
密码子与反密码子的本质与拓展
2020-10-22 01:20:20金志忠云南省安宁中学云南安宁650300
生物学通报 2020年11期
金志忠 (云南省安宁中学 云南安宁 650300)
遗传信息表达是高中生物学的重点知识,基于科学史阐述密码子与反密码子的本质与特性,旨在帮助学生形成科学思维与生命观念,进而提高教学深度。
1 遗传密码的破译实验
英国科学家克里克以T4噬菌体为实验材料,研究其中某个基因的碱基的增加或减少对其编码的蛋白质的影响,结果证明遗传密码中3 个碱基编码1 个氨基酸,同时表明遗传密码从一个固定的起点开始,以非重叠的方式阅读,编码之间没有分隔符。就在克里克的实验完成的同一年,年轻的尼伦伯格和马太采用蛋白质的体外合成技术破译了第1 个遗传密码,实验示意如图1。
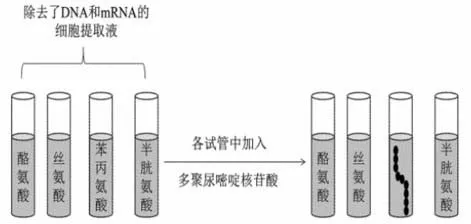
图1 蛋白质体外合成的实验示意图
实验结果表明:加入了苯丙氨酸的试管中出现了多聚苯丙氨酸的肽链。该实验结果说明多聚尿嘧啶核苷酸导致了多聚苯丙氨酸的合成,即尿嘧啶的碱基序列编码由苯丙氨酸组成的肽链,结合克里克的3 个碱基决定1 个氨基酸的实验结论,得出与苯丙氨酸对应的密码子应该是UUU,即苯丙氨酸的密码子为UUU。此后,科学家沿着蛋白质体外合成的思路,不断改进实验方法,破译了全部的密码子。
2 密码子种类
破译了密码子后,科学家将mRNA 上3 个相邻的决定1 个氨基酸的碱基定义为密码子,共有64 种,其中起始密码子2 种,终止密码子3 种。绝大多数生物体内以AUG 作为起始密码子,同时兼作甲硫氨酸的密码子[1]。嗜盐古生菌hsp70 等少数细菌以GUG 作为起始密码子,真核生物偶尔也会以CUG 作为起始密码子。UAA、UAG、UGA 是终止密码子,无编码肽链的能力,起到“句号”的作用[2]。终止密码子也称作无意义密码子,其中UAA 的终止效率最高,UAG 容易被读通,终止效率最低,为保证翻译准确结束,有时成熟的mRNA 末端会以连续2 个甚至3 个终止密码子强化终止。
3 反密码子阅读密码子规律
以转录形成的成熟mRNA 为模板,利用细胞质中各种游离的氨基酸合成具有一定氨基酸顺序的蛋白质的过程称为遗传信息的翻译。该过程需要用tRNA 搬运氨基酸,tRNA 分子比mRNA小很多,分子结构也很特别,经过折叠,像三叶草的叶形,其一端是携带氨基酸的部位,另一端有3 个碱基,这3 个碱基可与mRNA 上的密码子互补配对,称为反密码子。通常情况下,信息的翻译从mRNA 的5′端向3′端进行,习惯性地将5′端写在左侧,3′端写在右侧[3]。查阅各种生物化学、分子生物学等书籍,发现对反密码子阅读密码子的方式的叙述有所差异,目前普遍接受的是按照5′→3′方向进行阅读,即tRNA 的反密码子在核糖体内通过碱基的反向配对与mRNA上的密码子相互作用,反向配对的基本方式如图2所示。

图2 反密码子阅读密码子规律示意图
由图2可知翻译时,反密码子的第1 位碱基与密码子的第3 位碱基互补配对,通常密码子均从5′端→3′端(从左往右)书写,例如,甲硫氨酸的密码子为AUG,即5′端—AUG—3′端,反密码子与mRNA反向配对,阅读也按照5′端→3′端进行,但书写时一般都是以书写密码子的方向写,因此,甲硫氨酸的反密码子写作UAC,第1 位写在右侧。
4 密码子特性
4.1 密码子简并性 能编码氨基酸的61 种密码子决定20 种氨基酸,这必然就会出现1 种氨基酸拥有多个密码子的现象,此现象称为密码子的简并性。除甲硫氨酸和色氨酸只有1 个密码子外,其他18 种氨基酸均至少有2 个密码子,例如,UUA、UUG、CUU、CUC、CUA、CUG 均编码甘氨酸。密码子的简并性可减少因碱基对替换造成基因突变带来的有害突变,即某个基因突变后转录形成新的mRNA 上的密码子与突变前的密码子决定同一种氨基酸,使得突变后的基因仍然翻译出正常的蛋白质,执行正常的生命活动。密码子的简并性主要表现在第3 位碱基上,一般同一种氨基酸的密码子前2 位碱基相同,第3 位碱基不同,只有2 种密码子的氨基酸,密码子的第3 位碱基一般是嘧啶或嘌呤,例如,谷氨酸的密码子为GAA、GAG,第3位碱基均为嘌呤。
4.2 密码子变偶性 在密码子简并性的基础上,Crick 于1966年提出遗传密码的变偶性,他发现反密码子的第1 位碱基与密码子的第3 位碱基的配对可在一定范围内变动,即密码子与反密码子的配对具有摆动性[4]。密码子变偶性主要得益于其第3 位碱基具有较大的灵活性,研究已知,一级结构的tRNA 发现其反密码子的第1 位碱基为I、G、C、U,未出现A,显然I 是由A 转变而来,密码子变偶性具体表现如表1。

表1 密码子与反密码子配对的变偶性
I 属于稀有碱基次黄嘌呤,反密码子中的次黄苷酸能与尿苷酸、胞苷酸或腺苷酸配对,这种配对方式对异亮氨酸的密码子极其重要,已知异亮氨酸的密码子为AUA、AUC、AUU,甲硫氨酸的密码子为AUG。当反密码子的第1 位碱基为U 时,该反密码子识别AUA 的同时也会识别AUG,可能使编码异亮氨酸的密码子被阅读为甲硫氨酸的密码子,导致蛋白质合成过程中氨基酸进位错误。然而次黄苷酸为第1 位的反密码子只识别AUA、AUC、AUU,无法识别AUG,这样,翻译过程中氨基酸进位错误得以一定程度的控制,同时,由于次黄苷酸的多种配对能力,使得异亮氨酸的3 个密码子顺利阅读出来[5]。密码子变偶的重要意义在于当密码子的第3 位碱基发生改变时,仍可能翻译出正确的氨基酸序列,合成的蛋白质具有正常的生物学活性,同时防止氨基酸进位错误,提高遗传信息翻译的效率。
4.3 密码子防错性 由于绝大多数种氨基酸对应多种密码子,因此,当某个密码子的第3 位碱基发生改变时基本不改变其所编码的氨基酸。例如,编码丝氨酸的一个密码子为UCC,若第3 位碱基转换为A,其编码的氨基酸依然是丝氨酸;即使编码的氨基酸改变,也常常由性质相似的氨基酸代替,例如,丝氨酸和苏氨酸的密码子分别是UCN 和ACN(N 代表U、C、A、G 4 种碱基中的任意一种),编码丝氨酸的密码子的第1 位碱基U 突变为A 后,编码的丝氨酸由苏氨酸代替,2 种氨基酸的理化性质很接近,对翻译出的蛋白结构和功能影响较小。例外的是密码子的第3 位转换会引起甲硫氨酸(AUG)和异亮氨酸(AUU、AUA、AUC)间互换,或引起色氨酸(UGG)与终止密码(UGA)间互换。
此外,同义密码子在密码子表中的分布很有规律,密码子的碱基排列顺序与其所编码的氨基酸的理化性质有一定的关系,尤其是第2 位碱基,该碱基决定了氨基酸的极性。例如,当密码子的第2 位碱基为尿嘧啶时,常编码非极性、疏水和支链氨基酸,分布在球蛋白的内部,例如,苯丙氨酸、亮氨酸和缬氨酸等[6]。
由此可见,即使密码子的一个碱基被置换,大多数情况下依可编码出相同的氨基酸,或以理化性质最相近的氨基酸取代,这样将突变带来的危害降至最低,体现了密码子的防错功能。
4.4 密码子“特例” 教材的标准密码子表中,编码甲硫氨酸和色氨酸的密码子各只有1 个,分别是AUG 和UGG,但在果蝇、酿酒酵母等动物线粒体中的基因表达时,AUA 也可编码甲硫氨酸,在支原体中终止密码子UGA 可编码色氨酸。在哺乳动物线粒体基因表达时,原编码精氨酸的密码子AGA、AGG 变成了终止密码子。在部分微生物中编码相同氨基酸的不同密码子被使用的概率也各不相同,例如,在大肠杆菌中,编码苏氨酸的密码子中,ACU 被使用概率约为55.38%,ACC 被使用概率约为40%,ACA 被使用概率约为4.62%,而ACG 完全没有被使用,这种经常被使用的密码子称为偏爱密码子[7]。
近年来科学家发现了一种特殊的氨基酸——硒代半胱氨酸(称为第21 种氨基酸),通常由终止密码子UGA 编码,该密码子可被硒代半胱氨酸—tRNA 识别。硒代半胱氨酸掺入到蛋白质受到mRNA 阅读框内的UGA 密码子和下游链的茎-环(stem-loop)结构控制,这种特殊结构称为硒代半胱氨酸插入序列(SECIS)。在部分生物中,当mRNA链上UGA 密码子的下游出现SECIS 时,UGA 正常编码硒代半胱氨酸,使硒代半胱氨酸掺入到新合成的肽链中。
4.5 密码子进展 科学家一直在探究和发展密码子和反密码子的本质与拓展,目前,已用自然界中不存在的、实验室合成的X—Y 碱基对和相应的氨基酸,成功创造了包含A、T、G、C、X、Y 这6种碱基的全新生命体[8]。这说明人工合成的碱基对在特定环境中也能完成遗传信息的储存、复制和表达,也意味着密码子和反密码子种类和作用机理在不断被拓展和更新。
5 副密码子结构与功能
tRNA 通过反密码子与mRNA 上的密码子相互识别配对,将特定的氨基酸运送到核糖体的肽链合成位点上,但tRNA 与特定氨基酸结合不完全取决于反密码子,侯雅明等[9]将丙氨酸抑制型tRNA 突变很多位点,结果发现许多突变体均不改变携带丙氨酸的性质,唯独改变“5′G 端—G3·U70—3′端”中的“G3·U70”这一碱基对才表现明显的突变效应,tRNA 上类似“G3·U70”这样的小元件称为副密码子,即tRNA 分子上决定其携带氨基酸的区域。副密码子并没有固定的位置,大多集中在tRNA 的接受臂或反密码子环上,副密码子也可能并不止一个碱基对,例如,tRNACys/CUA 和tRNAPhe/CUA都具有“C3·G70”,显然其副密码子不在或不完全在“C3·G70”上,因为其携带2 种不同的氨基酸。副密码子在一定程度上能使tRNA 中的反密码子发生突变后继续将相应的氨基酸携带至核糖体的相应位点,以确保翻译正常进行。tRNA 分子起源于能携带氨基酸的寡聚核苷酸,这种识别氨基酸的特异性在某种程度上被保留下来,而反密码子是在tRNA 分子进化过程中后来才慢慢出现的,这在某种程度上说明副密码子比密码子和反密码子更为古老、更为原始。
6 无义抑制突变特性
一般情况下,产生终止密码子而使翻译提前终止的突变称为无义突变。无义突变中产生UAA 的称为赫石型,产生UAG 的称为琥珀型,产生UGA 的称为乳石型[9]。无义抑制突变是指抑制无义突变表现的突变,抑制突变通常由编码tRNA 的基因引起,该基因突变后形成了一种可携带氨基酸的tRNA 分子(称为抑制型tRNA),该种tRNA 分子可识别终止密码子,使翻译通过无义突变而继续进行,例如,无义突变使“5′—AUG…UUG…UAA—3′”突变为“5′—AUG…UAG…UAA—3′”,而此时tRNATyr/AUG发生抑制突变形成tRNATyr/AUC,顺利阅读UAG密码子,直到遇见下一个终止密码。若上述抑制型tRNA 携带的氨基酸与野生型不同,则会使蛋白质的结构和功能发生改变,影响生命活动。
无义抑制突变在不同进化地位的生物中既有一定的保守性,也有一些差异,高等真核生物中无义抑制突变相对普遍,但在原生生物中尚无机制层面的研究[10],抑制型tRNA 在一定程度上可很好地矫正无义突变和错义突变,这为生物多样性的发展和稳态奠定了良好的基础。
7 小结
密码子和反密码子是将DNA 中的遗传信息传递到蛋白质的桥梁,深度剖析密码子与反密码子的本质和作用机理,能使学生准确理解遗传信息的表达过程,同时深刻感受科学技术不断进步带来的新知识的魅力,也能客观、理性评价生物界的基因多样性、物种多样性和生态系统多样性,充分领略生物在不断进化,知识在不断更新和发展。也许将来科学家会利用更精准的技术创造出更多的人工碱基对添加到遗传密码中,使某些非天然氨基酸被编码到蛋白质中,发挥独特的功能,为医疗、养生等大健康领域研究提供广阔的前景。
猜你喜欢
青年文学家(2024年10期)2024-05-26 18:27:59
食品科学(2023年13期)2023-08-12 00:41:48
北京大学学报(医学版)(2022年2期)2022-11-21 03:29:12
中国生物化学与分子生物学报(2022年7期)2022-09-07 05:28:48
中国药科大学学报(2021年6期)2021-12-31 03:05:20
世界最新医学信息文摘(2021年24期)2021-04-29 03:47:14
遗传(2019年5期)2019-05-21 09:58:28
岷峨诗稿(2018年2期)2018-11-14 18:51:09
分析测试学报(2015年4期)2016-01-13 06:18:25
分析测试学报(2015年9期)2015-12-17 16:44:28
