
植物多酚调控肠道微生态改善炎症性肠病的研究进展①
2023-11-30 02:09闫成花况文东廖永翠万红娇马广强
中国免疫学杂志 2023年11期
闫成花 况文东 廖永翠 丁 坚 靳 亮 万红娇 马广强
(江西中医药大学中医学院/生命科学学院,南昌 330004)
炎症性肠病(inflammatory bowel diseases,IBD)是一种回肠和结肠中含有大量炎症因子导致的自身免疫病,也是一种病因较为复杂的胃肠道紊乱性疾病,主要包括溃疡性结肠炎(ulcerative colitis,UC)和克罗恩病(Crohn´s disease,CD)[1-2]。UC 主要表现为结肠及直肠内壁长时间的炎症和溃疡,而CD 主要表现为整个消化系统存在大量炎症。无论是UC还是CD,患者都伴有严重腹泻、腹痛、便血和体重减轻,且慢性长期炎症也是癌症,如消化系统癌症的主要诱因,临床发现肠炎患者结直肠癌发病率高于健康人[3-4]。一直被视为西方世界疾病的IBD 目前已经全球化,发病率不断升高[5-6]。IBD 是一种慢性进展性疾病,需长期治疗,且治疗药物昂贵,给社会带来了沉重的经济负担。
IBD 难以治愈且易复发,其病因尚不清楚,既具有遗传倾向,又与环境因素、免疫因素、肠道菌群等密切相关[7]。目前常用治疗手段主要为手术切除炎症组织或采用抗炎药物。重症患者治疗主要依赖于肿瘤坏死因子(tumor necrosis factor α,TNF-α)等中和性抗体或免疫抑制剂,尽管TNF-α 拮抗剂在治疗IBD 方面疗效显著,但10%~30%患者对该药物不敏感,30%~50%应答患者用药1 年后产生耐药性[8-9]。这种治疗手段仅能进行病症缓解而无法根治,且很多药物使用后会出现明显副作用[10]。手术治疗也有并发症风险,如盆腔炎、大出血和肠穿孔,且受患者年龄和身体状况限制[11]。因此,IBD 目前仍无有效治疗手段。植物多酚化合物具有抗炎、抗菌及抗氧化等多种生物学活性,在多种疾病中发挥重要作用,尤其是炎症性疾病,本文旨在探讨植物多酚调节肠道菌群在IBD中的研究进展。
1 IBD的发病机制
IBD 发病机制复杂,主要受肠道菌群、遗传及环境等因素影响。人体肠道微生物群由1 000 多种微生物组成,主要分为4个门:厚壁菌门、拟杆菌门、变形菌门和放线菌门。健康个体的肠道微生物群对宿主有益,可促进机体消化吸收、合成维生素,为机体提供营养物质,还可辅助清除病原微生物,保护肠黏膜[12]。肠道微生物群失调包括有益菌减少、有害菌增加以及微生物多样性减少,将改变宿主-微生物相互作用,影响宿主免疫系统及营养物质吸收代谢,进而破坏肠道上皮黏膜屏障,促进肠道炎症[13-14]。随着GWAS 技术和二代测序技术发展,已确定了240 多个与IBD 相关的非重叠遗传风险位点,其中约30 个基因位点是CD 和UC 共有的[15-16]。与IBD 相关的易感基因包括IL-23R、NOD2、自噬相关基因ATG16L1和IRGM、IL12B、JAK2、STAT3、CARD15等[17]。IL-23R是第一个通过GWAS 鉴定出的IBD 相关基因,GWAS 分析发现IL-23R中存在多个与CD 和UC 高度相关的单核苷酸多态性(single nucleotide polymorphism,SNP)[18]。核苷酸结合寡聚结构域2(nucleotide-binding oligomerization domain 2,NOD2)是NOD 家族成员之一,约1/3的CD 患者会发生NOD2突变[19]。NOD2 突变后,NF-κB 信号通路异常激活,从而诱导大量炎症因子产生。自噬相关基因在IBD 中也具有重要作用,ATG16L1编码的T300A 突变促进SQSTM1/p62 积累,TRAF6 及RIPK2多泛素化增强,使细胞具有异常的TLR 信号[20]。除NF-κB 信号通路和自噬相关信号通路外,JAK/STAT和TGF-β1/SMADs 信号通路也参与IBD 发生发展[21]。研究表明,许多环境因素与IBD 发病密切相关。调查显示,空气污染物排放总量与成人 IBD 住院人数直接相关[22]。大数据分析发现,与从不吸烟者相比,吸烟者患CD 风险增加,患UC 风险降低[23]。此外,扁桃体切除术、阑尾切除术、抗生素使用、口服避孕药、维生素D缺乏、不良饮食等也是与IBD 发生密切相关的环境因素[24]。高脂肪、高蛋白、高糖、低纤维饮食与IBD发病率呈正相关。角叉菜胶和表面活性剂等食品添加剂可能增加患IBD风险。饮食中的软饮料、饱和脂肪、红肉和麸质含量也有助于IBD发展[25]。
2 肠道菌群在IBD中的作用
越来越多的证据表明,肠道微生物群失调与人类IBD 发展有关。与健康受试者相比,IBD 患者肠道中微生物群组成发生改变,主要为厚壁菌门和拟杆菌门比例降低,变形杆菌比例升高[26]。乳酸杆菌等有益菌可减少结肠中的氧化应激,IBD 患者肠道中有益菌比例降低导致活性氧过度产生,进一步加剧肠道菌群失调[27]。CD和UC患者中黏附侵袭型大肠杆菌比例分别为75%和69%,远高于健康人,侵袭型大肠杆菌等有害菌通过与肠道肠腔内免疫细胞相互作用促进IBD,甚至导致结直肠癌[28]。除肠道微生物组成及比例变化影响IBD 外,微生物多样性也参与IBD 调节。与健康个体相比,IBD 患者肠道微生物群多样性显著降低,多样性降低主要来自厚壁菌门,CD 患者中仅有13 个厚壁菌门,而健康受试者中有43 个[29]。此外,微生物代谢产物在IBD 中也具有重要作用。研究发现,IBD 患者中产生短链脂肪酸(short-chain fatty acid,SCFA)的菌株减少,SCFA 作为结肠上皮细胞的主要能量来源,可调节免疫细胞,减少促炎因子,缓解IBD[30]。梭状芽孢杆菌参与色氨酸代谢,将色氨酸分解为吲哚衍生物,UC患者色氨酸代谢失调,产生的吲哚衍生物明显减少[31]。次级胆汁酸主要为胆汁酸在肠道微生物作用下转变而来,IBD 患者中次级胆汁酸产生明显减少[32]。
肠道中有大量CD4+T 细胞定居,肠道微生物群通过调节CD4+T 细胞亚群分化调控IBD 发生发展。研究发现分段丝状杆菌(SFB)定植水平与肠道中Th17丰度密切相关,SFB特异性抗原被表达MHC-Ⅱ的CD11c+DCs 识别,作用于下游Th17,促进Th17 分化[33]。梭菌属Ⅳ簇和ⅩⅣa 簇细菌可提供促进Treg分化的环境,并促进Treg 表达iCOS,分泌IL-10,减轻结肠炎[34]。微生物群衍生代谢物SCFA 中的丁酸盐刺激CD103+DCs 产生高水平的TGF-β,TGF-β 与DCs(GPR109A)和CD4+T 细胞(GPR43)的GPCRs 结合,促进T 细胞向Treg 分化[35]。IBD 患者肠道组织中色氨酸代谢物受体(aryl hydrocarbon receptor,AHR)表达增加,Th17 向Treg 转分化一定程度依赖于AHR[36]。肠道胆汁酸及其有效的次级胆汁酸诱导RORγ+Treg对维持肠道免疫稳态至关重要[37]。肠道菌群也是肠道黏膜屏障的重要组成,肠道黏膜屏障具有防止微生物侵入、维护肠道健康的作用,同时为肠道微生物提供营养和栖息地。反过来,肠道微生物群调节黏蛋白产生和分泌影响其功能,如将益生菌菌株鼠李糖乳杆菌CNCM Ⅰ-3690 用于小鼠,可通过增加黏液产生和恢复杯状细胞群改善结肠屏障功能[38]。肠道菌群失调会增加肠道通透性,影响肠道免疫,最终导致肠道黏膜屏障损伤,引起IBD[39]。
综上,肠道微生态失调是调控IBD的关键因素,肠道菌群失调会改变宿主与微生物相互作用。因此,促进肠道有益细菌、减少有害菌、增加微生物多样性可有效调节Th17/Treg平衡,促进黏膜屏障完整性,是缓解结肠炎的有效策略。
3 常用的IBD临床治疗药物
过去几十年中,人们进行了深入研究阐明IBD发病机制,寻找有效的IBD 治疗药物靶点。常用临床药物包括皮质类固醇、氨基水杨酸盐、免疫抑制剂、黏附分子及整合素拮抗剂、TNF 抑制剂和IL-12/IL-23 抑制剂等,旨在抑制肠道炎症,修复消化道黏膜组织,从而改善IBD 患者生活质量[40]。但这些治疗药物主要是快速缓解疾病,在疗效维持方面效果不佳,且长期使用副作用较大。皮质类固醇(强的松、氢化可的松、布地奈德、地塞米松)主要用于中重度IBD治疗,尤其是急性发作期患者,长期使用抑制下丘脑垂体肾上腺轴,导致睡眠和情绪障碍[41]。一些氨基水杨酸药物,如美沙拉秦、奥沙拉嗪和柳氮磺胺吡啶,主要用于轻度和中度活动期UC 缓解,该类药物主要不良反应为胰腺炎、肾毒性、肝毒性和性功能障碍等[42]。TNF抑制剂也是最常用治疗药物之一,包括英夫利昔单抗、阿达木单抗和戈利单抗,使用抗TNF-α 药物治疗的患者会增加其他疾病患病风险,如严重感染、恶性肿瘤、心力衰竭等疾病[43]。随着肠道菌群研究深入,口服益生菌及粪便移植等新兴治疗手段逐步用于临床,但临床试验较少,还需深入研究[44-46]。与西药相比,植物多酚具有不良反应少、耐受性好、来源广泛等优点,日渐成为IBD治疗药物的研究热点。
4 植物多酚调节肠道菌群在IBD中的作用
多酚是植物性食物如水果、蔬菜和谷物中的天然化合物,包括黄酮类、酚酸类、单宁类、木脂素类和芪类。黄酮类进一步分为黄酮醇、黄酮、黄烷-3-醇、异黄酮、黄烷酮、花青素和查耳酮等[47]。研究表明,多酚是一种益生元,可以改善肠道微生物群的多样性,增加益生菌如乳酸杆菌、双歧杆菌等,减少有害菌,如侵袭型大肠杆菌、艰难梭状芽孢杆菌等,进而调控肠道菌的代谢产物,如SCFA、胆汁酸等,也可以通过肠道菌群改变影响T细胞分化以及肠黏膜屏障完整性,从而抑制肠道炎症,有效预防和缓解IBD[48]。
多酚对肠道菌群的调控作用主要是通过影响细菌代谢和细菌群体感应抑制遗传物质合成以及干扰细胞膜生物功能等实现。不同微生物新陈代谢不同,梭状芽孢杆菌等肠道细菌生长需要铁,而双歧杆菌和乳酸杆菌生长不需要铁,多酚通过螯合金属离子(铁和钴)抑制细菌酶活性,从而改变微生物代谢,选择性增加双歧杆菌和乳酸杆菌等益生菌生长,促进其在肠道组织定居,并抑制梭状芽孢杆菌等致病菌增殖[49]。多酚可抑制细菌生物膜形成,是在IBD 患者结肠组织中大量存在的一种共生转化的病原菌具核梭杆菌。研究发现1.56 µg/ml(6.8 µmol/L)白藜芦醇可显著抑制具核梭杆菌生物膜形成,从而抑制其生长繁殖[50]。多酚还可通过产生、释放和感应被认为是自诱导剂的小信号分子(革兰阳性菌中的寡肽和革兰阴性菌中的酰化高丝氨酸内酯)破坏细菌群体感应[51]。多酚也可抑制细菌DNA 和RNA 生物合成,据报道,槲皮素作用于大肠杆菌DNA 促旋酶,黄酮类化合物则抑制金黄色葡萄球菌DNA复制必需的解旋酶,从而抑制细菌DNA等遗传物质生物合成[48,52]。此外,儿茶素通过产生H2O2、改变微生物细胞膜通透性发挥抗菌作用[53-54]。肠道微生物如产气荚膜梭菌、粪肠球菌、乳球菌和乳酸杆菌等表达细胞外电子转移(extracellular electron transfer,EET)相关基因,成为肠道环境中多酚的靶标,多酚通过促进这些细菌的EET 或无氧呼吸促进其生长。此外,多酚还可螯合参与EET 的金属离子,如铁和铜,间接影响微生物生长[55]。多酚经微生物群分解产生的代谢物可促进SCFA 合成相关微生物增加,如梭状芽孢杆菌ⅩⅣa簇细菌及双歧杆菌等,促进SCFA 合成[56]。研究发现,EGCG 可抑制上消化道中的α-淀粉酶活性,导致淀粉未完全消化,这些未完全消化的底物可供结肠中合成SCFA的微生物使用,从而促进SCFA 合成[57]。综上,多酚主要通过选择性影响微生物代谢、生物膜形成及功能、电子转移等多种途径调节微生物组成。肠道微生物参与多酚代谢过程,肠道中相对高浓度的多酚可能选择性地影响细菌生长。这种相互作用导致多酚分解为各种代谢物,调节肠道微生物群,并增加SCFA等合成。
研究发现多酚化合物通过与肠道菌群相互作用在调节T细胞及维持肠黏膜完整性中发挥重要作用。白藜芦醇通过逆转TNBS 诱导的肠道菌群失调减少Th17 数量,增加Treg 数量,调节Treg/Th17 平衡,减轻宿主结肠炎。因此,白藜芦醇补充剂是IBD肠道炎症的动态调节剂[58]。槲皮素能通过调节肠道巨噬细胞恢复肠道宿主-微生物平衡,抑制T 细胞活化,促进IL-10 表达,显著降低结肠固有层和肠系膜淋巴结中CD4+T 细胞比例及分泌IFN-γ 和TNF-α的CD4+T细胞比例,最终改善肠炎[59]。此外,槲皮素是AHR 激动剂,通过与AHR 结合抑制炎症性Th17,促进Treg 增殖[60]。在IBD 中,肠黏膜组织Th17 大量浸润,Th17 相关细胞因子表达增加,紫肉马铃薯含有大量花青素,紫肉马铃薯饮食提高小鼠肠道中阿克曼菌相对丰度,并降低结肠中IL-17 mRNA 表达,因此,紫肉马铃薯有助于改善UC 症状[61]。肠道黏膜层为微生物群提供碳和营养源,促进其共生,黏液层成分改变影响肠道微生物群,微生物刺激又是黏蛋白分泌所必需的。多酚-微生物群相互作用产生的多酚分解代谢物和SCFAs 保护肠道黏膜层完整,减轻肠炎。研究发现,白藜芦醇上调claudin-4蛋白表达,降低细胞通透性,阻止大肠杆菌易位[62]。芒果增加了肠道乳杆菌属丰度,促进SCFAs 产生,尤其是丁酸,从而增强黏膜屏障功能[63]。SCFA 通过结合其受体GPR41 和GPR43 提高细胞Ca2+水平,并进一步激活Erk1/2 和Akt 信号,改善上皮屏障功能[64]。已明确报道能够调节肠道菌群缓解肠炎的常见多酚化合物及其作用机制见图1。
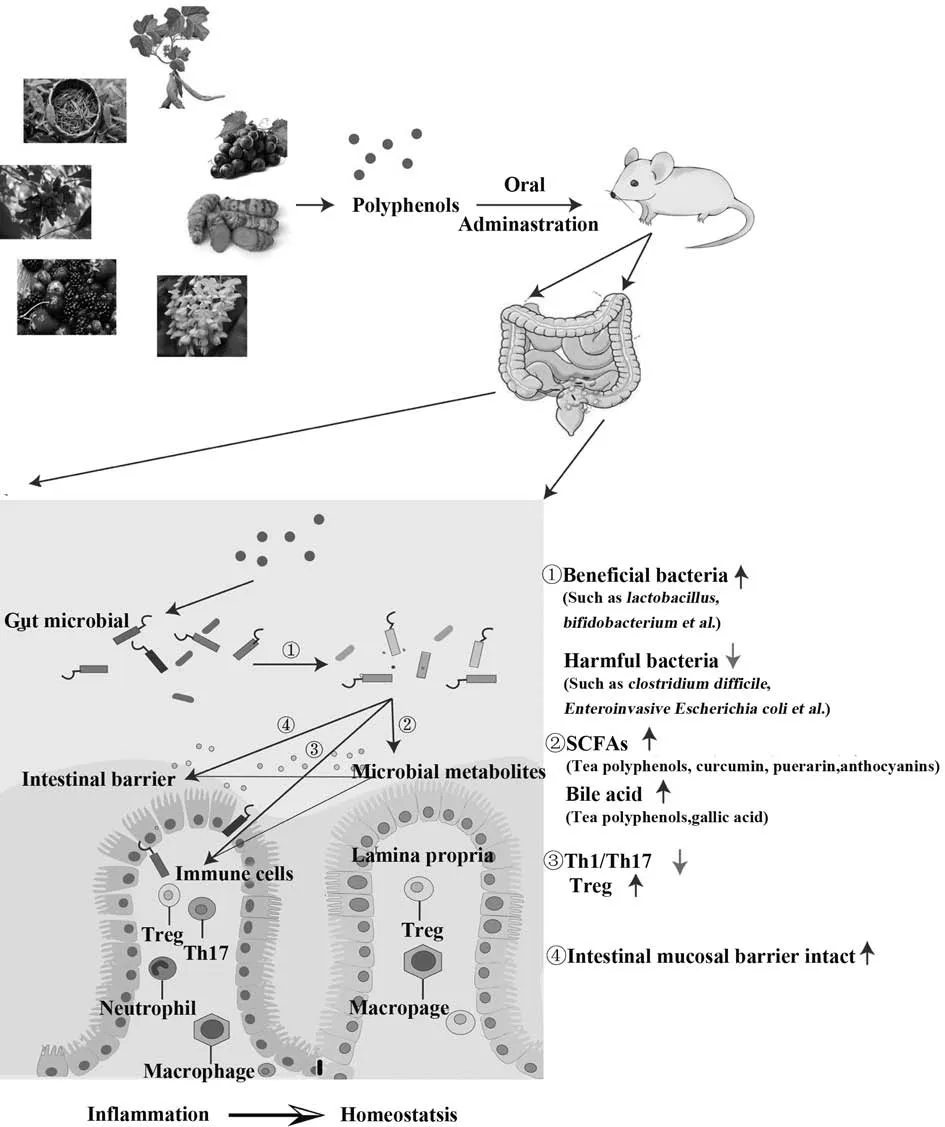
图1 植物多酚通过调节肠道菌群缓解肠道炎症的机制Fig.1 Mechanism of plant polyphenols alleviating intestinal inflammation by regulating gut microbiota
4.1 茶多酚(green tea polyphenols,GTPs)GTPs是存在于食品和药用植物中的天然多酚化合物,是绿茶发挥作用的主要活性成分。GTPs 主要包括类黄酮(儿茶素)、单宁、茶黄素和酚酸。越来越多的研究表明,摄入富含GTPs的食物可预防和治疗人类慢性疾病,如IBD。GTPs 通过与肠道菌群相互作用促进有益菌双歧杆菌生长,促进SCFA 合成,抑制有害的梭状芽孢杆菌生长,从而维持肠道健康[65-66]。GTPs 也可增加肠道中产生次级胆汁酸的微生物丰度,如拟杆菌属和双歧杆菌,促进次级胆汁酸产生,调节免疫细胞,抑制肠道炎症[67]。IBD 患者肠黏膜中拟杆菌门和厚壁菌门数量显著减少,而变形杆菌门和放线菌门数量显著增加,饮食中加入GTPs可显著改善IBD 模型小鼠结肠炎症[68]。儿茶素是GTPs的主要成分,也具有调节肠道菌群、善肠道微环境、缓解IBD的作用[69]。研究表明表没食子儿茶素没食子酸酯(epigallocatechin gallate,EGCG)可通过与组蛋白样DNA 结合蛋白结合有效抑制幽门螺杆菌生长[70]。初步临床研究发现,健康受试者使用GTPs后,粪便中梭状芽孢杆菌等细菌减少,双歧杆菌和乳酸杆菌增加[71]。UC 患者口服EGCG 可缓解对5-氨基水杨酸和硫唑嘌呤的耐药性,且副作用小,有望成为治疗轻度至中度活动性UC 患者的新选择[72]。此外,与常用临床药物柳氮磺吡啶相比,EGCG 显著提高抗氧化水平并缓解结肠炎症状,且EGCG 是IBD 和健康个体淋巴细胞中氧化应激的拮抗剂[73]。GTPs 是否通过调节肠道菌群缓解IBD 尚缺乏临床证据。
4.2 白藜芦醇 白藜芦醇(3,5,4´,三羟基反式芪)是一种天然多酚(芪类),主要存在于虎杖、决明子、桑树等常见药用植物以及葡萄、花生和石榴等食物中,具有广谱药理活性,如抗炎、抗癌和抗氧化等,可用于治疗多种疾病,包括糖尿病、组织损伤、炎症性疾病和癌症[74-76]。研究表明白藜芦醇显著促进乳酸乳球菌生长,并抑制粪肠球菌生长,从而正向调节宿主免疫,降低炎症反应[27]。白藜芦醇处理葡聚糖硫酸钠(dextran sulphate sodium,DSS)诱导的大鼠,可增加大鼠肠道中双歧杆菌和乳酸杆菌丰度,抑制大肠杆菌增长,并减少一氧化氮合酶表达,使DSS 损伤的肠黏膜屏障得以修复[73]。同样,补充白藜芦醇可增加DSS诱导的结肠炎小鼠粪便中的有益菌乳酸杆菌和双歧杆菌,并减少包括嗜胆菌属在内的潜在致病菌[77]。白藜芦醇通过调节肠道菌群与机体免疫系统相互作用诱导Treg 产生,抑制Th1/Th17 产生,保护宿主免受结肠炎影响[78]。白藜芦醇通过促进有益菌阿克曼氏菌增殖及代谢产物丁酸产生,显著缓解TNBS 诱导的结肠炎[58]。研究发现,与对照小鼠相比,结肠炎小鼠长期摄入白藜芦醇后,肠道中厚壁菌门/拟杆菌门比例和乳杆菌属丰度显著升高,肠道微生物群组成在控制白藜芦醇代谢中起重要作用,此外,厚壁菌门/拟杆菌门比例显著升高,进一步促进二氢白藜芦醇代谢[79]。临床研究发现,饮用富含白藜芦醇的红酒后,代谢综合征(肥胖)受试者粪便微生物增加,尤其是有益菌乳酸杆菌和双歧杆菌显著增加,而炎症标志物水平显著降低[80]。白藜芦醇与肿瘤治疗常用化疗药物一起服用显著降低常规化疗药物引起的不良反应及耐药性[81]。一项随机双盲临床试验表明,50 例活动性轻度至中度UC 患者连续6 周每天补充500 mg 白藜芦醇,可显著降低血浆TNF-α 水平,临床结肠炎活动指数得分显著下降,表明白藜芦醇可部分通过调节炎症因子表达有效缓解UC[82]。另有临床试验发现,白藜芦醇可提高轻度至中度UC 患者抗氧化能力并降低氧化应激,改善UC 患者疾病活动性和生活质量[83]。白藜芦醇可通过调节肠道微生物群改变抗氧化、抗炎和免疫应答,显著改善肠道功能,降低结肠炎及结直肠癌风险和发病率。
4.3 姜黄素 姜黄素最早是从植物姜黄中分离到的多酚(黄酮类),广泛分布于药用植物姜黄、莪术和菖蒲的根茎中,是一种强抗氧化剂、抗炎剂、抗菌剂和抗病毒剂,常用于常见疾病治疗,尤其用于治疗胆道疾病、风湿病、糖尿病、肠炎、关节炎等[84-85]。姜黄素具有调节肠道菌群、改变肠道微生物丰富度和多样性作用,通过维持产生SCFAs 的细菌数量和比例促进SCFAs 合成,保护肠黏膜,抑制肠道炎症[86]。通过给予结肠炎小鼠模型姜黄素纳米颗粒,显著增加产生丁酸盐的微生物群,从而诱导Tregs及免疫耐受性T细胞生成,改善肠道屏障功能,减少肠道炎症[87]。利用IL-10 缺陷小鼠构建结肠炎模型,发现姜黄素处理可防止年龄相关的细菌α多样性降低,增加细菌丰富度,并防止结直肠癌发展[88]。此外,姜黄素有助于调节一些特定的肠炎高度相关细菌丰度,如阿克曼氏菌、粪肠球菌、罗氏菌属及F16、肠球菌科等;通过增加紧密连接蛋白如occludin、ZO-1、claudin-3 表达增强肠道屏障;通过抑制caspase-3 通路减轻DSS 诱导的肠道细胞凋亡;通过抑制MAPK/NF-κB/STAT3 通路减轻肠道炎症[89]。目前姜黄素对人肠道微生物群的影响尚缺乏研究。常规临床药物治疗患者易复发,一项针对姜黄素预防复发的随机、双盲临床试验也显示,接受姜黄素和常规治疗药物治疗的43 例患者中,2 例在治疗6 个月内复发(4.65%),而安慰剂组39 例患者中有8 例(20.51%)复发,表明服用姜黄素的患者复发率显著降低[90]。
4.4 槲皮素 槲皮素是一种常用的具有多种生物学活性的天然多酚(黄酮类),通常以糖基形式存在,分布广泛,100 多种药用植物(如槐米、侧柏叶、高良姜、款冬花、桑寄生、三七、银杏、接骨木等)均含有此成分。COMALADA 等[91]在实验大鼠中观察到槲皮素对DSS 诱导的结肠炎具有保护作用,肠道微生物群分解糖苷类时释放槲皮素,槲皮素通过抑制NF-κB 信号通路抑制细胞因子产生并诱导一氧化氮合酶表达,减轻DSS 诱导的结肠炎。槲皮素还可调节肠道菌群,增加有益菌如双歧杆菌和乳酸杆菌数量,显著降低艰难梭状芽孢杆菌和肠球菌数量。槲皮素处理DSS 诱导的结肠炎模型小鼠,发现小鼠体质量明显增加,氧化应激明显减弱,厚壁菌门细菌数量增加,变形菌门细菌数量减少,表明槲皮素可通过调节氧化应激改善肠道菌群缓解小鼠UC[92]。幼稚T 细胞过继转移到Rag1 缺陷小鼠并诱导T 细胞依赖性结肠炎,该小鼠补充槲皮素后拟杆菌门/厚壁菌门比例升高,潜在致病微生物分段丝状细菌和大肠杆菌相对丰度降低[59]。研究发现槲皮素对柠檬酸杆菌诱导的结肠炎小鼠也具有很好的缓解作用,槲皮素增加拟杆菌属、双歧杆菌属和乳杆菌属数量,并显著减少梭杆菌属和肠球菌属数量,抑制促炎细胞因子如IL-17、TNF-α 和IL-6 产生,促进结肠组织中抗炎细胞因子IL-10 产生[93]。槲皮素针对各种代谢综合征(如糖尿病、肥胖和某些类型癌症)进行了大量临床试验,但在IBD临床研究中尚未见报道。
4.5 葛根素 葛根素是从传统中药材葛根中分离的异黄酮类衍生物,具有抗炎、抗氧化、抗肿瘤等多种药理学活性,广泛用于心血管疾病、糖尿病和癌症等多种疾病治疗[94-97]。近年越来越多的研究发现葛根素具有调节肠道菌群作用。葛根素显著抑制毛螺菌科和脱硫弧菌科的相对丰度,进而调节肠道菌群释放的SCFAs 和修复肠黏膜完整性,改善肠道菌群失衡,从而改善大鼠骨质疏松症[98]。肠道菌群与肠炎发生密切相关,葛根素处理TNBS 诱导的结肠炎模型,改变了共生微生物群结构,降低拟杆菌相对丰度,提高阿克曼氏菌比例,显著增加吲哚-3-丙酸含量,增强杯状细胞增殖及黏液分泌改善肠道屏障功能[99]。相似研究也发现,梭状芽孢杆菌属中瘤胃球菌1 和瘤胃球菌UCG-009 在UC 大鼠中丰度较高,随着葛根素摄入,两个属均恢复正常水平;UC大鼠补充葛根素改变了利用黏蛋白的细菌组成,增加SCFAs 产生,从而影响黏膜层渗透性及修复,缓解肠炎[100]。葛根素通过调节肠道菌群及其代谢产物SCFA 促进肠道黏膜屏障修复,但缺乏临床应用研究。
4.6 花青素 花青素是存在于水果和蔬菜中的天然水溶性多酚,因其具有抑制氧化应激作用而广泛用于预防氧化应激相关疾病,如心血管、神经退行性和炎症性疾病。研究表明,花青素也具有调节肠道微生物群作用,可促进有益菌如乳酸杆菌和双歧杆菌生长[101]。花青素还可与淀粉、SCFA 相互作用间接调节肠道微生物群。而肠道微生物群又可分解花青素产生SCFA 和酚酸,刺激益生菌增殖,增加肠道益生菌数量[102]。另有研究发现花青素能够通过抑制消化酶如α-淀粉酶影响淀粉消化。难消化的淀粉进入大肠,可作为乳酸杆菌、双歧杆菌等多种益生菌能量来源,对人体健康有益[103]。临床研究也发现富含花青素的红葡萄酒和脱醇红葡萄酒可增加双歧杆菌等有益菌[104]。虽然花青素在IBD 中的作用已较明确,但分子机制尚不清楚。目前IBD常用治疗药物存在疗效不佳或出现不良反应问题,花青素通过减弱氧化应激和抑制细胞炎症信号通路显示出有效的保护和治疗功能。轻度至中度UC患者以160 g/d 剂量服用富含花青素的越橘(含840 mg花青素/d),持续6周,发现摄入越橘1周后临床疾病活动指数显著下降,6 周后,UC 患者结肠活检标本中IFN-γ 和IFN-γ R2 表达降低,约50%患者观察到抗炎细胞因子IL-10 升高[105]。KAMALI 等[106]研究了使用富含花青素的石榴皮作为UC 常规药物治疗的补充疗法,一项随机双盲人体试验中,29 例患者每天接受6 g 果皮提取物,持续4 周,安慰剂组33 例患者接受糖浆,与安慰剂组相比,接受果皮提取物的患者结肠炎活动指数(大便失禁、一般健康状况等)得到改善。花青素在临床前研究中显示出有益效果,可缓解IBD,并降低IBD 患者治疗费用。此外,富含花青素的膳食补充剂与现有药物组合可为IBD患者提供新的治疗选择。
4.7 没食子酸(gallic acid,GA)GA 又称棓酸,是一种天然植物中提取的酚酸类物质,主要存在于五倍子、山茱萸、牡丹、皮掌叶大黄等中药中,具有多种药理学作用,在抗肿瘤及抗炎等方面具有良好效果,对多种人类疾病包括UC 亦具有很好的缓解作用[107-109]。但GA在UC中的作用机制尤其是GA对肠道菌群的调控作用知之甚少。研究发现,GA 可调节DSS诱导的UC大鼠肠道微生物组成,促进益生菌如乳杆菌科和普雷沃氏菌科细菌增加,并减少以厚壁菌门和变形杆菌门为主的病原菌产生。粪便代谢物是宿主和微生物群相互作用的副产物,体内代谢物变化可作为微生物群改变的证据。因此,研究者进一步通过代谢组学分析揭示了GA 诱导的代谢物变化主要表现为增加碳水化合物代谢和胆汁酸代谢及降低氨基酸代谢[110]。该研究从肠道菌群角度提出了GA 缓解UC 的新机制。仍缺乏GA 及其衍生物对人类肠道菌群调控的研究,且GA 在人类胃肠道健康和疾病中的有效性临床证据非常有限,潜在作用机制仍不清楚。结合宏基因组学及代谢组学分析人类粪便、尿液和血液中的代谢物水平可深入了解GA对人类IBD的影响。
5 展望
随着世界范围内IBD 发病率上升以及现有IBD临床治疗药物的副作用和局限性,急需新的有效治疗方法。多酚类化合物具有抗炎、抗肿瘤、抗氧化、调节机体免疫力等重要功能,与传统治疗药物相比,多酚化合物在治疗IBD 方面更安全,副作用更小,有望成为治疗肠炎的有效药物,因此,大量研究集中于多酚化合物对肠炎发生发展的影响。肠道菌群是人类健康的关键调节剂,可精细调节肠道微生态平衡,影响宿主代谢功能维持、免疫系统发育以及对肠道病原体入侵的抵抗力。肠道微生态失调与IBD发生发展密切相关。现有证据表明多酚化合物通过与肠道菌群相互作用调节肠道微生态。一方面,多酚化合物调节肠道代谢产物影响细菌生长及肠道菌群丰度。另一方面,肠道微生物可分解多酚产生代谢产物,进而调节微生物群落。虽然人类和动物研究表明多酚化合物可选择性增强特定的肠道微生物,但这些调节作用的机制尚不清楚。未来需要通过代谢组学、转录组学和蛋白质组学方法进行微生物功能分析,进一步描述多酚-肠道微生物群的相互作用。此外,多酚化合物对肠炎缓解作用的研究多在动物模型上开展,临床试验较少,后期还需进一步评估多酚化合物的临床疗效以及患者使用的最佳剂量。虽然多酚化合物在治疗IBD方面有很大潜力,但多酚类化合物稳定性差,溶解度低,生物利用度低,还需深入研究其安全性、剂量以及吸收利用率等,提高多酚类化合物生物利用度,充分发挥多酚化合物治疗肠炎的优势和特色,开发新的IBD防治途径。
猜你喜欢
中老年保健(2022年2期)2022-08-24
中老年保健(2022年5期)2022-08-24
云南化工(2021年11期)2022-01-12
科学(2020年4期)2020-11-26
动物营养学报(2015年10期)2015-12-01
现代检验医学杂志(2015年4期)2015-02-06
中国药理学通报(2014年2期)2014-05-09
中医研究(2014年6期)2014-03-11
中国中医药现代远程教育(2014年22期)2014-03-01
中国中医药现代远程教育(2014年15期)2014-03-01
