
貂熊线粒体基因组全序列分析及其系统进化
2023-11-25 08:57张士芳刘应竹刘凤春朱世兵
野生动物学报 2023年4期
张士芳,魏 丹,郭 媛,刘应竹,路 平,刘凤春,朱世兵
(1.黑龙江省科学院自然与生态研究所,哈尔滨,150040;2.黑龙江双河国家级自然保护区管理局,塔河,165204)
线粒体基因组是线粒体中的遗传物质,参与蛋白质合成,能自主转录、复制和翻译成蛋白质,具有进化速率快、分子质量小、结构简单和母系遗传等特点,已被广泛应用在物种的起源、进化和系统发育研究中[1-2],对研究物种系统进化和分类地位、物种遗传多样性和群体遗传结构等也都具有十分重要的意义。
貂熊(Gulo gulo)是食肉目(Carnivora)鼬科(Mustelidae)貂熊属(Gulo)下的唯一物种,列入国家一级重点保护野生动物名录。貂熊是环北极型动物,分布同寒温带针叶林分布一致,广泛分布在北美和欧亚大陆北部[3]。貂熊仅在中国东北大兴安岭部分区域和新疆北部阿勒泰地区有分布,种群数量估计为300 只左右[4]。由于貂熊在国内分布面积和数量均较少,属于研究较少的物种,目前只在貂熊的种群[5-6]、生境[7-8]、食性[8]和线粒体[9-10]等方面有研究。而在欧洲和北美,对貂熊的全面研究工作都较为重视,在分布区拥有完善的保护和管理措施。貂熊与其他鼬科动物无论是形态还是习性差异都较大,分类地位也一直存在争议。貂熊和貂属动物虽同属于鼬科,但由于生态和形态差异,长期以来被认为是远亲,分属于不同的亚科[11],而部分形态学和行为学观察[12]及分子研究[13-14]则认为貂熊和貂属(Martes)动物亲缘关系更为密切。本研究对中国的貂熊进行线粒体基因组全序列测定和生物信息学分析,并结合GenBank 中其他21 种鼬科动物线粒体基因组数据,对鼬科部分物种的系统发生关系进行研究,以期为貂熊的分类、进化和系统发育提供基因依据。
1 材料与方法
1.1 材料
貂熊DNA 提取于貂熊血液,样品来自救助于大兴安岭地区饲养在齐齐哈尔龙沙动植物园的1 只雌性个体。采后肢静脉血液5 mL 放置于含抗凝剂的采血管内,冰箱内4 ℃保存。基因组DNA 的提取参考从哺乳动物血液中提取DNA 的方法[15],使用酚-氯仿法提取基因组DNA,经1%琼脂糖凝胶电泳检测。-20 ℃保存备用。
选取GenBank 已公布的日本貂(Martes melampus)(登录号:AB291076)和分布于瑞典的貂熊(登录号:AM711901)线粒体基因组全序列,设计可覆盖貂熊线粒体基因组全序列引物。PCR 扩增产物经2%琼脂糖凝胶电泳回收,纯化后利用ABI 3730xl 测序仪测序。
1.2 序列分析
测序得到的序列通过Chromas 和DNAman(LynnonBiosoft,Canada)分析,人工校正后拼接,最后得到貂熊线粒体基因组全序列,通过与近缘物种线粒体基因组比较及通过ClustalX 和DNAman 软件分析定位蛋白编码基因、tRNA 基因、rRNA 基因和非编码控制区。DNAstar统计序列全长、碱基含量等信息。利用在线软件OGDRAW(http://ogdraw.mpimp-golm.mpg.de/index.shtml)绘制貂熊线粒体基因结构图。使用RNAfold 在线预测网站RNAfold web server(http://rna.tbi.univie.ac.at)对rRNA 二级结构进行预测。
1.3 系统进化分析
在NCBI数据库,通过BLAST相互比对,下载21种相关鼬科动物线粒体基因全序列,结合本研究测定的貂熊线粒体基因全序列,组成22种鼬科动物序列库,以棕熊(Ursus arctos)为外群,序列信息见表1。通过MEGA11采用邻接法(neighbor-joining,NJ)和最大似然法(maximum likelihood,ML)构建系统进化树,其中NJ 法经Bootstrap 法检验,共1 000 次循环,根据置信度选择模型maximum composite likelihood。ML 进化树经Bootstrap 法检验,共1 000 次循环,比对后最优模型为GTR+G+I,进化树模型选择general time reversible model,点位进化率(rates among sites)选择G+I,其他参数设为默认。
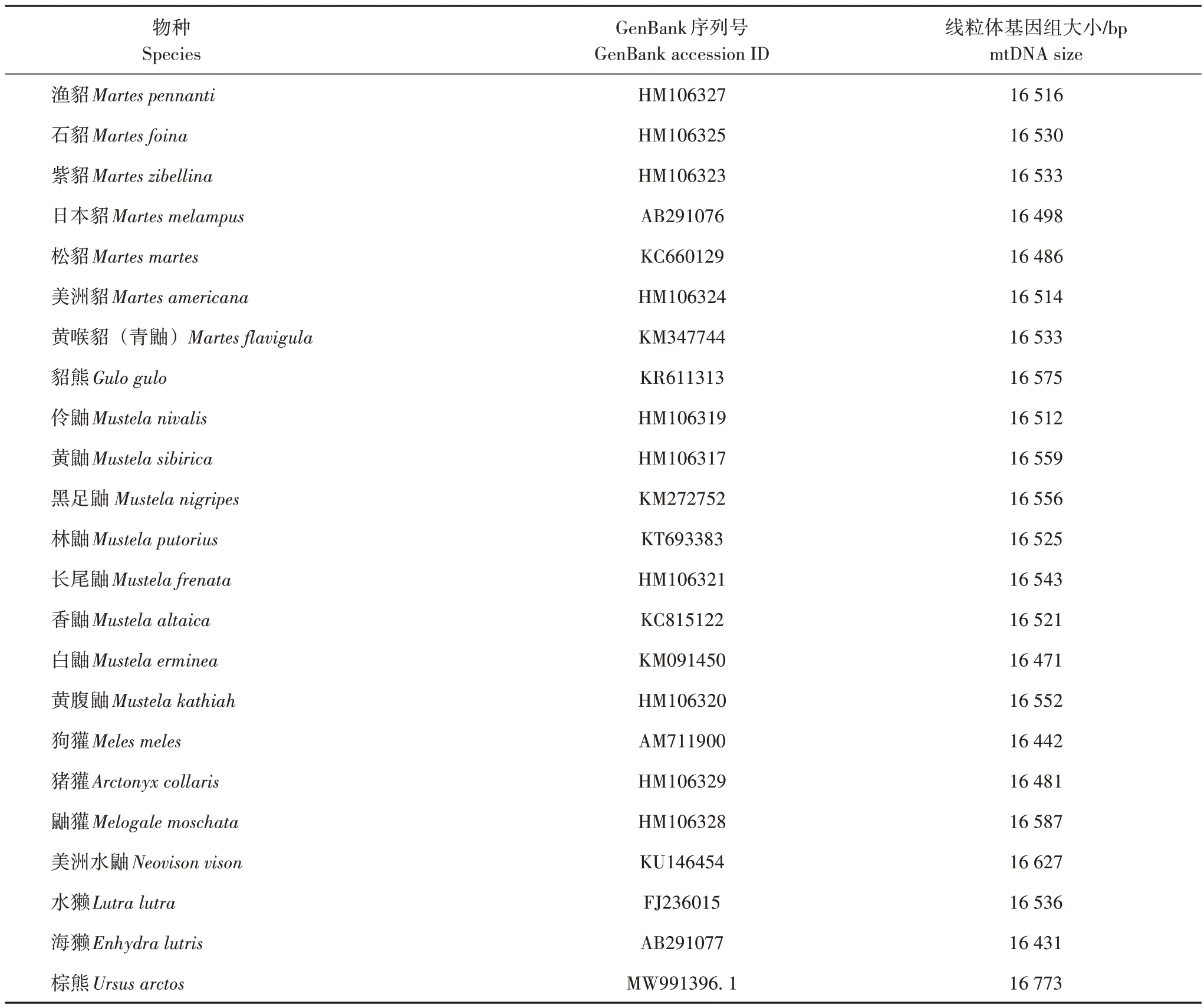
表1 进化分析物种的线粒体全基因组序列信息Tab.1 Information on samples for phylogenetic analysis
2 结果
2.1 基因组组成与基因排列
获得的貂熊线粒体基因组全序列全长为16 575 bp,序列提交NCBI 基因数据库,GenBank 登录号为KR611313。与大多数哺乳动物相似,貂熊的线粒体基因组成包括13种蛋白质编码基因、22种转运RNA(tRNA)、2 种核糖体RNA(rRNA)和1 个非编码控制区(D-loop)。在整个基因组中只有ND61个蛋白质编码基因和tRNA-Gln、tRNA-Ala、tRNA-Asn、tRNA-Cys、tRNA-Tyr、tRNA-Ser、tRNA-Glu和tRNA-Pro8 个tRNA基因位于L链,其余基因均位于H链(图1)。
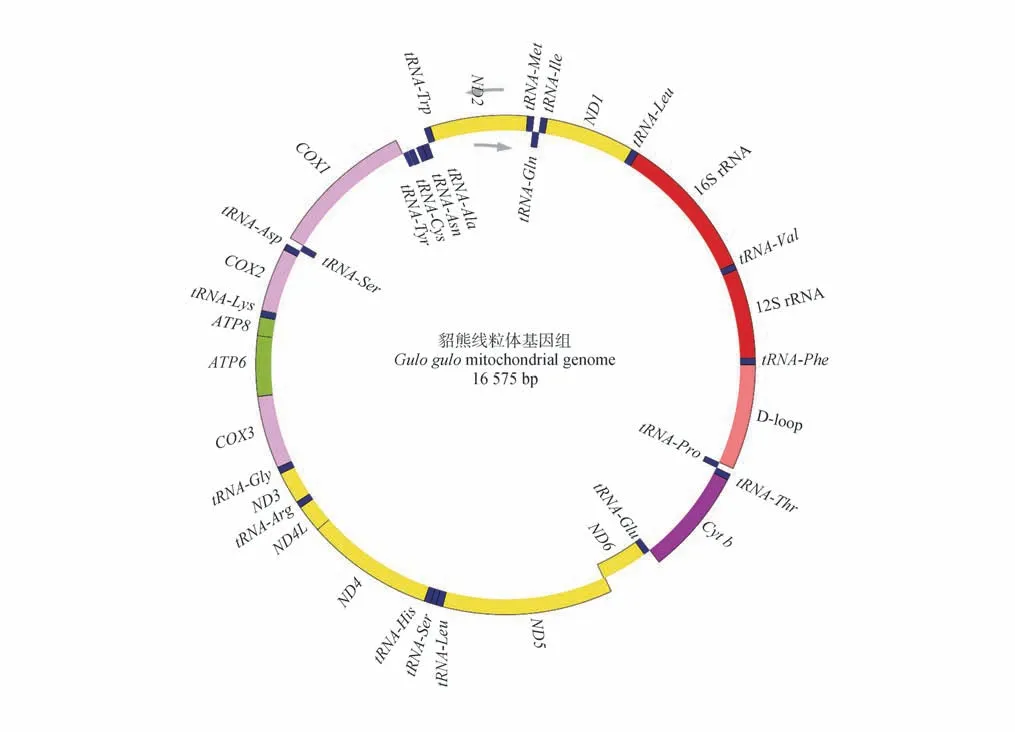
图1 貂熊线粒体结构Fig.1 Mitochondria structure of wolverine
整个貂熊线粒体基因排列十分紧密,在编码区37 个基因之间共有11 个间隔,间隔总长度为33 bp,占DNA 总长的0.20%。7 个重叠区(75 bp)占DNA总长的0.45%,在tRNA-Ile与tRNA-Gln,COX1与tRNA-Ser,ATP8与ATP6,ATP6与COX3,ND4L与ND4,ND5与ND6,以及tRNA-Thr与tRNA-Pro之间,最大重叠为43 bp,位于ATP8与ATP6之间(表2)。在tRNA-Asn与tRNA-Cys之间有1 个长度为33 bp 的茎环结构保守序列Rep Origin,其二级结构如图2 所示,柄部有3个核苷酸,茎部由8对互补的碱基组成,环结构由14个核苷酸组成。
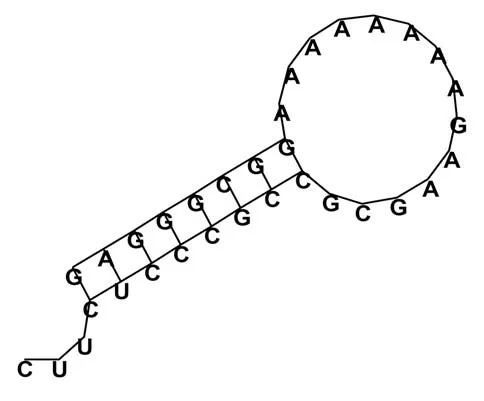
图2 貂熊Rep Origin的二级结构Fig.2 The Rep Origin secondary structure of wolverine

表2 貂熊线粒体基因组组成Tab.2 Gene contents of wolverine mitochondrial genome
2.2 核苷酸组成
由表3 可知,貂熊mtDNA 4 种碱基含量分别为:A=32.18%、G=14.26%、T=26.75%和C=26.81%;AT含量(58.93%)>CG 含量(41.07%),存在显著的AT富集。蛋白质编码基因包括3 782 个密码子,其AT含量为58.58%,与全基因组相似,密码子的碱基组成是第3位点密码子AT含量最高(60.80%),且明显高于D-loop 区AT 含量(55.62%),而密码子第1位点和第2位点的AT含量均低于全序列。

表3 貂熊线粒体基因组核苷酸组成Tab.3 Nucleotide composition of wolverine mitochondrial genome
2.3 蛋白质编码基因
貂熊线粒体基因组同其他脊椎动物相同,有13 个蛋白质编码基因,包括1 个细胞色素b基因(Cyt b),2个ATP酶亚基基因ATP8和ATP6,3个细胞色素氧化酶亚基基因,即COX1、COX2和COX3,以及7 个NADH 脱氢酶亚基基因ND1、ND2、ND3、ND4L、ND4、ND5和ND6。貂熊蛋白质编码基因总长度为11 346 bp,编码3 739 个氨基酸,碱基组成A=31.01%、G=13.16%、T=27.57% 和C=28.27% ;AT 含量(58.58%)>CG 含量(41.42%),与全序列碱基组成相似。在貂熊蛋白质编码基因中,H链上有5个阅读框重叠,COX1和tRNA-Ser有3个重叠,ATP8和ATP6有43 个重叠,ATP6和COX3有1 个重叠,ND4L和ND4有7 个重叠,ND5和ND6有17 个重叠,与紫貂(Martes zibellina)线粒体基因组非常相似[16],仅COX1和tRNA-Ser间的重叠数不同。
在貂熊的13 种蛋白编码基因中,共有4 种起始密码子,ND6以TTA为起始密码子,ND3以ATA为起始密码子,ND2和ND5两个蛋白质基因均是以ATT为起始密码子,其余9 个蛋白质编码基因COX1、COX2、COX3、ATP6、ATP8、ND1、ND4、ND4L和Cyt b均以ATG 为起始密码子,具有明显的多样性。终止密码子方面,貂熊的13个蛋白质编码基因中,6个蛋白质编码基因COX1,COX2、ATP8、ATP6、ND4L和ND5以TAA 为终止密码子,Cyt b以TGA 为终止密码子,这7 种蛋白质编码基因具有完全终止密码子,其余6 个蛋白质编码基因ND1、ND2、COX3、ND3、ND4和ND6均以不完全终止密码子T 为终止密码子(表2)。不完全终止密码子T 的使用频率也比较高。在已知的鼬科动物中,日本貂出现不完全终止密码子4 次,紫貂有7 次,而貂熊有 6 次,使用较频繁。在其他貂属动物线粒体DNA中,使用T或TA的不完全终止密码子的现象也十分常见[17],其3'端通常与tRNA紧密相连,甚至发生重叠,通过在3'端添加Poly A 来完成转录过程。
2.4 tRNA
tRNA 即转运RNA(transfer RNA),貂熊线粒体基因组包括22 个tRNA 基因(表2),长度为61~75 bp,排列顺序与其他鼬科动物相同,其中tRNAGln、tRNA-Ala、tRNA-Asn、tRNA-Cys、tRNA-Tyr、tRNASer、tRNA-Glu和tRNA-Pro由L 链编码,其余14 种tRNA均由H链编码。
对貂熊22 个tRNA 基因的二级结构进行预测,结果显示除了tRNA-Lys二氢尿嘧啶环缺失外,其余21 个tRNA 基因均为标准的三叶草二级结构。部分tRNA 基因的二级结构见图3。在22 个tRNA 基因二级结构中出现了35 处碱基错配的情况,其中U—G错配最多,有25 处,U—U 错配5 处,A—A 错配3 处,以及A—C 错配2 处。通常认为这些错配符合摆动配对原则,有利于维持tRNA二级结构稳定[18]。
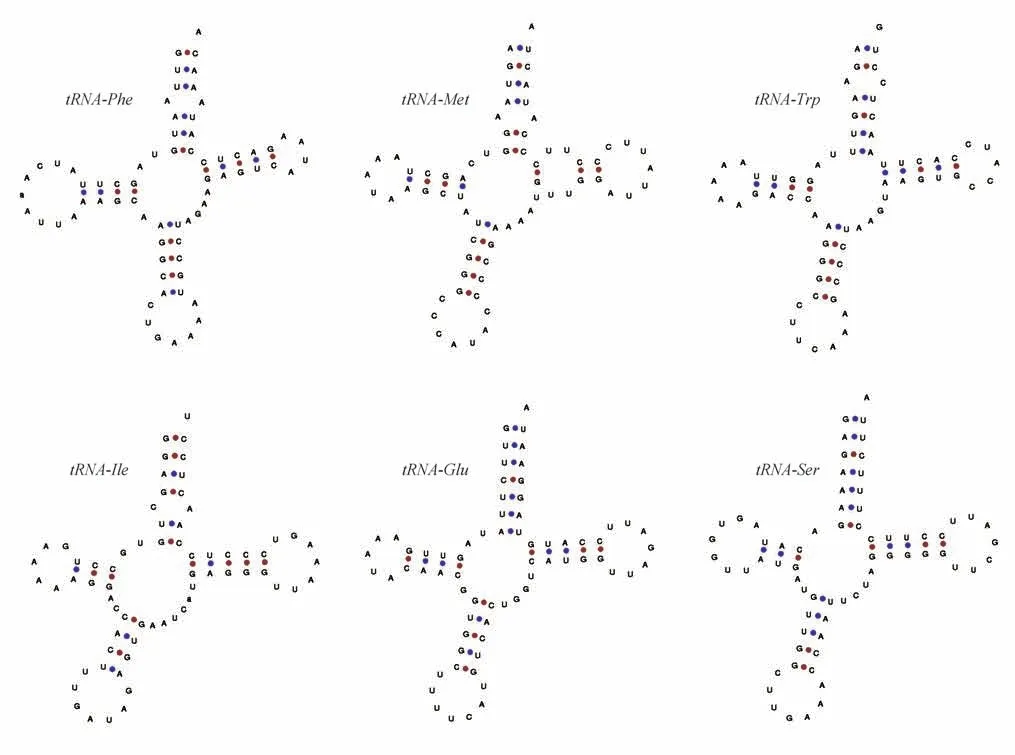
图3 貂熊部分tRNA二级结构Fig.3 Secondary structure of some tRNA of wolverine
2.5 rRNA
貂熊核糖体RNA 基因即rRNA 基因(12S rRNA和16S rRNA)长度分别为961、1 569 bp,分别位于tRNA-Phe和tRNA-Val之间,tRNA-Val与tRNA-Leu之间,2 个rRNA 基因间由tRNA-Val相隔。2个rRNA 的AT 含量分别为59.00%和59.85%,均略高于全序列含量。利用RNAfold 软件预测12S rRNA 和16S rRNA 基因二级结构,结果显示,rRNA 的二级结构比较复杂,形成多个复杂的茎环结构(图4)。
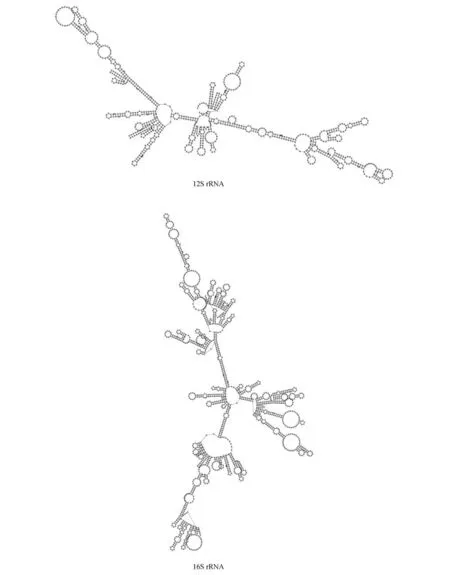
图4 貂熊rRNA二级结构Fig.4 rRNA secondary structure of wolverine
2.6 D-loop控制区
D-loop 区是线粒体DNA 上的主要非编码区,受到的进化压力较小,有较高的变异性,在整个线粒体基因组中属于进化速度较快的区域[19-20]。貂熊的控制区序列长1 129 bp,位于tRNA-Phe和tRNA-Pro之间。碱基组成A=30.12%、G=14.35%、T=25.51%、C=30.03%;AT含量(55.62%)>CG含量(44.38%)。
2.7 系统进化
在NCBI数据库中通过BLAST比对,下载21种相关鼬科动物线粒体基因全序列组成基因组全序列数据,以棕熊为外群物种,分别采用邻接法(NJ法)和最大似然法(ML法)构建鼬科22种动物的进化树(图5,图6)。结果显示,2种方法构建的系统进化树的拓扑结构基本相似,且各个节点都具有较高支持率。外群棕熊单独一支,位于最底部,22种鼬科动物分为5个大支,鼬属(Mustela)的黑足鼬(M.nigripes)、林鼬(M.putorius)、黄鼬(M.sibirica)、香鼬(M.altaica)、伶鼬(M.nivalis)、白鼬(M.erminea)和黄腹鼬(M.kathiah)聚为一支;鼬属的长尾鼬(M.frenata)同美洲水鼬属(Neovison)的美洲水鼬(N.vison)聚为一支;水獭亚科(Lutrinae)的水獭(Lutra lutra)和海獭(Enhydra lutris)则构成另一分支;狗獾属(Meles)的狗獾(M.meles)和猪獾属(Arctonyx)的猪獾(A.collaris)聚为一支;紫貂和松貂(Martes martes)先聚为一支,后依次与日本貂、美洲貂(M.americana)、石貂(M.foina)、黄喉貂(M.flavignla)、貂熊和渔貂(M.pennanti)聚合形成一个大的分支。2种方法测得的结果均显示,貂熊同渔貂和黄喉貂的关系较近。这同Marmi等[21]基于线粒体Cyt b基因数据对貂类系统发育进行研究时所获结果相似。张洪海等[22]通过对线粒体DNA控制区全序列数据的分析也确定了鼬亚科(Mustelinae)和貂熊属的分类地位;此外,徐纯柱等[16]基于部分线粒体控制区基因序列构建的貂类进化树,显示貂熊同渔貂和黄喉貂亲缘关系最近。
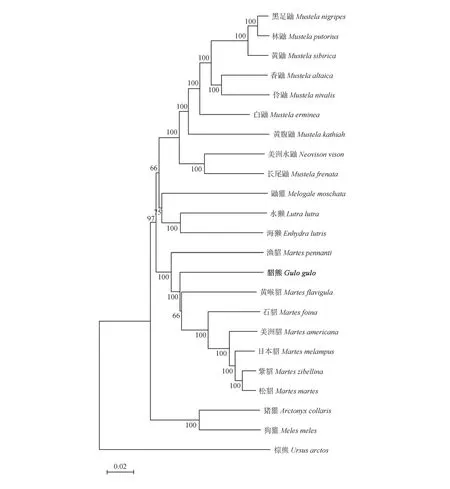
图5 22种鼬科动物NJ进化树Fig.5 NJ evolution tree of 22 species of Mustelidae
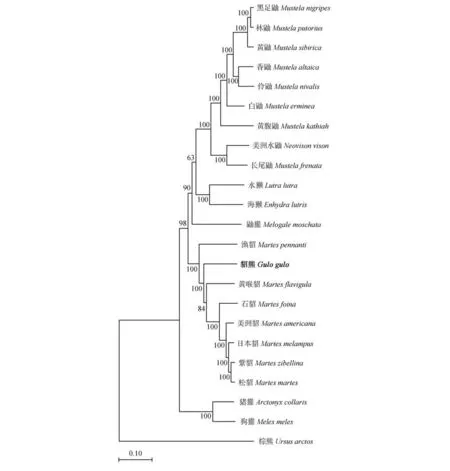
图6 22种鼬科动物ML进化树Fig.6 ML evolution tree of 22 species of Mustelidae
3 讨论
鼬科是食肉目中一个多样性较高的科,各成员之间无论是形态,还是生活习性都有很大差异,多数鼬科动物是动作敏捷的小型食肉动物,也有体型略大的,如貂熊和海獭等,既有陆栖动物,也有水栖或半水栖动物。而貂熊是体型最大的鼬科动物,并且是貂熊属下唯一种,是一种特化的物种,能够捕食大中型动物且有腐食性。貂熊分类地位和亲缘关系的研究具有重要意义,研究人员已从不同方向对其进行了研究。依据形态学研究,Miller[23]认为,虽然貂熊齿形与鼬亚科其他属动物相似,但是它的牙齿十分粗钝,上裂齿特别强大,而小前臼齿在上下颌紧闭时不能接触,加上半蹠行性的足等特点,这些差异足以构成亚科特征。而Ellerman 等[24]认为,貂熊齿形和头骨结构等主要特征与鼬科各属动物相近,认为应该将其归并在鼬科内。目前,后一种观点得到了较多支持。
随着测序技术越来越先进,研究人员可以通过GenBank 简便快速获取大量的分子数据,这对系统发育学研究非常有利,因为更多的分类单元采样可以提高系统发育的准确性[25]。本研究基于mtDNA全序列构建的系统发育树支持貂熊归为鼬科动物的观点。采用NJ 和ML 法构建的鼬科22 种动物的进化树基本一致,表明结果可靠。进化树显示貂熊先同鼬科貂属的7 种动物聚成一支,再与鼬属动物和猪獾、狗獾聚合,说明貂熊同貂属动物进化关系较近。2 种进化树均显示貂熊系统分化节点位于黄喉貂和渔貂之间。这也同近年来分子系统学研究认为貂熊同貂属的关系更近,应该归为貂亚科(Martinae)[26]或者貂熊亚科(Guloninae)[27-30]的结论一致。貂属动物的进化树中,紫貂和松貂先聚为一支,再与日本貂聚合,然后与美洲貂聚合,再同石貂聚合这一拓扑结构,也同形态学研究认为的松貂、紫貂、日本貂和美洲貂的亲缘关系较近,而与石貂的亲缘关系较远[31]相一致。
本研究通过貂熊线粒体基因组全序列对貂熊与鼬科动物的系统进化关系进行了探索,但为了更准确地反映其系统进化关系,后续还需结合动物地理学、形态学及古生物学等方法进行综合分析。
猜你喜欢
天津市教科院学报(2021年5期)2021-11-10
海洋通报(2021年1期)2021-07-23
生物学通报(2021年9期)2021-07-01
生物学通报(2021年4期)2021-03-16
生物学通报(2020年11期)2020-10-22
中成药(2018年7期)2018-08-04
江苏农业科学(2016年8期)2017-02-15
淮阴工学院学报(2014年5期)2014-09-10
癌变·畸变·突变(2014年1期)2014-03-01
茶叶通讯(2014年4期)2014-02-27
