23.54‰与6.52‰—12.42‰(图3)。其中δ13C最小值为-30.45‰(蒙古鲌Culter mongolicus),最大值为-23.54‰(黄尾鲴);δ15N最小值为6.52‰(黄尾鲴),最大值为12.42‰(蒙古鲌)。水生无脊椎动物有摇蚊幼虫与水蚯蚓两种,其δ13C值和δ15N值分别为-45.98‰— -31.39‰与4.40‰—5.61‰。
根据图2和图3所示,蒙古鲌、翘嘴鲌和鳜(Siniperca chuatsi)等肉食性鱼类在夏季和秋季均位于食物网的顶端,其次是一些杂食性偏肉食的鱼类,如花䱻(Hemibarbus maculatus)。杂食偏植食性鱼类(鲤Cyprinus carpio、鲫Carassius auratus)和植食性鱼类(团头鲂)在图中处于较低的位置。
2.2 不同食性鱼类的碳源利用
夏季鱼类的稳定碳同位素为-28.69‰— -20.22‰,而秋季鱼类的稳定碳同位素为-30.45‰— -23.54‰,整体上夏季不同食性鱼类的稳定碳同位素值范围更大(表1)。按照鱼类的食性进行分析,杂食性鱼类、植食性鱼类和肉食性鱼类在夏季的碳值范围较大,分别为-28.69‰— -22.05‰、-26.48‰—-20.22‰和-27.89‰— -20.92‰;而碎屑食性鱼类的碳值范围在秋季较大,为-28.37‰— -23.54‰。杂食性、植食性、碎屑食性和肉食性鱼类在夏季和秋季的CR值分别为6.64与4.39、6.26与2.75、3.33与4.83、6.97与6.36。
贝叶斯混合模型分析表明,洈水水库夏季和秋季鱼类利用的主要碳源均为外源性碳。4种基础食物源中陆生植物和有机碎屑属于外源性碳,周丛藻类属于内源碳,POM包括内源性营养物质和外源性营养物质。夏季陆生树叶对鱼类的贡献最大,鱼类对陆生植物的平均利用率达52.85%,POM、周丛藻类和有机碎屑对鱼类的贡献率分别为10.00%、21.30%和15.85%。总体上,4种食物源对鱼类的贡献率为陆生植物>周丛藻类>有机碎屑>POM(图4)。秋季陆生树叶对鱼类的贡献率下降明显(31.36%),但有机碎屑的贡献比例显著升高(29.90%)。POM和周丛藻类的贡献率均有一定程度的升高,分别为13.52%、25.23%。秋季4种食物源对鱼类的贡献率为陆生植物>有机碎屑>周丛藻类>POM(图5)。
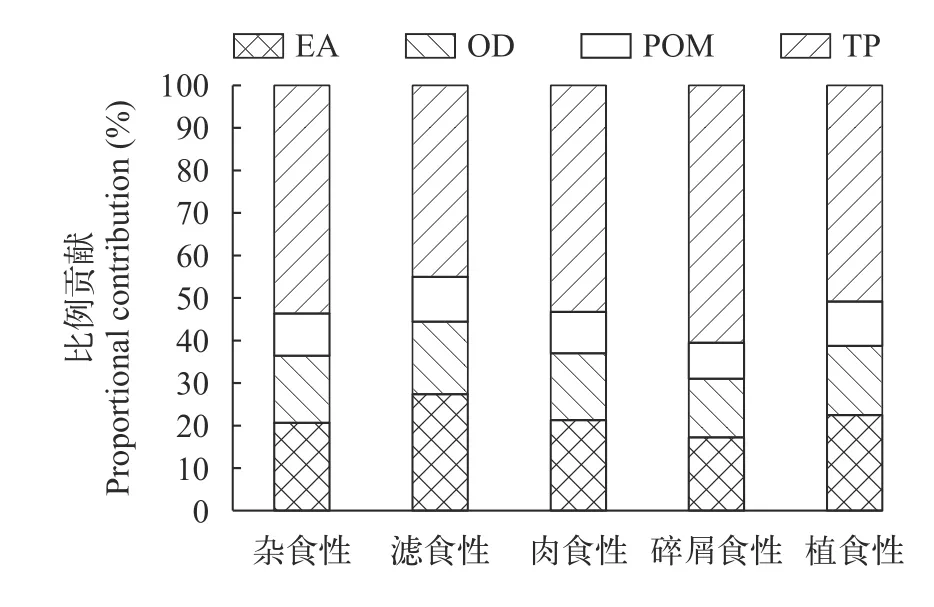
图4 夏季基础食物源对主要鱼类的贡献比例Fig.4 The proportional contribution of basal food sources to main fish species in summer
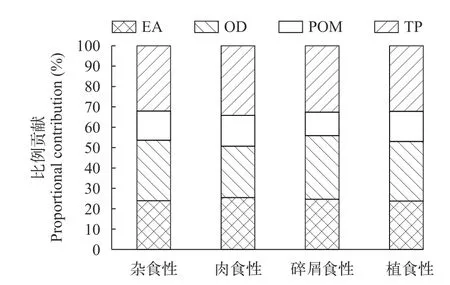
图5 秋季基础食物源对主要鱼类的贡献比例Fig.5 The proportional contribution of basal food sources to main fish species in autumn
2.3 不同食性鱼类的营养级
在本研究中,我们以摇蚊幼虫作为基线生物评价了17种不同食性鱼类的营养级(图6),其中夏季和秋季分别有14和13种,两个季节均有采集的鱼类为9种。夏季鱼类营养级为2.5—3.7,平均3.01。秋季鱼类营养级为2.9—3.8,平均3.32。鱼类食性分析显示,在夏季杂食性偏植食性鱼类、碎屑食性鱼类所处的营养位置较低(2.3—2.8),如鲤和黄尾鲴分别为2.31和2.65。肉食性鱼类所处的营养位置较高(2.8—3.4),其中鳜和翘嘴鲌的营养级分别为3.20和3.34。植食性和杂食偏肉食性鱼类所处的营养位置居中(2.7—3.0),如团头鲂和䱗的营养级分别为2.97和2.92。秋季植食性和杂食性鱼类所处的营养位置较低(2.9—3.4),如鲤和团头鲂的营养级分别为2.91和3.08;碎屑食性鱼类所处的营养位置居中,黄尾鲴的营养级为3.20;秋季肉食性鱼类所处的营养位置最高(3.4—3.8),其中鳜营养级为3.77。
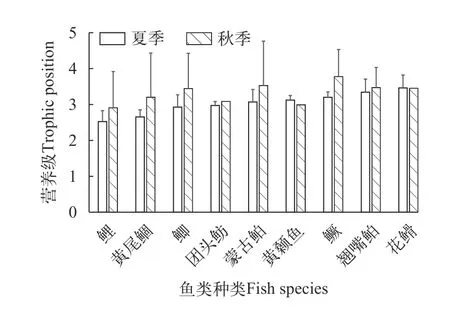
图6 洈水水库夏季和秋季主要鱼类的营养级Fig.6 Trophic positions of main fish species in summer and autumn
2.4 夏季与秋季的鱼类群落营养结构
基于稳定同位素混合模型(SIAR)评价了洈水水库夏季和秋季鱼类群落的营养结构(图7),主要通过SEAc、TA、CR、CD、MNND、NR和SDNND等7个指标反映鱼类群落的营养结构。夏季的鱼类群落核心生态位空间(SEAc)、生态位总空间(TA)和基础食物来源(CR)、营养多样性(CD)和鱼类群落的整体密度(MNND)、食物链长度(NR)和鱼类群落营养生态位分布范围(SDNND)均高于秋季(表2)。
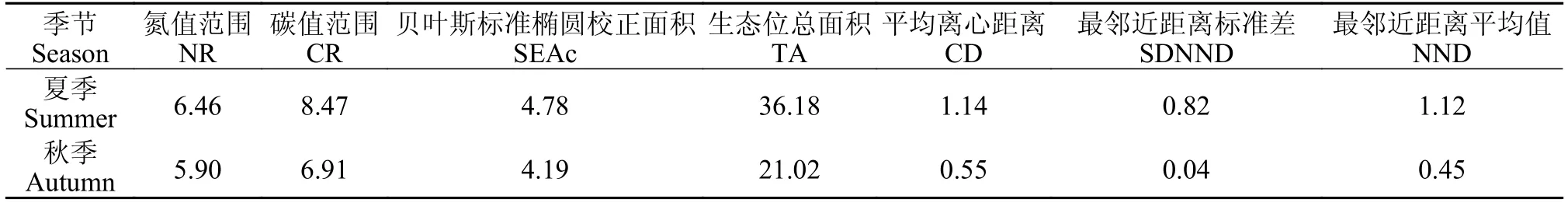
表2 基于稳定同位素混合模型的鱼类群落营养结构Tab.2 Trophic structure of fish communities based on stable isotope mixing model
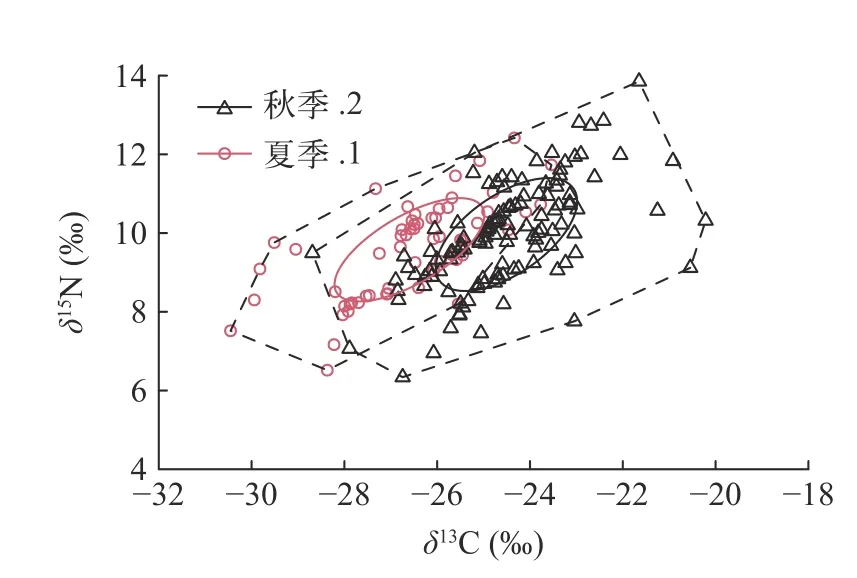
图7 基于贝叶斯稳定同位素混合模型的夏季和秋季主要鱼类群落的营养结构Fig.7 Trophic structure of main fish communities in summer and autumn based on the Bayesian stable isotope mixing model
3 讨论
3.1 洈水水库夏季和秋季鱼类的碳源利用
碳稳定同位素(δ13C)在相邻两个营养级之间能产生0—1‰的同位素分馏,我们把δ13C的范围(CR)作为衡量动物食物来源广泛程度的指标[21,22]。CR值越大,代表其食物来源越广泛[23]。本研究中夏季的CR值高于秋季,表明夏季鱼类的食物来源要比秋季更广泛。杂食性鱼类在夏季摄食植物性饵料,而在秋季更倾向于摄食动物性饵料,如水蚯蚓、摇蚊幼虫。夏季水库的蓄水导致大量湿生植物被淹没,为植食性鱼类和杂食性鱼类提供了丰富的食物来源[24]。而在秋季,陆生树叶变得枯黄,不适合作为植食性鱼类的食物源,同时秋季降雨量减少与水库排水等原因导致水库水位显著降低,因此植食性鱼类在秋季的食物来源没有夏季丰富,且秋季陆生植物对植食性鱼类和杂食性鱼类的贡献亦显著降低。秋季水库周边森林产生大量的落叶并随地表径流进入水体,落叶也会在微生物的作用下被分解为有机碎屑[25],参与水域生态系统的营养循环,为碎屑食性鱼类提供了丰富的食物来源[26,27],这可能是秋季有机碎屑对鱼类的贡献显著升高的原因。肉食性鱼类在两个季节的食物资源比较专一,均为小型鱼类和虾,其碳值范围没有显著的季节差异(P>0.05)。周丛藻类作为水生态系统中重要的内源性碳源,虽然其在两个季节对鱼类的贡献不大,但其对水体消费者的支持作用不可忽略[14]。秋季周丛藻类对鱼类的贡献比例略高于夏季,夏季频繁降雨将大量泥沙带入水库,水体的透明度下降明显。库区排水水流较快,水体频繁交换导致底层的泥沙再悬浮至水体表层,这也是水体透明度降低的重要原因,而透明度和流速是限制藻类生长的重要因素[10,29,30],因此夏季周丛藻类对鱼类的贡献率略低于秋季。外源性碳在夏季和秋季均为支撑洈水水库食物网的主要碳源,但其利用模式在两个季节存在明显的差异(P<0.05)。Wang等[31]利用IsoSource混合模型发现陆生植物是支持长江中游水生食物网的主要碳源。陆生植物和有机碎屑是水库鱼类主要的外来碳源,但鱼类对陆生植物和有机碎屑的利用比例存在差异。夏季洈水水库水位较高,两岸大量的植被覆盖和河床的类型(淤泥)都促进了水生无脊椎动物和鱼类对陆地来源的有机物质的利用,因此陆生碳源(陆生C4植物)可以从初级消费者转移到更高营养级的鱼类[31]。洈水水库秋季腐殖质的贡献随着大量树叶的凋落显著提高。虽然秋季陆生植物对鱼类贡献显著降低,有机碎屑对鱼类的贡献显著升高,但鱼类仍然对外源碳的依赖程度很高[32]。综上所述,无论在蓄水期还是枯水期,外源性碳源在支持洈水水库的食物网方面优于内源性碳,这主要与水库周边密集的森林覆盖有关[10,15],因此我们要持续性提高洈水水库周边的森林覆盖率。
3.2 洈水水库夏季和秋季鱼类的营养级
氮稳定同位素(δ15N)沿着食物链每隔一个营养级会产生3‰—4‰的营养富集,因此我们可计算每种水生生物在食物网中的营养级[33]。洈水水库夏季鲤的营养级最低(2.31),花䱻的营养级最高(3.46),这与李峥等[34]在南湾水库的研究结果一致。此前,洈水水库受网箱养殖等人为活动的影响较大,水体呈现富营养化的趋势。水体中N、P含量较高引起底栖动物的δ15N比值偏高,因此,以底栖动物为食的花䱻的δ15N最高(11.67‰)。洈水水库秋季鲤的营养级最低(2.91),鳜营养级最高(3.77),相较于夏季鲤(2.31)和鳜(3.20)的营养级均得到升高。全球范围内湖泊生态系统的食物链长度在4.0左右[36—38],水库的食物链长度一般短于湖泊。上海青草沙水库库容为5.53×108m3,与洈水水库库容相当,胡忠军[14]计算出其食物网长度为3.60,本研究中夏季和秋季的食物链长度与该研究结果相近。有研究显示,食物链的长度和湖泊的面积有关,面积较大的湖泊一般食物链长度较大[39]。Vander Zanden等[38]和Post等[18]都报道了湖泊的生态系统面积的大小与食物链长度的正相关关系,他们所研究的湖泊都分布在很窄的一个经纬度范围内。这表明淡水湖泊生态系统的食物链长度可能还受气候、温度、日照、降水和地理区位等非生物因素的影响。同时也表明了在其他非生物环境因素一致的条件下,淡水湖泊食物链长度主要受湖泊面积的大小影响。例如国内第一大水库——三峡水库(和洈水水库均分布在一个很窄的经纬度范围内)库区面积为1084 km2,洈水水库库区面积仅占其3.15%,和周正等[13]计算出三峡水库的食物链长度为4.12。在两个季节均有采集到的9种鱼类中,有7种鱼类在秋季的营养级高于夏季,表明秋季鱼类的营养级普遍升高。秋季水库水位下降,库区面积缩小使得鱼类活动范围减小,捕食强度就会随之增大,致使肉食性鱼类的比例增加,进而引起秋季鱼类的营养级普遍升高[24]。在枯水季节,由于湖泊面积缩小,食物来源缺乏,鱼类的活动范围受到限制,此时种群密度增大,捕食等相互作用的强度加大,导致鱼类的肉食性增加[8]。而在洪水季节,鱼类可利用的食物源丰富,陆生碳源及沿岸带植物对鱼类的贡献较大[9,10]。同时,湖面面积的增大使得鱼类更加分散,捕食强度相对降低。
3.3 鱼类群落营养结构变化特征
研究鱼类群落营养结构不仅能揭示各生物物种在能量流动过程中较为稳定的摄食与营养关系[8],亦可为渔业资源的管理及可持续利用提供依据[9,10]。为了研究鱼类群落结构的变化趋势,Layman等[19]提出了鱼类群落的同位素量化指标。其中CR、NR、TA和CD用来表示群落营养结构的多样性,MNND、SDNND表示群落冗余度的大小。本研究中夏季鱼类的CR、NR、TA、CD、MNND与SDNND值均高于秋季(表2),表明洈水水库夏季鱼类的初级食物源比秋季的鱼类更加丰富,食物链更长,营养层次更多,核心生态位宽幅更宽,鱼类食物网营养级多样性的总程度较高,营养生态位均匀度较高。夏季CR值更高主要是由于洈水水库在夏季蓄水过程中产生的高水位淹没了大量的陆生植物,同时大量陆生的外源性物质进入水体,为鱼类提供了丰富的饵料资源,而多样化的食物资源为水体食物网营养生态位的增大提供了物质基础[40]。夏季NR值较秋季更高表明群落中有更多的营养层次[19],秋季库区水位下降鱼类食物资源竞争加剧,导致鱼类捕食作用加强[24]。TA值夏季高于秋季(图7和表2),说明夏季群落生态位空间更大,鱼类群落竞争关系得到缓和,这也与夏季饵料资源较为丰富的结果相吻合(表2)。为了减少TA值造成的误差,使用校正后的贝叶斯标准椭圆面积(SEAc)计算生态位空间,计算结果与TA值相似,表明洈水水库鱼类群落对饵料资源的利用水平存在季节差异(表2)。MNND和SDNND值反映了食物网营养冗余度的高低。当食物网中具有相似生态营养级的物种占多数时,NND值则较低,而此时食物网营养冗余程度较高,群落中不同种类之间的营养生态位比较均匀,宽幅较小。与之类似,较低的SDNND值意味着营养级分布更均匀,营养冗余程度较高[19]。从夏季到秋季其营养冗余程度快速降低,表明该生态系统抵抗外界干扰的能力在加强,稳定性增加。CD值代表食物网营养级多样性平均程度,CD值夏季高于秋季,表明洈水水库秋季鱼类的营养级多样性平均程度较低,因此可以通过合理增殖放流鱼类和土著鱼类产卵场营造等方式来提高和保护洈水水库鱼类多样性。
3.4 基于食物网营养结构分析的洈水水库渔业结构调整
夏季水库蓄水,大量陆生植物被淹没,使得植食性和杂食性鱼类在夏季具有广泛的食物来源[22]。秋季水库排水,杂食性、植食性鱼类在夏季和秋季的CR值分别为6.64与4.39、6.26与2.75,表明植食性鱼类和杂食性鱼类在秋季食物资源相对匮乏。同时,较低的水温使得这两种食性鱼类的摄食强度显著降低。因此,建议植食性和杂食性鱼类的鱼苗(草鱼、团头鲂)投放时间在早春(2—3月),草鱼和团头鲂的投放规格应为50尾/kg[41]。随着水温逐步升高,鱼苗的摄食强度增大,水库丰富的饵料有利于鱼苗快速生长[42]。此外,夏季较高的水位增加了库区水域面积,能够有效地规避肉食性鱼类对鱼苗的捕食作用。碎屑食性鱼类在夏季和秋季的CR值分别为3.33和4.83,表明秋季碎屑食性鱼类具有丰富的食物来源。秋季水库周边森林形成的大量落叶进入水体,在微生物的作用下分解为有机碎屑,有利于碎屑食性鱼类在秋季获取丰富的食物资源。因而,建议碎屑食性鱼类鱼苗的投放时间放在秋季更加合适,但由于秋季水位下降,库区面积减少使得捕食强度增大[43],我们应该增加投放鱼苗的大小规格来提升其存活率。肉食性鱼类的CR值在夏季和秋季分别为6.97和6.36,其摄食活动受季节影响较小,但其作为顶级捕食者对维持水体生态系统的稳定具有重要作用[13]。一方面,鳜、鲌等肉食性鱼类通过摄食䱗等小型鱼类提高水库主养鱼类的经济效益[44];另一方面,利用鳜、鲌等肉食性鱼类控制过多的小型鱼类(䱗),让鲢、鳙等滤食性鱼类获得更+多食物资源[45],充分发挥“以渔抑藻、以渔净水”的效果。
猜你喜欢
环境工程技术学报(2022年3期)2022-06-05 07:20:20
阅读(快乐英语高年级)(2022年3期)2022-03-30 21:36:18
当代水产(2021年10期)2022-01-12 06:21:02
橡塑技术与装备(2021年16期)2021-08-27 11:49:12
昆钢科技(2021年6期)2021-03-09 06:10:20
上海包装(2019年2期)2019-05-20 09:10:56
伴侣(2018年2期)2018-02-28 20:38:24
录井工程(2017年4期)2017-03-16 06:10:28
广东石油化工学院学报(2016年3期)2016-05-17 05:16:24
电源技术(2016年9期)2016-02-27 09:05:25
