
千岛湖蒙古鲌种群生物学特征及资源量评估
2024-02-15 18:14:20马玉兴梁智策廖传松盘家永邵建强何光喜刘家寿郭传波
水生生物学报 2024年1期
文 峰 马玉兴 梁智策, 廖传松盘家永邵建强何光喜刘家寿,郭传波,
(1.大连海洋大学,大连 116023;2.中国科学院水生生物研究所淡水生态与生物技术国家重点实验室,武汉 430072;3.中国科学院大学,北京 100039;4.杭州千岛湖发展集团有限公司,淳安 311700)
千岛湖是典型的截流蓄水形成的分枝山谷型水库,具有发电、防洪和航运等功能,也是杭州市的重要饮用水水源地[1]。千岛湖现阶段主要通过人工增殖鲢、鳙等滤食性鱼类发展大水面生态渔业,对食物网中其他食性鱼类的研究和关注较少。鱼食性鱼类(Piscivorous fish)常处于食物链的顶端,是水生生态系统的顶级捕食者[2],在水域生态系统功能中具有重要作用,如通过营养级联调控食物网和生态系统[3]、参与营养物质循环[4,5]、调控物种入侵[6]等。鱼食性鱼类也是经典生物操纵的重要对象,即通过放养鱼食性鱼类控制浮游动物食性小型鱼类生物量,间接维持浮游动物生物量,增强对浮游植物的控制作用,从而达到控藻和净水目的[7]。因此,鱼食性鱼类也是千岛湖食物网的重要组成部分。
现阶段,千岛湖的鱼食性鱼类资源主要包括蒙古鲌(Culter mongolicusBasilewsky 1855)、翘嘴鲌(Culter alburnusBasilewsky 1855)、大眼鳜(Siniperca kneriiGarman 1912)和斑鳜(Siniperca scherzeriSteindachner 1892)等[8,9],是当地渔民捕捞和垂钓的重要渔获物。千岛湖鱼食性鱼类资源的管理和开发利用由淳安县农业综合执法部门统一管理,其推行了包括设置禁渔区和禁渔期、限制捕捞(游钓)的渔具渔法、发放规定数量的捕捞(游钓)许可证在内的一系列渔业管理措施。当地渔民和游钓者获得许可后方能在非禁渔期(每年3—6月为禁渔期)和非禁渔区(中心和西北湖区部分核心水域常年禁渔)使用规定的渔具渔法开展相应的渔业捕捞及垂钓。但近年来,包括鱼食性鱼类在内的土著鱼类出现了资源量明显下滑的趋势[10]。
蒙古鲌隶属于鲤形目(Cypriniformes)鲤科(Cyprinidae)鲌属(Culter),是千岛湖生态系统中优势的鱼食性鱼类,也是千岛湖周边重要的名贵经济鱼类和休闲游钓的主要对象,俗称“红珠”,市场价格达200元/kg。近年来,千岛湖蒙古鲌资源也出现了较大幅度的下滑,但有关其种群生物学特征和资源量却鲜有研究和报道。
因此,本文系统研究了千岛湖蒙古鲌的年龄、生长、繁殖和死亡等种群生物学特征,并对其种群资源利用率和资源量进行了估算,为进一步保护、恢复和科学利用千岛湖鱼食性鱼类资源,优化和完善鱼类群落结构提供理论依据。
1 材料与方法
1.1 样品采集与处理
2021年3月至2022年9月,逐月在千岛湖采集蒙古鲌新鲜样本共431尾(表1)。在新鲜状态下,测量蒙古鲌体长(Standard length,SL,1 mm)和体重(Body weight,BW,0.01 g)。随机选取部分样本进行生物学解剖,鉴定性别(雌、雄和未分辨),并称量性腺重(Gonad weight,GW,0.01 g),根据其性腺成熟度进行分期(Ⅰ—Ⅵ)。对于Ⅳ期卵巢,随机称取约0.5 g卵粒,用5%的甲醛溶液固定保存。随后移除所有内部器官后称量空壳重(Somatic weight,SW,0.01 g)。采集背鳍起点下方、侧线鳞以上的5—10枚鳞片作为年龄鉴定材料,鳞片经4%NaOH溶液浸泡12h,洗净后置于解剖镜(OLYMPUS)下观察、鉴定年龄,并测量鳞径(Scale radius,SR,0.01 mm)。计数所取卵巢样本的卵粒数,并采用重量估算法计算成熟雌性个体的繁殖力,并使用LightTools软件测量卵径(Oocyte diameter,OD,0.01 μm)。鱼类年龄的鉴定及性腺成熟度分期方法等参照殷名称[11]进行。

表1 千岛湖蒙古鲌样本数量及信息Tab.1 Quantity and information ofC.mongolicusin Qiandaohu Lake
1.2 数据分析
不同性别蒙古鲌个体平均体长和体重的差异、体长与体重关系(BW=aSLb)差异分别采用Wilcoxon秩和检验和协方差分析(ANCOVA)进行比较。蒙古鲌种群体长体重关系的b值和理论值3之间的差异使用t检验进行判断从而确定鱼类的生长型。不同性别蒙古鲌体长与鳞径关系的差异同样采用协方差分析(ANCOVA)进行检验。以25 mm为组距对蒙古鲌体长进行分组[12],并使用体长频率法(Electronic Length Frequency Analysis,ELEFAN)对其生长参数L∞和k值进行估算,理论生长起点年龄t0根据Pauly[13]的经验公式获得:
随后选择Von Bertalanffy (VBGF)生长方程拟合并描述千岛湖蒙古鲌体长和体重的生长规律:
式中,Lt和Wt分别表示t龄鱼的体长和体重,L∞和W∞分别表示渐近体长和渐近体重,k表示生长曲线的生长系数,t0表示理论上Lt=0时的年龄。
使用卡方检验判断蒙古鲌雌雄性比与理论值1∶1的差异。利用性体指数(Gonado somatic index,GSI)来判断鱼类繁殖时间,计算公式为:
依据卵粒计数结果和所取卵粒总重计算每克重卵粒数,并结合性腺重、空壳重,分别推算千岛湖蒙古鲌的绝对繁殖力(Absolute fecundity,AF)与相对繁殖力(Relative fecundity,RF):
式中,n表示统计的卵粒数,W0为所取卵粒样本重量。
本研究采用体长变换渔获曲线法(Length-converted catch curves)估算总死亡系数(Z),自然死亡系数(M)则根据Pauly等[14]提出的经验公式计算:
式中,T为千岛湖年平均水温,为21.45℃。捕捞死亡系数(F)为总死亡系数和自然死亡系数之差。鱼类种群开发率(E)则通过捕捞死亡率和总死亡率(E=F/Z)进行推算。运用Pauly和Soriano的动态综合模型来预测相对单位补充量渔获量(Y′/R)和相对单位补充量生物量(B′/R),并通过FiSAT Ⅱ软件中的刀刃式选择假设模型(Knife-edge model)构建Y′/R和B′/R曲线[14],公式如下:
式中,Lc表示开捕体长,使用千岛湖采集的蒙古鲌样本中规格最小的20尾平均值。种群开发率(E)一般有常见几种水平:Emax为获得最大渔获量时的开发率;E0.1为Y′/R边际增长减少10%时的开发率;E0.5为资源量下降到原始水平50%时的开发率[15]。
本研究采用体长股分析法(Length based cohort analysis,LCA) 对蒙古鲌种群资源量进行估算[16]。利用千岛湖蒙古鲌的年渔获量、渐近体长(L∞)、生长系数(k)、自然死亡系数(M)、经迭代稳定后最大体长组的捕捞死亡系数(Ft)及体长与体重关系式中的常数(a)和生长指数(b)等,经模型计算得出各体长组的捕捞死亡系数和资源量[12]。
本研究中采用FiSAT Ⅱ和R 4.2.2分析数据和作图,数据采用平均值±标准误(mean±SE)表示,以P<0.05为显著性水平。
2 结果
2.1 蒙古鲌种群体长体重与年龄结构
千岛湖蒙古鲌的体长为135—645 mm (N=431),平均体长为(265.99±2.42) mm,优势体长集中在200—300 mm,占总数的81.90%。体长在200 mm以下和400 mm以上个体比例较低,仅占5.10%(图1a)。在能分辨性别的个体中,雌性个体体长为182—645 mm(N=209),平均体长为(274.99±4.03) mm,雌性个体的优势体长为250—300 mm,占比为56.46%;雄性个体为180—392 mm (N=202),平均体长为(259.83±2.71) mm,雄性个体的优势体长为200—300 mm,占总体的79.70%。雌性个体平均体长显著大于雄性个体(Wilcoxon test,W=24461,P<0.01)。
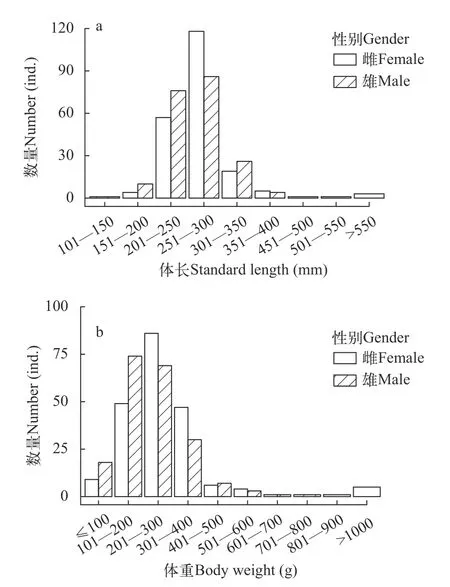
图1 千岛湖不同性别蒙古鲌的体长体重分布Fig.1 Standard length and body weight distributions ofC.mongolicuswith different genders in Qiandaohu Lake
千岛湖蒙古鲌的体重为31.20—4690.23 g,平均体重为(270.77±15.68) g,优势体重集中在100—400 g,占总数的85.61% (图1b)。其中,雌性个体体重为55.21—4690.23 g,平均体重为(319.80±31.04) g,雌性个体的优势体重为200—300 g,占比为41.15%;雄性个体体重为61.85—750.01 g,平均体重为(230.25±7.68) g,雄性个体的优势体重为100—300 g,占总体的70.30%。雌性个体平均体重显著大于雄性个体(Wilcoxon test,W=25557,P<0.001)。
本研究共鉴定242尾蒙古鲌样本的年龄。结果显示: 蒙古鲌种群的年龄组成为1—8龄,以2—3龄为优势年龄组,占总数的85.95%,高龄(>4)和低龄(<2)个体数量较少,仅占总数的4.96%(图2)。总体上看,种群年龄结构较为完善,但趋于低龄化。鳞径与体长关系为: SL=91.7SR+57.5(R2=0.864,P<0.001),鳞径与体长的关系在雌雄个体间差异不显著(ANCOVA,F=2.155,P>0.05)。按照鳞径与体长的关系进行体长退算(表2),各年龄段实测平均体长与退算体长间无显著性差异(t-test,P>0.05)。

图2 千岛湖蒙古鲌的年龄结构Fig.2 Age structures ofC.mongolicusin Qiandaohu Lake

表2 千岛湖蒙古鲌鳞径的体长退算Tab.2 Back calculation of the standard length ofC.mongolicusin Qiandaohu Lake
2.2 蒙古鲌种群生长特征
蒙古鲌的雌、雄性个体体长与体重关系无显著差异(ANCOVA,F=1.709,P>0.05),因此可将雌、雄群体合并使用同一方程式来表示其体长与体重关系,其表达式为: BW=2.67×10-6SL3.27(R2=0.981,P<0.001,N=431)。式中,b值(3.27)与理论值3存在显著差异(t-test,P<0.001),说明蒙古鲌的生长类型属于正异速生长类型。
蒙古鲌体长和体重Von Bertalanffy生长方程分别为Lt=656.25·[1-e-0.33(t+0.35826)]和Wt=4348.38·[1-e-0.33(t+0.35826)]3.27。对上述Von Bertalanffy生长方程分别求一阶和二阶微分,得到千岛湖蒙古鲌体长和体重生长速度及生长加速度的方程如下:
体长生长速度: dL/dt=216.56·e-0.33(t+0.35826)
体长生长加速度: d2L/dt2=-71.47·e-0.33(t+0.35826)
体重生长速度: dW/dt=4692.34·e-0.33(t+0.35826)·[1-e-0.33(t+0.35826)]2.27
体重生长加速度: d2W/dt2=1548.47·e-0.33(t+0.35826)·[1-e-0.33(t+0.35826)]1.27·[3.27e-0.33(t+0.35826)-1]
蒙古鲌体长和体重的生长速度曲线、生长加速度曲线显示,蒙古鲌体长生长速度随着年龄的增长而递减,在高年龄时趋近于0,体长生长加速度却持续增加,最终也趋近于0(图3a)。蒙古鲌体重生长速度和加速度曲线均具有明显的拐点,拐点对应年龄为3.23龄,对应体长和体重分别455.43 mm和为1316.88 g。在拐点前体重生长速度随着年龄增加逐渐上升,至拐点处达到最大值。拐点之后加速度为负值,对应体重生长速度呈递减变化。在5—6龄处体重生长加速度降至最低,随后缓慢上升并趋近于0(图3b)。
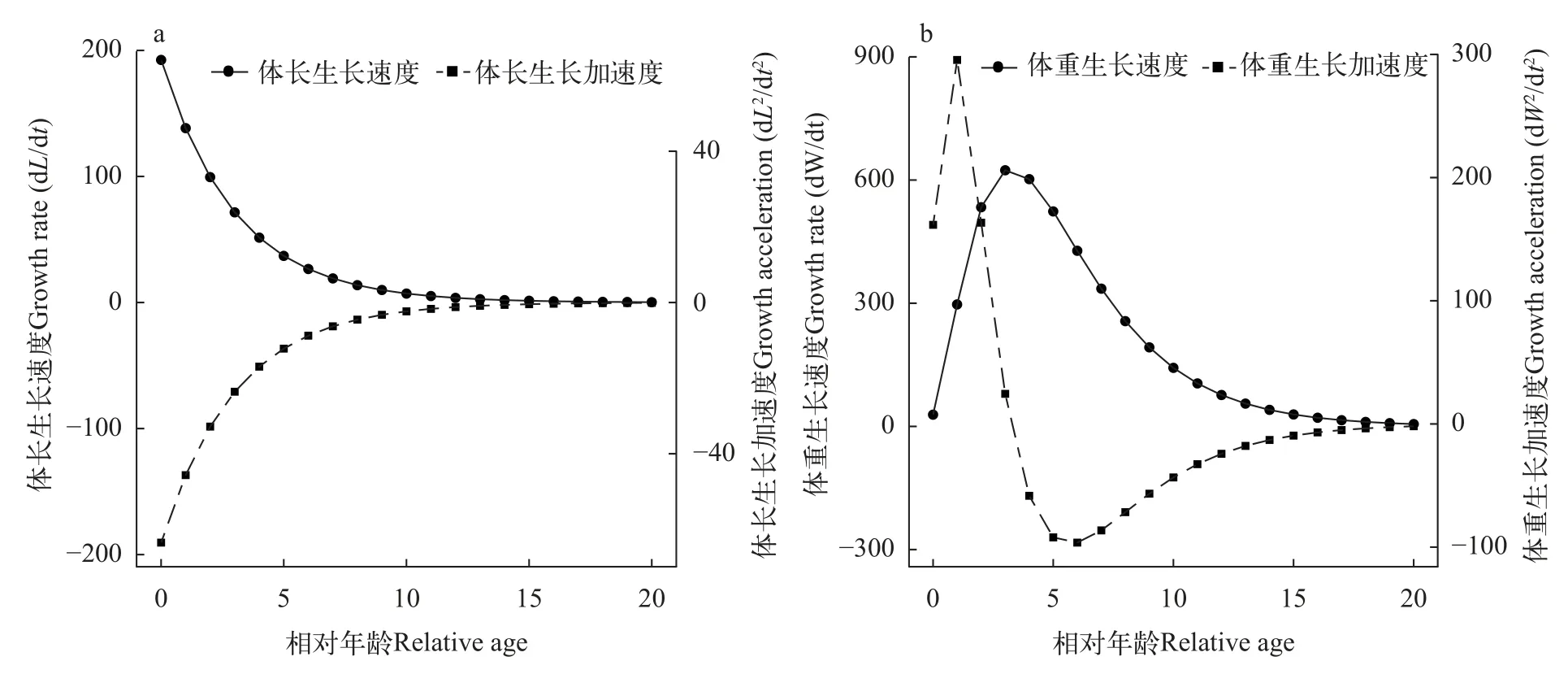
图3 千岛湖蒙古鲌体长体重生长速度/加速度曲线Fig.3 Growth rate,growth acceleration rate of standard length and body weight ofC.mongolicusin Qiandaohu Lake
2.3 蒙古鲌种群繁殖特征
蒙古鲌雌雄性比为1.03(雌209尾,雄202尾),与理论值1∶1无显著差异(χ2=0.044,P>0.05)。蒙古鲌雌性个体的性体指数(GSI)在4—5月开始逐步上升,于7月到达最高值,随后在8、9月迅速回落,9月后至翌年3月GSI持续保持低水平。雄性个体的GSI变化趋势与雌性个体相似(图4a)。蒙古鲌性腺发育月度变化显示,4月开始已发现部分个体性腺发育至Ⅳ期,5月时Ⅳ期个体占比最高(44.44%),6月和7月超过半数个体性腺已发育至Ⅴ期(56.10%;57.84%),在8月和9月Ⅵ期性腺占比逐步升至最高(34.21%;58.82%),至12月几乎全部退化到Ⅱ期(图4b)。因此,千岛湖蒙古鲌的繁殖期为4—8月,繁殖高峰则为6—7月。

图4 千岛湖蒙古鲌性体指数和性腺发育时期月变化Fig.4 Gonado-somatic index and gonadal development stage variation by month ofC.mongolicusin Qiandaohu Lake
采集并测量了86尾雌鱼成熟卵粒的卵径,卵径范围为433.00—2145.00 μm,平均卵径为(1085.76±6.11) μm (n=1670),主要集中在900.00—1300.00 μm(66.29%)。卵径分布呈单峰状(图5),结合GSI急剧的变化趋势,初步推测蒙古鲌为单批次产卵类型。蒙古鲌个体绝对繁殖力为6054—1078974粒/尾,平均绝对繁殖力为(58681±13186)粒/尾,个体体重相对繁殖力为23.92—535.59粒/g,平均相对繁殖力为(216.87±9.60)粒/g。
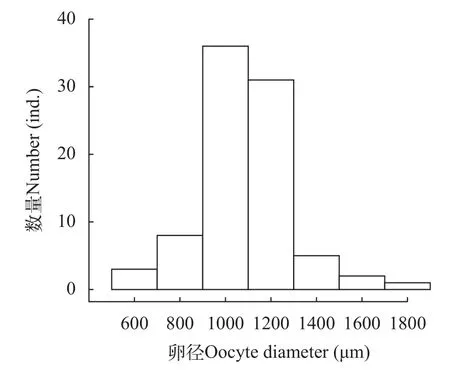
图5 千岛湖蒙古鲌卵径分布Fig.5 Oocyte diameter distributions ofC.mongolicusin Qiandaohu Lake
2.4 蒙古鲌种群死亡特征及资源量评估
运用体长变换渔获物曲线法估算出千岛湖蒙古鲌的总死亡系数(Z)为2.26。将前述千岛湖蒙古鲌的渐近体长(L∞)、生长系数(k)及调查年度千岛湖的平均水温(T)代入经验公式,计算出其自然死亡系数(M)为0.32,进一步计算得到当前捕捞状态下的捕捞死亡系数(F)为1.94,并由E=F/Z得出种群开发率(E)为0.86。
选取20尾最小规格样本的体长平均值作为千岛湖蒙古鲌的开捕体长,即Lc=188.85 mm。通过刀刃式假设模型建立了相对单位补充渔获量、生物量与种群开发率的关系,以二维分析反映千岛湖蒙古鲌的开发程度(图6),估算出蒙古鲌种群的E0.1为0.40、E0.5为0.32、Emax为0.52。对比发现,当前蒙古鲌种群开发率E=0.86>Emax,说明千岛湖蒙古鲌处于过度捕捞的状态。采用估算最大体长组的资源尾数为起点计算千岛湖蒙古鲌的现存资源量,最大体长组的捕捞死亡系数(Ft)取0.5为初始值,经重复迭代后将稳定的数值代入VPA模块,估算出千岛湖蒙古鲌的年资源数量约347887尾,通过已有的体长-体重关系换算成年资源生物量则为259.4吨(表3)。
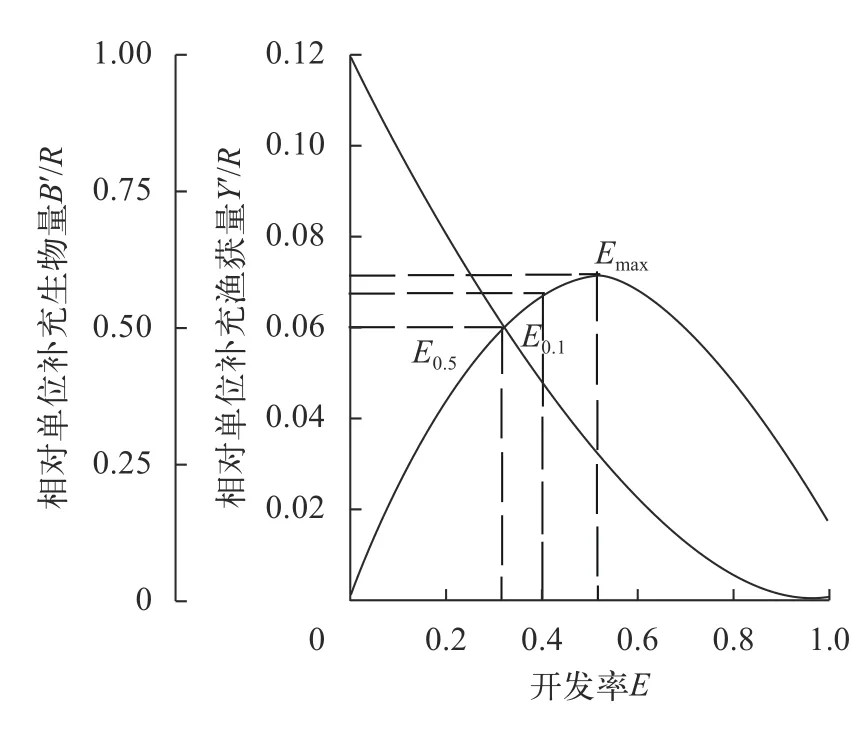
图6 千岛湖蒙古鲌相对单位补充渔获量和生物量的二维分析Fig.6 The relative yield per recruit (Y′/R) and the relative biomass per recruit (B′/R) curves ofC.mongolicusin Qiandaohu Lake
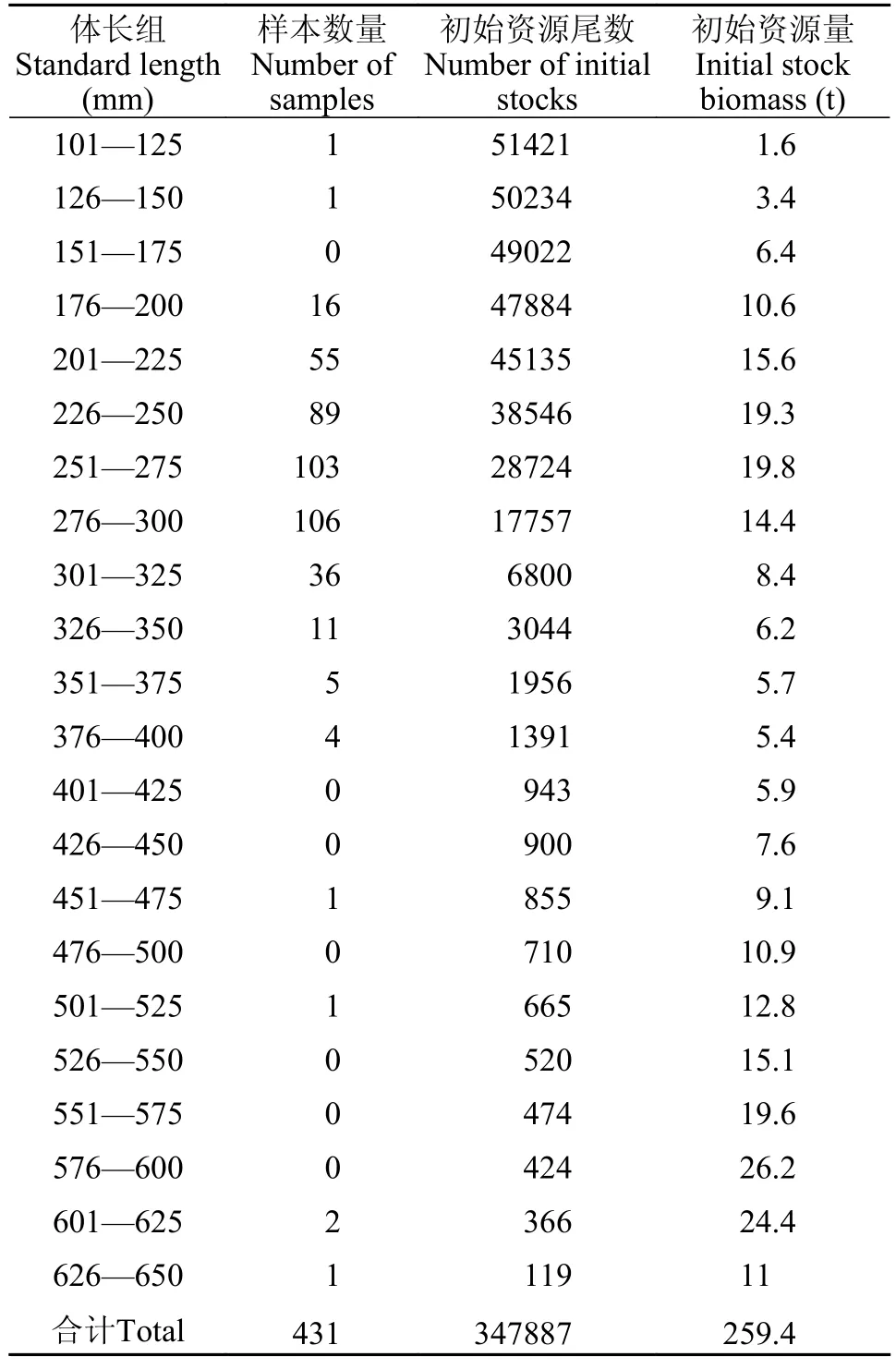
表3 千岛湖蒙古鲌初始资源量估算Tab.3 Initial stock biomass ofC.mongolicusin Qiandaohu Lake
3 讨论
3.1 千岛湖蒙古鲌的年龄与生长特征
本研究发现千岛湖的蒙古鲌种群共8个年龄组,其年龄结构比其他水域的蒙古鲌种群更为复杂,具有更完整的种群结构,如鄱阳湖(1—5龄)[17]、常德西湖(1—6龄)[18]、镜泊湖(1—6龄)[19]、老江河水库(1—4龄)[20]及泼河水库(1—3龄)[21]。其中低龄鱼(2—3龄)的数量占绝对优势(85.95%),1龄个体的数量较少,这可能由于1龄个体规格较小尚不能完全被渔业活动所捕获;高龄个体数量也较少,说明千岛湖的蒙古鲌种群出现低龄化现象。
鱼类生长参数可以有效反映鱼类的生命周期和生长速度[22],是研究资源群体数量变动和合理利用的基础[12],有利于渔业管理和鱼类资源保护。现有的研究认为生长系数k值为0.05—0.10的鱼类是生长缓慢的鱼类,0.10—0.20为均匀生长种,0.20—0.50为快速生长种[23],因此,千岛湖蒙古鲌种群属快速生长种(k=0.33)。与其他水域的蒙古鲌种群相比,千岛湖蒙古鲌的生长系数(k)相较鄱阳湖[17]、常德西湖[18]、老江河水库[20]、泼河水库[21]及武汉东湖[24]的蒙古鲌种群更高,也高于同为鲌属的翘嘴鲌[25—27]、达氏鲌[28]、拟尖头鲌[29]等。造成这种现象的原因可能与其水域生态环境、水体饵料生物的丰度及种质资源差异有关[30]。
本研究由Von Bertalanffy生长方程拟合了千岛湖蒙古鲌的生长规律,其渐近体长L∞=656.25 mm、渐近体重W∞=4348.38 g,均大于鄱阳湖[17]、老江河水库[20]及泼河水库[21]的蒙古鲌种群,且与采集样本的最大观测数据已较为接近,表明千岛湖蒙古鲌生长潜能得到了充分发挥,已接近其生长极限。综合来看,千岛湖水域各项水体指标较好,水体透明度高,具有良好的水域生态环境,有利于鱼类种群生长[31]。鱼类群落采样调查结果显示: 千岛湖的飘鱼(Pseudolaubuca sinensis)、似鱎(Toxabramis swinhonis)和䱗(Hemiculter leucisculus)等小型鱼类资源量庞大,其数量在渔获物中比例高达53.87%,是千岛湖鱼类群落中的主要优势种(数据未发表),为蒙古鲌等鱼食性鱼类提供了充足的饵料资源。通过渔业码头渔获物调查也发现,渔获物组成中小型鱼类比例较高、鱼食性鱼类资源较少,侧面反映了千岛湖潜在饵料鱼类资源丰富而鱼食性鱼类间捕食竞争较弱。这些因素共同保证了千岛湖蒙古鲌种群具有较快生长速度和较高的生长潜力。
关于鱼类的生长速度,可划分为3个主要的阶段,即性成熟前的生长迅速阶段、性成熟后的生长稳定阶段和生长衰老阶段[24,32,33]。有些鱼类的生长迅速阶段可维持到性成熟后的1—2年,以保证其较高的繁殖力。生长指标和生长拐点理论上能较客观地反映这一点[34]。拐点年龄理论上是鱼类生长速度转折点,在生产实践中也常依托于此确定起捕年龄和规格[22]。蒙古鲌2龄即可达到性成熟[35],本研究结果显示千岛湖蒙古鲌的体重生长拐点年龄为3.23龄,对应体长455.43 mm,正处于性成熟年龄后1—2年内,这表明千岛湖蒙古鲌在性成熟后仍能保持较快生长速度,为提高其绝对繁殖力和在复杂环境中维持物种延续提供了有利条件。但我们发现千岛湖目前针对种群快速生长时期(2—3龄)的个体并未有渔业捕捞和休闲垂钓方面的限制,反而该年龄组个体在渔获物中占比较高。因此,建议将开捕体长和垂钓限制体长设置为拐点年龄对应体长,此举可保障千岛湖蒙古鲌种群的快速生长,利于种群资源的可持续开发。
3.2 千岛湖蒙古鲌种群的繁殖策略
繁殖是鱼类生活史的重要环节,为鱼类种群的繁衍和发展提供保障。繁殖策略是鱼类长期适应自然环境的结果,鱼类总是不断地寻求最优的两性比例、繁殖时间和场所以及繁殖方式,最大限度地提高后代的成活率[36,37]。鱼类种群的雌雄性比通常接近1∶1,但产卵鱼群的性比多数为雄性多于雌性,以提高卵子的受精率[38]。在本研究中,蒙古鲌的雌雄性比为1.03,与理论值1∶1没有显著的差异。一方面表明目前的捕捞方式未导致蒙古鲌种群性比失衡;另一方面可能是因为在性成熟后小型个体中雄性占比较大(55.10%),而大型个体中雌性占比较大(63.11%),与鱼类性比在不同体长组可能不同的观点相符合[11]。
前期的研究发现贵州地区的蒙古鲌产卵时间在5—7月,以6月为产卵高峰期[32]。而千岛湖蒙古鲌明显具有更长的繁殖时期,结合性体指数(GSI)的周年变化趋势推测千岛湖蒙古鲌种群自4月开始进入繁殖期,繁殖盛期集中在6—7月,8—9月产卵结束。有研究发现,鱼类倾向于延长繁殖时期,从而使其后代获得优渥的发育环境条件[11]。雌性个体的GSI较雄性个体更高,体现了亲鱼资源物质主要分配给卵巢,较大的生殖器官一方面能储存更多的营养物质,可增加产卵数量并提高仔鱼的存活率;另一方面有利于延长繁殖周期,能够在更多的时间内参与繁殖,为种群延续和鱼类资源可持续利用提供保障。千岛湖自4月起水体中开始出现温跃层和斜温层,可认为此时水温明显升高。表层水温和20 m深处水温分别在7和8月达到最高[31]。千岛湖蒙古鲌的GSI月变化趋势与水温变化趋势具有一致性,表明水温的变化可能是驱动千岛湖蒙古鲌繁殖的关键因素。
鱼类的繁殖力和卵径是繁殖策略的重要表征,两者通常存在权衡,以提高对环境的适应力[11]。千岛湖蒙古鲌的平均个体绝对繁殖力为(58681±13186)粒/尾,平均相对繁殖力为(216.87±9.60)粒/g,该数值均小于常德西湖[39]和丹江口水库[40]蒙古鲌的繁殖力,可能因为千岛湖采集的卵粒样本多数来自于相对年龄较小的个体。一般而言,鱼类的繁殖力与年龄密切相关,前期繁殖力随年龄增大,到了高龄后繁殖力增加有所减缓甚至下降[11]。这也从侧面说明目前千岛湖的捕捞模式易导致蒙古鲌种群的低龄化,不利于蒙古鲌资源的可持续利用。
鱼类产卵类型一般通过判断鱼类卵径分布类型为单峰或多峰,辅以GSI变化趋势来确定。曹克驹等[40]发现丹江口水库蒙古鲌的卵径出现双峰,认为其为多批次产卵类型。现有研究发现长湖[38]和长江[41]的达氏鲌及徐家河水库[34]的翘嘴鲌可能为分批产卵类型,而太湖的翘嘴鲌则被认为是单批次产卵[32]。但本研究中蒙古鲌卵径分布呈单峰状,主要集中在0.9—13.0 mm。此外,千岛湖蒙古鲌的卵径自5月至7月逐渐增大,结合雌性个体的GSI在繁殖结束时期表现出急剧下降的趋势,推测千岛湖蒙古鲌群体可能为单批次产卵类型。综合来看,千岛湖蒙古鲌繁殖季节相对较长,使其获得更多的繁殖机会,具有较高的繁殖力,整体表现周期主义的繁殖策略[42],从而使蒙古鲌在千岛湖与其他种类的竞争处于优势地位,最终成为千岛湖优势的鱼食性鱼类。
3.3 千岛湖蒙古鲌种群资源现状
影响渔业资源量及其可持续性的因素有很多,例如鱼类种群规模大小、开捕年龄及捕捞强度等[43]。在我国内陆捕捞渔业中,捕捞强度过大已成为渔业生产可持续发展面临的重要威胁,也给渔政监管工作带来很大的压力[44]。本研究估算出千岛湖蒙古鲌总死亡系数、自然死亡系数、捕捞死亡系数及开发率分别为2.26、0.32、1.94和0.86,初始资源尾数为34.79万尾,初始资源总量为259.4 吨。有研究提出总死亡系数(Z)与生长系数(k)的比值Z/k≤3时,表明自然死亡为种群死亡的主要原因;当Z/k>3时,则表明捕捞死亡为其主要原因[12],本研究中千岛湖蒙古鲌的Z/k=6.85,说明千岛湖蒙古鲌种群的死亡原因主要由人为捕捞导致。有研究提出鱼类最适利用率为0.5[45],而当前千岛湖蒙古鲌的种群开发率(E)为0.86,已超过最适利用率。在Y′/R与E关系曲线中,Emax的左侧区域即开发率E<Emax,代表当前渔业资源能可持续发展[46],千岛湖蒙古鲌的开发率明显位于Emax的右侧区域。从生物量资源的维持角度来看,当开发率达到E0.5时约50%的生物量资源可以被保持,开发率增加至Emax时仅有约25%的生物量资源能得到保持,若在当前开发率下,仅能保持现有蒙古鲌生物量资源的5%不被破坏。本研究发现蒙古鲌样本体长集中在200—300 mm,该体长范围远低于拐点年龄对应体长(455.43 mm),上述数据均说明渔业捕捞对于蒙古鲌尤其是低龄个体的捕捞强度过大,千岛湖蒙古鲌的资源正处于过度开发状态。据不完全统计,千岛湖现有可开展土著鱼类捕捞渔业的渔民301户,渔船378艘,垂钓证接近5000张。鉴于此,应当进一步加强蒙古鲌种群资源的管理。
4 结论
伴随着经济快速发展,包括千岛湖在内的大多数水域生态系统中顶级捕食者普遍被过量捕捞。此外水体污染、生境破坏等人为扰动也是造成顶级捕食者资源量衰退的主要原因[47,48]。越来越多的证据表明,顶级捕食者的资源下滑会对生态系统功能产生深远的影响[3,49]。多个研究结果表明,鱼食性鱼类的资源衰退后,水域生态系统中小型鱼类会因捕食压力减弱而出现暴发式增长,最终造成鱼类群落趋于小型化[50,51]。蒙古鲌作为千岛湖鱼食性鱼类的优势种之一[52],若将其鱼类资源量保持在合理的范围内,一方面有利于控制湖泊和水库中常见的小型定居性鱼类(如千岛湖中常见的飘鱼、似鱎和䱗等),将经济价值低的小型鱼类资源转化为经济价值较高的水产品,对于渔业经济效益也具有重大意义[53];另一方面,也可以通过捕食和竞争控制外来入侵的太阳鱼种群在千岛湖大量繁殖和扩张。因此,为保障千岛湖渔业的可持续发展、维护水域生态系统的健康,亟须对千岛湖蒙古鲌资源开展科学的保护、恢复及合理利用。
千岛湖蒙古鲌种群已处于过度开发阶段,如不及时开展科学、有效的管理措施,其资源量将会进一步衰退,渔业价值持续降低,也不利于千岛湖生态系统的健康和稳定发展。有鉴于此,基于本研究的研究结果,针对千岛湖蒙古鲌资源的管理提出以下建议: (1)限制捕捞规格。千岛湖蒙古鲌的拐点年龄为3.23龄,对应体长为455.43 mm,远高于渔获物中蒙古鲌的平均体长(265.99±2.42) mm。建议将千岛湖蒙古鲌的起捕年龄设置为3龄以上,起捕体长大于455.43 mm;(2)调整禁渔时限。千岛湖蒙古鲌的繁殖盛期为7月,但目前千岛湖大部分水域禁渔期为每年3月1日至6月30日,仅有西北和中心湖区部分核心水域常年禁渔。为有效保护繁殖种群以及蒙古鲌资源可持续开发和利用,建议将禁渔期延长至8月末;(3)加强生境营造。蒙古鲌产黏性卵,卵排出后易黏附于杂草或其他物体上,其产卵场主要集中在千岛湖上游入湖口。因此,可在上游入湖口水域加强人工生境的营造,保护和恢复蒙古鲌的产卵生境。
猜你喜欢
人文天下(2022年7期)2022-09-15 11:57:34
天天爱科学(2022年9期)2022-09-15 01:12:28
科普童话·百科探秘(2022年8期)2022-05-30 10:48:04
昆明医科大学学报(2022年1期)2022-02-28 07:46:32
阅读与作文(小学高年级版)(2019年8期)2019-10-16 04:46:50
西域历史语言研究集刊(2018年0期)2018-11-09 01:01:40
草原歌声(2017年1期)2017-04-23 05:08:53
中国自行车(2017年1期)2017-04-16 02:54:08
快乐作文·低年级(2016年2期)2016-02-24 19:12:09
交通建设与管理(2015年15期)2015-03-20 15:19:28
