
不同冻结方式对手抓羊肉微观结构、水分迁移及品质的影响
2023-12-05 08:09毕永昭罗瑞明
中国食品学报 2023年10期
毕永昭,罗瑞明
(宁夏大学食品与葡萄酒学院 银川 750021)
冷冻是肉制品品质保真的常用方式之一,能够较大限度地保持肉制品原有的品质。冷冻肉制品的品质受冻结速率的影响,冰晶的数量、体积和分布与冻结速率有关[1]。慢速冻结时冰晶成核率低,形成少量大冰晶,可能会对肉制品的微观结构造成损害,而快速冻结促进高成核率,使肉制品中形成小冰晶,从而最大限度地减少对质地的损害[2-3]。因此,通过控制冻结速率影响冰晶的形成来提高肉制品的品质非常重要,然而,长时间维持高冻结速率造成的能耗过高问题势必给企业增大成本。共晶点是物料内全部的游离水从液态转换为固态时的温度,位于物料最大冰晶生成带之后,是产品在冻结后达到的一种相对稳定的状态[4-5]。通过高速率冻结使产品在极短时间内迅速降温至共晶点温度,直接进行预包装后运入商业速冻库(-18 ℃),在提高产品品质的同时减少生产周期,可降低企业的生产成本[6-7]。目前,有关冻结速率对手抓羊肉品质特性的影响研究还未见报道。
宁夏手抓羊肉以滩羊肋排为原料加工而成,是中国西北地区极具民族特色的传统煮制类肉制品,因独特的风味与口感,长期以来深受消费者喜爱[8-9]。随着“盐池滩羊”品牌效应的扩大及全产业链的发展,手抓羊肉逐渐走向全国[10]。目前市售的手抓羊肉商品多以热杀菌技术或添加食品防腐剂来延长货架期,而热杀菌导致的肉制品不良品质变化和外源添加引起的消费者担忧仍然严重阻碍着滩羊肉制品加工产业的发展[11-12]。本文以共晶点作为冻结终温,研究不同冻结方式对手抓羊肉品质特性的影响,以期解决维持高冻结速率所导致的成本过高问题,为“无化学”肉制品冷加工过程和民族特色肉制品低温品质保持提供理论参考。
1 材料与方法
1.1 材料与试剂
滩羊肋排由宁夏盐池县大夏牧场食品有限公司提供。随机选取饲养条件一致的6 月龄阉割公羊,经屠宰、放血、去内脏、清洗,在屠宰放血后约2.5 h 沿脊柱垂直于肋排方向取左侧第4 至第10根肋排并切割成单排,以泡沫箱冰袋的运输形式在采后3 h 内运至实验室,4 ℃冷藏过夜,切成10 cm 羊排备用。
四氧化锇、戊二醛,德国徕卡;醋酸双氧铀、柠檬酸铅染液、812 环氧树脂包埋套装,北京中镜科仪技术有限公司。
1.2 仪器与设备
CR-400 型色彩色差计,日本KONICA 公司;LT202E 型电子天平,常熟市天量仪器有限责任公司;TA-XT plus12587 质构仪,英国Stable Micro System 公司;DHG9075A 电热鼓风干燥箱,上海一恒科学仪器有限公司;NMI20 型核磁共振成像仪,上海纽迈电子有限公司;EMTP 组织处理机、EMKMR3 玻璃制刀机、Leica UC7 超薄切片机,德国徕卡仪器(中国)有限公司;JEM-1400FLASH 透射电子显微镜,日本电子株式会社(JEOL);DW-8L398S 超低温保存箱,无锡冠亚恒温制冷技术有限公司;MDF-40V328 医用低温保存箱,安徽中科都菱商用电器股份有限公司;BCD-536WKN 食品专用冰箱,合肥美的电冰箱有限公司;MAL20-B05电磁炉,中山市麦勒电器有限公司;G1224 不锈钢汤锅,广东海龙不锈钢器皿有限公司;DSC 800003061404 型差示扫描量热仪,美国Perkin Elmer 公司;TC-08 热电偶数据记录仪,英国Pico公司。
1.3 方法
1.3.1 样品处理 单次称取(500±10)g 肋排,浸泡30 min,捞出沥干水分放入锅中,倒入2 L 纯净水(肉水比1∶4),除食盐外不加任何调味料,用大火(2 200 W)煮制30 min 至沸腾(宁夏银川的海拔1 500 m,水的沸点为95.6 ℃),撇净浮沫,调至小火(800 W)继续煮制70 min,煮制完成捞出沥干水分,装入240 mm×350 mm×0.12 mm 规格的聚乙烯自封袋中,自然晾凉至室温,立即进行冻结处理[8,13]。对照组为未经冷冻的样品,处理组分别在-18,-40,-80 ℃条件下冻结至共晶点温度,达到冻结终点后立即取出置于4 ℃冰箱冷藏解冻24 h,完成解冻后剔除骨、皮、筋膜、脂肪等其它部分进行试验测定[14]。
1.3.2 共晶点测定 采用差示扫描量热法(Differential scanning calorimetry,DSC)测定物料的共晶点温度[4-5]。试验过程采取降温方式进行,称取2~8 mg 熟制滩羊肉置于差示扫描量热仪的铝坩埚中,并采用氮气保护,以2 ℃/min 的降温速率从25 ℃开始冷却到-70 ℃,根据样品在降温过程中的放热峰峰值判定样品共晶点,结果采取3 次平行的平均值。
1.3.3 冻结过程温度测定及冻结速率的计算 将样品随机分成4 组,采用TC-08 热电偶数据记录仪进行多通道温度测定和记录,热电偶温度探针插入到手抓羊肉的几何中心,记录温度变化[15],每隔1 min 记录1 次,直至冻结温度达到共晶点,平行测定6 次,并绘制冻结温度曲线。
1.3.4 色泽测定 参照Zhang 等[16]的方法对解冻样品的色泽进行测定。测定前校准色差计,每个处理组选取3 个样品,每个样品表面随机选取4 个位置测定其L*(亮度值)、a*(红度值)、b*(黄度值),测量时镜头垂直并紧贴肉面,防止漏光。
1.3.5 解冻损失率测定 参照Zhang 等[17]的方法测定。
1.3.6 水分含量测定 采用《食品安全国家标准食品中水分的测定》(GB 5009.3-2016)中的直接干燥法[18],每个处理组做3 次平行试验。
1.3.7 低场核磁共振测定 根据单启梅[19]的方法并稍作修改,每个处理做3 次平行试验。正式测样前进行仪器校准,玻璃检测管(直径15 mm)中放入油样,通过核磁共振分析应用软件中的FID(Free induction decay)脉冲序列自动寻找90°脉宽P1,并校正中心频率O1。将解冻肉样修整为1 cm×1 cm×2 cm 肉块置于检测管底部,选择CPMG(Carr-Purcell-Meiboom-Gill)脉冲序列检测T2弛豫图谱。参数设置:使用CPMG 序列,线圈温度32℃,质子的共振频率18 MHz。采样频率SW=200 kHz,模拟增 益RG1=20,P1=16 μs,数字增 益DRG1=3,TD=251 038,PRG=1,重复采样间隔时间TW=3 000 ms,累加次数NS=8,P2=33 μs,回波时间TE=0.251 ms,回波个数NECH=5 000。根据CPMG 指数衰减曲线图,用分析软件进行迭代反演得到横向弛豫时间T2图谱。
1.3.8 质构测定 根据Zhang 等[16]的方法并稍作修改,用质构仪TPA 模式对解冻肉样(1 cm×1 cm×2 cm)进行全质构测定,每个处理做3 次平行试验。将肉样置于TPA 平板上做2 次压缩试验,压缩距离为10 mm,触发力为5 g,间隔时间5 s,测前速率为2.0 mm/s,测中和测后速率为1.0 mm/s,方向平行于肌纤维,探头型号为P/50。测定完毕用TPA-macro 软件对测定结果进行分析。
1.3.9 嫩度测定 根据Bai 等[20]的方法并稍作修改,用质构仪剪切力模式对解冻肉样(1 cm×1 cm×2 cm)进行嫩度测定,每个处理做3 次平行试验。参数设置为:距离为30 mm,触发力为20 g,测前和测中速度为2.0 mm/s,测后速度为10.0 mm/s,方向垂直于肌纤维,探头型号为HDP/BSW 探头。
1.3.10 透射电子显微镜观察 将样品切至0.5 cm×0.5 cm×0.5 cm 块状,经3%戊二醛预固定,1%四氧化锇再固定,丙酮逐级脱水,脱水剂体积分数依次为30%,50%,70%,80%,90%,95%,100%(100%浓度中换3 次),将脱完水的样品先后经过脱水剂和环氧树脂(型号为Epon812)渗透液,体积比分别为3∶1,1∶1,1∶3,每步30~60 min。将渗透好的样品块放到适当模具中,灌上包埋液包埋经过加温聚合形成一种固体基质(包埋块),已备下一步切片。采用超薄切片机制备约60~70 nm 厚的超薄切片后,漂浮于刀槽液面上,再捞至铜网,先用醋酸铀染色10~15 min,再用枸橼酸铅染色1~2 min,室温下染色。采用JEM-1400FLASH透射电镜对铜网进行图像采集,每张铜网先于6 000 倍下观察,选择要观察的区域采集图片。
1.4 数据分析
使用IBM SPSS Statistics 26(SPSS Corp,Chicago,USA)软件进行显著性分析(P 〈 0.05),使用Microsoft Excel 2019 软件计算平均值和标准偏差,采用Origin 2021 作图。
2 结果与分析
2.1 共晶点测定结果
目前检测物料共晶点的方法有两种,分别为DSC 法和电阻法,其中DSC 法是利用物料在发生相变时会发生相应的焓变,导致物料在该温度下有一定的放热或吸热过程,通过仪器记录放热或吸热过程随着时间的变化关系来检测这个共晶点温度[5]。样品在降温的过程中,其内部的游离水会大量结晶,并放出大量的焓变热,检测到手抓羊肉样品DSC 曲线如图1 所示,在整个降温过程中只有一个突出的放热峰,从而得出样品的共晶点温度为(-9.66±0.24)℃。

图1 样品DSC 降温曲线Fig.1 DSC cooling curve of sample
2.2 冻结温度对手抓羊肉冻结曲线和冻结速率的影响
冻结速率是影响肉制品品质的重要因素,与冰晶体形貌和水分的迁移有着密切的关系[21]。图2为手抓羊肉在不同冻结温度(表1)下的冻结曲线,揭示了不同温度冻结的手抓羊肉中心温度随时间的变化规律,两端下降较快,中间较为平坦,符合一般食品冻结曲线的趋势。目前在进行冷冻操作时,力求尽快通过最大冰晶生成带,以减少产品营养价值、风味以及结构受损。冻结曲线中较为平坦的阶段为最大冰晶生成带,图中可见共晶点温度位于最大冰晶生成带之后。
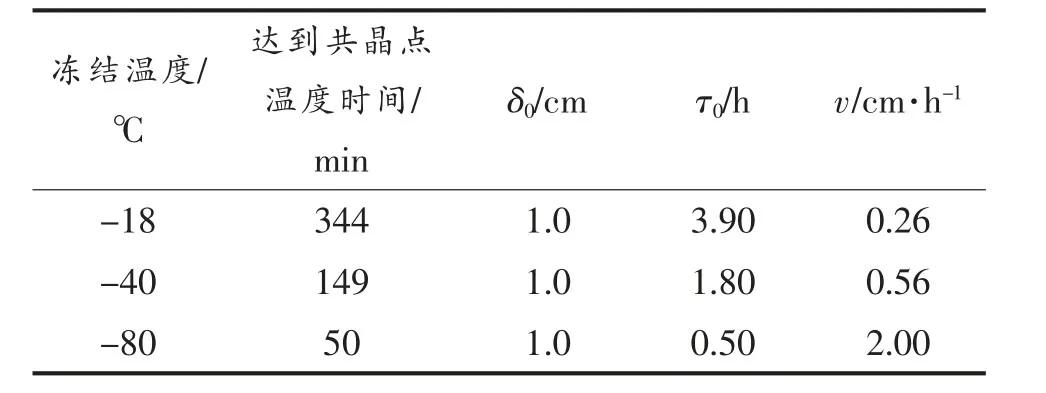
表1 不同冻结温度下的冻结特性Table 1 Freezing features of different freezing temperature
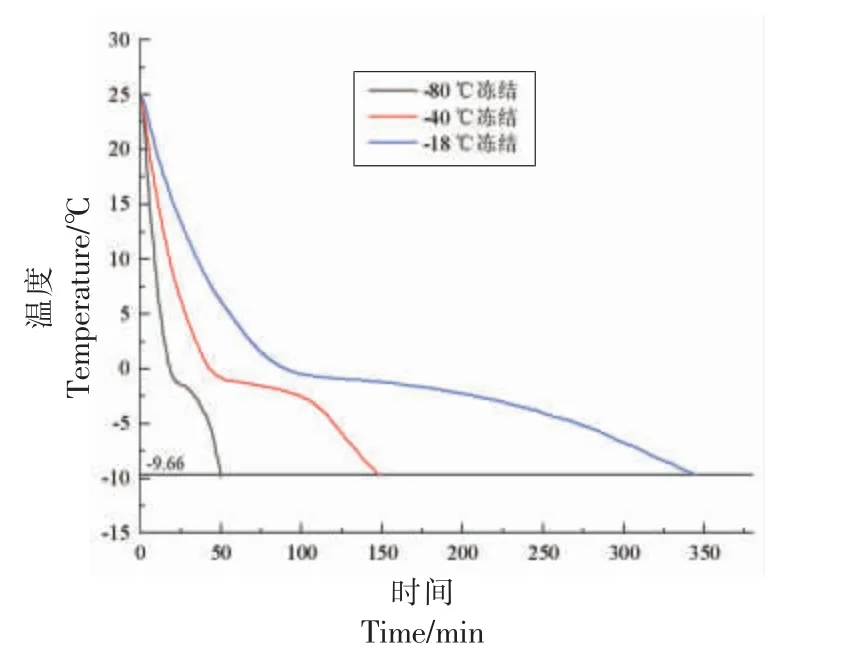
图2 不同冻结温度的冻结曲线Fig.2 Freezing curves of different freezing temperature
由表1 可知,冻结速率随冻结温度的降低而逐渐增大,-18 ℃冻结速率为0.26 cm/h,-40 ℃冻结速率为0.56 cm/h,-80 ℃冻结速率为2.00 cm/h。-40 ℃和-80 ℃由室温达到共晶点的时间远短于-18 ℃和-80 ℃条件下达到共晶点的时间,仅为50 min。
2.3 不同冻结方式的手抓羊肉品质特性变化分析
不同冻结方式的手抓羊肉品质特性具有一定差异,具体如下:
色泽对肉制品的外观和可接受性有重要影响。如图3 所示,与对照组相比,-80 ℃冻结L*值无显著差异(P〉0.05),而-18 ℃冻结与-40 ℃冻结样品L*值显著高于-80 ℃冻结(P〈0.05),这是由于肉类在冻结过程中,L*值受冻结速率的影响,与解冻样品中的水分状态和分布有关[17],冻结速率慢造成解冻损失较大,光反射更强,亮度更高。Muela 等[22]研究证明,慢速冷冻的羊肉解冻后比快速冷冻颜色更暗。此外,-80 ℃冻结样品的肌纤维收缩程度最低,这有助于减少光在肉表面的散射,增加L*值。此外,对照组的a*值显著高于冻结组(P〈0.05),b*值显著低于冻结组(P〈 0.05),冻结组间a*值与b*值差异均不显著(P〉0.05)。Wang等[23]研究表明,肉饼冻融后a*值显著降低,L*和b*显著升高。一般a*值的变化与肌红蛋白的氧化与还原过程有关,b*值显著升高可能是冻融过程温度变动使脂肪氧化导致。

图3 不同冻结方式手抓羊肉色泽对比Fig.3 Color comparison of hand grab mutton with different freezing methods
解冻损失率与肉制品的外观、质量、色泽和感官质量密切相关[24],能够反映冻结-解冻后肉中细胞和组织结构受损程度,肉样中组织和细胞的结构受损越严重,汁液流失越多,随之损失的营养物质也越多。冻结过程冰晶形成所产生的物理挤压和刺破作用,是造成冷冻肉制品汁液流失的主要原因[25-26]。Coleen 等[27]发现,冰晶形成后肌纤维的间距随之改变,造成原有空间容纳的水分子流失,降低了肉的保水性。Cai 等[28]也证实,冷冻储存导致样品肌纤维排列松散,损伤严重甚至出现凹坑和裂缝,造成一定程度的失水。由图4 可知,-80℃冻结样品解冻损失率最低,各组样品的解冻损失率差异显著(P〈0.05),说明快速冻结提高了冻结过程中手抓羊肉的保水性。解冻损失与冻结速率、冰晶位置和大小以及肌肉组织的完整性有关[29-30],冻结过程中冰晶生长导致的肌肉纤维断裂会降低肌肉的水结合能力[31]。-80 ℃冻结样品中解冻损失最低,与肉中形成的细小而均匀的冰晶有关,这会减少冻结对肌肉结构的损伤。对于-40 ℃与-18 ℃冻结样品来说,缓慢的冷冻速度导致水迁移到细胞外空间,这可能导致不规则的大冰晶的形成和肌肉结构的破坏[22]。因此,以较低速率冻结的样品在解冻后将融化的水重新吸收回细胞的能力较弱。

图4 不同冻藏条件手抓羊肉解冻损失率对比Fig.4 Comparison of thawing loss rate of hand grab mutton with different freezing methods
图5 为不同方式冻结的手抓羊肉解冻后水分含量对比情况,对照组含水量显著高于其它组(P〈 0.05),这是由于对照组样本未解冻失水。在经过冻结-解冻后,只有-80 ℃组含水量较高,这是由于-80 ℃冻结形成的冰晶更加均匀、细小、致密,对组织造成的机械损伤较小,解冻后水分保留率较高。

图5 不同冻结方式手抓羊肉水分含量对比Fig.5 Comparison of moisture content of hand grab mutton with different freezing methods
质构和嫩度是评价肉质的重要指标,与肉的含水量密切相关。如表2 所示,在嫩度方面,与对照组相比,剪切力有增加趋势,-18 ℃和-40 ℃组肉样剪切力显著增加,-80 ℃组剪切力变化不显著,说明较高的冻结速率能维持手抓羊肉的嫩度,慢速冻结的手抓羊肉嫩度会下降。研究发现,当肌原纤维损伤严重时,会造成肉制品水分大量流失,剪切力增加,嫩度降低[29,31-33]。肉的质构在解冻后会发生显著变化,而质构特性的微小变化会影响整体质量。在质构特性方面,解冻后硬度值、咀嚼性显著增加,内聚性、弹性、回复性显著降低。随着冻结速率增加,硬度值、咀嚼性呈下降趋势,内聚性、弹性、回复性呈上升趋势。Wang 等[23]发现,冻融后肉饼硬度上升,黏结性、咀嚼性和弹性均显著降低,这可能与冻融过程冰晶生长引起的汁液流失有关,本文研究结果与其具有一致性。
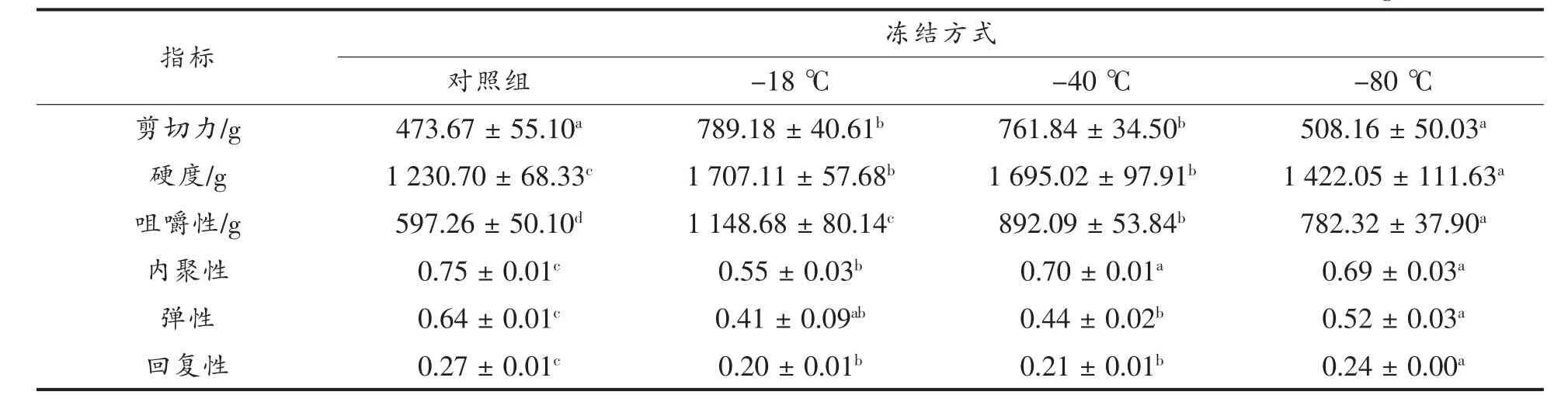
表2 不同冻结方式手抓羊肉嫩度和质构特性对比Table 2 Comparison of tenderness and texture characteristics of hand grasping mutton with different freezing methods
2.4 不同冻结方式手抓羊肉低场核磁结果分析
1)不同冻结方式手抓羊肉水分横向弛豫时间T2的变化 低场核磁共振是一种无损、无创的光谱技术,可用于评估水分子在肉制品中的迁移和分布[32]。近年来,基于LF-NMR 对生肉中水分变化的研究越来越多。然而,很少有研究分析肉制品解冻后水分分布与迁移规律的变化,因此本研究可以补充和验证冷冻肉制品的低场核磁共振分析结果。不同冻结方式的手抓羊肉解冻后,低场核磁共振弛豫时间分布曲线可以观察到3 种类型的峰,其中T2b对应与大分子紧密结合的结合水,T21代表被困在肌原纤维蛋白质网络中的固定水,T22对应存在于纤维束之间中的自由水[34]。为了进一步研究不同冻结方式手抓羊肉的品质变化,采用低场核磁共振技术对其内部水分分布情况进行检测。
弛豫时间T2表征不同状态的水分流动性,弛豫时间的变化可以表明手抓羊肉水分活度的变化,T2越大,与大分子结合力越小,水分自由度越大,流动性强;T2越小,流动性越差,与大分子结合越紧密,水分自由度越小[34]。自由水的弛豫时间一般在100~1 000 ms 之间,这是因为自由水相对于结合水和不易流动水更容易从磁场中吸收能量而产生振动,而当磁场消失后,自由水要恢复到基态需要更长的时间。结合水最难从磁场中获取能量,因而恢复到基态所需的时间最短,一般在1 ms 左右。而不易流动水则介于自由水和结合水之间,一般在10 ms 左右[35]。
图6 显示了不同冻结方式的手抓羊肉解冻后T2弛豫特性的变化。与对照组相比,冻结样品的不易流动水和自由水对应的弛豫时间T21和T22均发生了不同程度的右移(弛豫时间延长)。T21和T22右移程度表现为-18 ℃冻结〉-40 ℃冻结〉-80 ℃冻结。在冻结过程中,巨大的细胞外冰晶会破坏肌肉组织的物理结构。Isabel 等[36]也发现了类似的结果,他们发现随着冻结温度的降低,鳕鱼肌肉的T21显著增加。在冻结过程中,巨大的细胞外冰晶会破坏肌肉组织的物理结构。肉解冻后,受损的肌原纤维难以重新吸收细胞外空间中的融化水,从而导致部分固定水转化为自由水。在本研究中,-80 ℃冻结对肌肉纤维的损伤最小,因为它们的冰晶较小且分布均匀,因此,T21比-40 ℃冻结和-18 ℃冻结短。Renou 等[37]研究表明,弛豫时间与猪肉的品质特征之间存在关系,他们证实T2弛豫时间较长的肌肉样品具有较高的解冻损失。与-40 ℃冻结和-18 ℃冻结样品相比,-80 ℃冻结样品中的自由水流动性较低,与肌肉组织的结合更紧密,从而减少了解冻损失,这与前文解冻损失率的结果一致。

图6 不同冻结方式手抓羊肉水分弛豫时间变化Fig.6 Changes in the water transverse relaxation time of hand grab mutton with different freezing methods
2)不同冻结方式手抓羊肉各形态水占比的变化 不同冻结方式手抓羊肉LF-NMR T2图谱各状态水分占比(M2b、M21、M22)的变化如表3 所示。
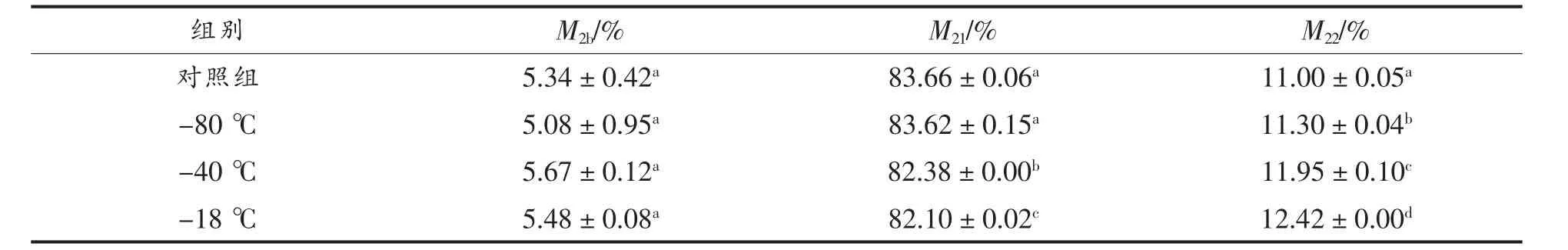
表3 不同冻结方式手抓羊肉各状态水占比的变化Table 3 Changes in the percentages of water in various forms of hand grab mutton with different freezing methods
3 种冻结方式中,冻结速率高的样品不易流动水占比M21显著高于低速率冻结(P〈0.05),表明高速率冻结样品解冻后可以结合更多的不易流动水。手抓羊肉M2b变化不显著(P〉0.05),这是因为结合水与大分子物质结合紧密[31]。此外,冻结速率高的样品具有更低的自由水占比M22(P〈0.05),这可能是由于羊肉内部冰晶融化,水分流失加快,表面湿润,对羊肉内部水分损失具有促进作用,这可以用于解释前文中低速率冻结手抓羊肉具有更高的解冻损失率的结论。
2.5 不同冻结方式手抓羊肉透射电子显微镜观察结果分析
透射电子显微镜观察到的是肌原纤维的纵截面。通过肌原纤维间空隙的大小,以及肌原纤维的完整性来判断肌肉组织中形成的冰晶大小[38]。
图7a 是未经冷冻的新鲜手抓羊肉,可以观察到其肌原纤维间几乎没有空隙,而冻结组(图7b~图7d)均出现空隙,说明冻结期间冰晶的形成导致肌原纤维分离。各冻结组间微观结构也有较为显著的差异,与图7a 相比,图7d 中出现均匀、狭细的空隙,图7b 和图7c 中空隙较大,且均一程度差。冻融后手抓羊肉肌原纤维间距不同的直接原因是不同冻结速率造成肌原纤维间水分凝结成的冰晶大小不一。

图7 不同冻藏条件手抓羊肉透射电子显微镜图Fig.7 TEM micrographs of hand grab mutton with different freezing methods
前文结果表明手抓羊肉未经冻结时,水分含量大约是52%左右,肉制品中水分主要存在于肌原纤维中、肌原纤维间、肌原纤维与细胞膜之间、细胞间和肌束之间的空隙中。水作为两性分子能与蛋白质分子中的电荷相互吸引从而与蛋白质分子相结合,这部分水称为结合水,含量基本恒定,这与低场核磁结果相对应。不易流动水受空间效应的影响而存在于肌肉中,部分与结合水结合,而不与蛋白质结合,这部分水易被冻结。快速冷冻时,水分子在过冷状态停留时间短,能很快通过最大冰晶生成带,通常形成的冰晶小且分布均匀,使肉制品接近新鲜状态时的水分分布[39]。而低速冷冻时,有足够的时间让液态水向正在生长的晶体扩散,形成较大的冰晶体[40]。冰晶的生长会加速破坏肌原纤维的完整性,肌原纤维蛋白维持着肉制品紧凑而均匀的三维凝胶结构[41-42]。研究发现,肌原纤维的凝胶强度随冻融循环次数的增加而降低,这可能是冰晶的生长导致蛋白氧化变性、结构破坏,从而降低了蛋白成胶能力[27,43],本文手抓羊肉解冻后仍保有较高的肌原纤维完整性,Z 线、明带、暗带皆清晰可辨,这可能是由于本研究样品仅经历了一个温度稳定的冻融循环,肌原纤维破坏程度较低,只因冻结速率影响而形成了大小程度不同的冰晶,导致不同冻结方式的手抓羊肉肌原纤维间隙的差异。
2.6 手抓羊肉品质指标变化相关性分析
对手抓羊肉的品质特性进行相关性分析,结果见图8。
由图8 可知,解冻损失、水分含量与横向弛豫时间T2b、T21、T22、不易流动水占比M21、自由水占比M22显著相关(P〈0.05),说明手抓羊肉中宏观水分变化与微观各形态水分布与迁移密切相关。质构特性、嫩度与解冻损失、水分含量、横向弛豫时间T2b、T21、T22、不易流动水占比M21、自由水占比M22显著相关(P〈0.05),说明手抓羊肉的组织结构变化与水分变化密切相关。L*值、a*值、b*值与解冻损失、水分含量、横向弛豫时间T2b、T21、T22、不易流动水占比M21、自由水占比M22显著相关(P〈0.05),说明色泽变化与肉中水分变化密切相关。
3 结论
较高的冻结速率对维持手抓羊肉色泽、嫩度、质构、持水能力、微观结构等方面具有显著的积极作用。冻融会对肉的品质指标产生不利影响,如水分子自由度上升,肌原纤维完整性被破坏等,从而导致水分流失加剧,色泽、嫩度及质构特性变差。高速率冻结可以使手抓羊肉中产生的冰晶更为细小、均匀、致密,从而延缓冰晶对肉品质的破坏进程,降低肉的品质劣变程度,使手抓羊肉在冻结后仍能最大限度保有新鲜煮制时原有的品质。在本文研究的3 种冻结方式中,-80 ℃冻结具有最高的冻结速率,同时也具有最佳的品质保真效果,且以共晶点作为冻结终温,达到冻结终点的时间短,可为工业化生产过程成本控制提供现实性指导。
猜你喜欢
航空学报(2023年1期)2023-01-31
科普童话·学霸日记(2021年12期)2021-05-30
模具制造(2019年3期)2019-06-06
现代营销(创富信息版)(2018年6期)2018-09-05
含能材料(2017年1期)2017-03-04
含能材料(2017年7期)2017-03-04
琴童(2016年5期)2016-05-14
当代化工研究(2016年6期)2016-03-20
食品工业科技(2014年23期)2014-03-11
食品工业科技(2014年23期)2014-03-11
