
基于PAE编码系统笼养黑叶猴的行为谱
2023-11-25 08:56何晓露赵秋程冯月婷李毅峰黄中豪李友邦
野生动物学报 2023年4期
何晓露,赵秋程,冯月婷,李毅峰,黄中豪,李友邦
(1.珍稀濒危动植物生态与环境保护教育部重点实验室,广西师范大学,桂林,541006;2.广西珍稀濒危动物生态学重点实验室,广西师范大学,桂林,541006;3.梧州市园林动植物研究所,梧州,543000)
行为的多样化是动物适应复杂多变的自然环境的外在表现,是动物行为学研究的基础[1]。根据动物行为的生态功能,在辨识和分类动物的行为后建立的行为目录,即为该动物行为谱(ethogram)[2]。为了对不同的行为谱进行数理方法的比较,依据动物行为发生的环境、采用的姿势和相应的动作(姿势-动作-环境,posture-act-environment,PAE)将其进一步分解成不同的行为层次和基本单元,构建可量化分析的PAE 行为编码分类系统[3]。PAE 编码系统的构建,有助于对动物行为的功能、行为与环境的相互关系的理解[1-2,4]。
非人灵长类动物的行为生态学研究得到了广泛关注[5-7]。目前,在非人灵长类动物行为谱的构建中,PAE 行为谱编码方式已得到较多应用,在野外状态下,主要有猕猴属(Macaca)[8-10]、仰鼻猴属(Rhinopithecus)[11-12]和叶猴属(Presbytis)[13]的研究;在圈养或半放养状态下,有狐猴属(Lemur)[14]、狨属(Callithrix)[15]和狒狒属(Papio)[16]等的研究,研究结果表明,非人灵长类所属的分类阶元关系越近,越易形成基本相同的典型行为,同时各分类阶元又有其特有的行为[9]。
黑叶猴(Trachypithecus francoisi)是越南和我国贵州、广西及重庆典型石灰岩地区的特有种[17]。现今国内外对黑叶猴行为生态学已有一定的研究,包括日活动时间分配[18-19]、玩耍行为[20]、栖息地利用行为[21-22]、觅食行为[23-25]、夜宿和漫游行为[26-28]、位置行为[29]及理毛行为[30]等。这些研究主要集中在黑叶猴的某个动作模式上,而在特定状态下系统描述黑叶猴表现出的所有行为模式[31]的研究较少。本研究采用PAE行为编码分类系统建立笼养黑叶猴的行为谱,为深入开展黑叶猴野生种群的行为生态学研究奠定基础。此外,圈养珍稀野生动物由于经常缺乏表达空间,行为多样性会减少,通过行为谱的建立,研究其刚性行为与弹性行为构成,对保护和救助的野生动物的野化放归也具有重要意义。
1 材料与方法
1.1 研究地点
广西梧州黑叶猴珍稀动物繁殖中心位于广西壮族自治区梧州市云龙公园内,三面环山,人为干扰强度低,是目前世界上规模最大的黑叶猴繁殖基地,拥有最完整的黑叶猴谱系档案[32]。研究区域属亚热带气候,年平均气温22 ℃,相对湿度76%~84%,旱雨季分明。
笼养黑叶猴的笼舍为笼网式。笼舍分为内、外两室,均为水泥地面。内室约为4 m×2 m×3 m,较暗且人为干扰小,有水泥平台和保温板,是黑叶猴躲避降雨、暴晒及夜间睡眠的场所。外室约为4 m×4 m×3 m,有树桩、轮胎和铁索等丰容设施,供猴群玩耍和休息。
1.2 研究对象
选择笼养的3 个繁殖群A3、B1 和B2 共17 只黑叶猴为研究对象,依据笼养黑叶猴年龄组的划分标准[31,33],将17 只黑叶猴的年龄组成划分为成年雄性猴(3 只)、成年雌性猴(8 只)、亚成年猴(1 只)、少年猴(2 只)和婴幼猴(3 只)。选取个体的亲缘关系及观察时段见表1。
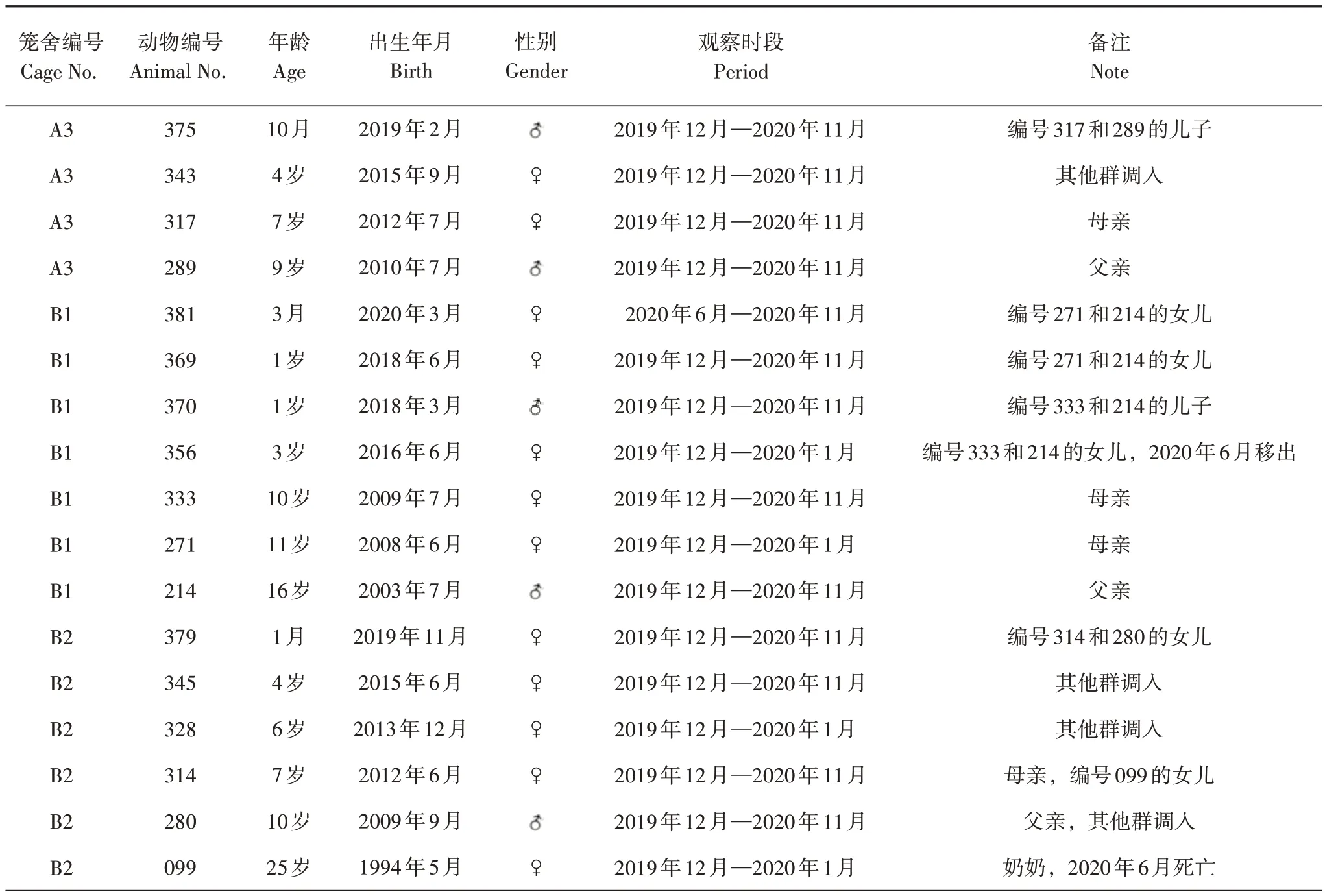
表1 笼养黑叶猴个体情况及观察时段Tab.1 Specific conditions and observation period for captive François’ langurs
1.3 研究方法
1.3.1 环境及行为定义
环境:将研究地点的环境条件划分为生物环境和非生物环境。生物环境指环境因素中其他有生命的生物。非生物环境是生态系统中由物理、化学或其他非生命物质组成的非生物因子的总称[34]。
姿势:动物身体的部分结构在一定时间内保持一定的位置和形态[35]。
动作:在较短的时间内,动物机体的肌群发生运动,使得机体的部分结构位移、舒张、收缩、弯曲和运动[35]。
1.3.2 预观察
对17 只黑叶猴进行1 个星期的预观察,初步掌握黑叶猴的活动规律和行为模式,同时让观察者和17 只黑叶猴彼此熟悉,以便17 只黑叶猴能真实反映在笼养状态下的行为特征。
1.3.3 行为取样方法
行为取样时间为2019 年12 月—2020 年11 月,由于新冠疫情原因,2020 年2—5 月停止观察,其余月份每月连续取样6 d。根据预观察结果,夏季观察时间为07:00—18:30,其他季节观察时间为07:30—18:00。采用瞬时扫描取样法(instantaneous scan sampling)和随机取样法(ad-libitum sampling)[36]对黑叶猴观察取样,参照Hunt 等[37]定义的行为模式,观察并记录笼养黑叶猴的全部行为,记录行为的姿势、动作和环境,参照非人灵长类动物PAE 行为谱的相关行为术语[7,9-11],赋予相应代码,同时记录个体的性别和年龄。从每个笼中猴群的最左侧开始扫描,依次记录个体表现的行为类型,每次扫描持续5 min,间隔10 min 再次取样,以保证各样本之间的独立性。
1.4 数据处理
将个体的每次行为扫描视为1 个独立样本。首先,按照不同性别年龄组的行为发生情况,计算每天各行为发生的频次比例,再将该月每天各行为发生的频次比例平均,得到每月各行为发生的频次比例。行为发生频次由低到高表示为:+(0~5%),++(>5%~10%),+++(>10%)[9];同理,计算不同繁殖群在研究期间的行为发生频次比例。采用Kruskal-Wallis 检验各行为在不同繁殖群之间的差异。差异检验的显著性水平为0.05。
2 结果
2.1 笼养黑叶猴姿势编码
通过观察、辨认,共记录到黑叶猴16 种姿势,前7种是静止姿势,走、跑、跳、臂摆荡、攀爬和爬跨为运动姿势,抱、携挂和哺乳多为成年雌性猴对未成年婴幼猴的照料等姿势(表2)。

表2 笼养黑叶猴姿势编码Tab.2 Posture codes for captive François’ langur
2.2 笼养黑叶猴动作编码
依次从头颈部、嘴部、眼耳鼻部、四肢和腰荐腹部对笼养黑叶猴的动作进行分辨,共记录到98 种动作,并对其编码(表3)。结果显示,黑叶猴的四肢动作最多,为39 种,其次是嘴部动作,为23 种,头颈部和眼耳鼻部动作最少,各为9种。

表3 笼养黑叶猴动作编码Tab.3 Act codes for captive François’ langur
2.3 笼养黑叶猴行为发生的环境编码
笼养黑叶猴行为发生的环境共有19 种,包括8 种生物环境和11 种非生物环境,行为发生的环境较简单(表4)。

表4 笼养黑叶猴行为发生的环境编码Tab.4 Codes of behaviors and associated environment for captive François’ langur
2.4 笼养黑叶猴PAE行为谱
研究期间共记录到笼养黑叶猴87 种行为,按照行为的生理和生态功能[9],将87种行为划分为摄食、排遗、调温、亲密、竞争、聚群、通讯、休息、运动、繁殖、异常和其他共12 类(表5,表6),各行为类型在3 个繁殖群间发生的比例相近,且在3个群间无显著差异(表5)。这些行为类型又可归为生存、社会、繁殖和稀有行为4类。
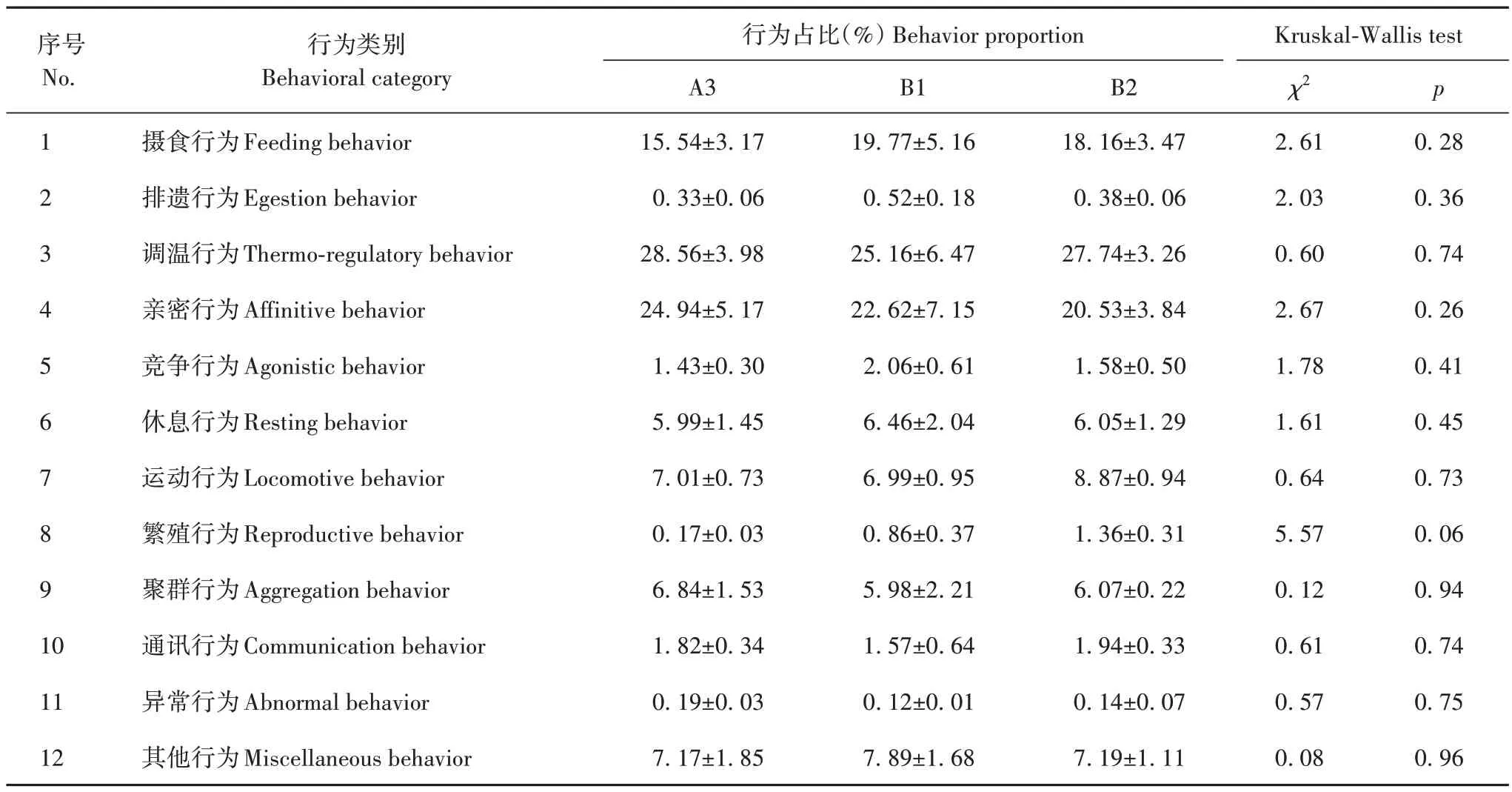
表5 不同行为类型在各繁殖群中发生的比例Tab.5 Proportions for each behavior at different François’ langurs breeding groups in captivity
2.4.1 生存行为
摄食行为(feeding behavior):黑叶猴个体在笼养环境中采食所能获得的食物资源(如植物的嫩叶、嫩芽等部位)、捡拾食物、饮水和婴幼个体吸乳等行为。
排遗行为(egestion behavior):黑叶猴个体在食物消化后或遇到紧急情况时的排尿、排粪等行为。
调温行为(thermo-regulatory behavior):黑叶猴个体为维持稳定的体温而做出的适应外界环境温度的一系列行为,包括树干静息、内室静息等行为。
2.4.2 社会行为
亲密行为(affinitive behavior):黑叶猴用来确定或维持个体之间亲和关系的行为,如挨坐、互相理毛和玩耍等行为。
竞争行为(agonistic behavior):黑叶猴个体之间由于冲突所发生的一系列撕咬、抓打和驱赶等行为。
聚群行为(aggregation behavior):多个黑叶猴个体通过特定的形式聚集,同时表现出相互联系和相互影响的行为。
通讯行为(communication behavior):黑叶猴个体之间在家庭单元内或家庭单元外通过特定的动作、姿势和声音等传递信息的行为。
休息行为(resting behavior):黑叶猴个体在合适的环境中保持一定的姿势,机体呈现放松的状态。
运动行为(locomotive behavior):黑叶猴个体通过活动四肢来完成身体位移的行为,如跳跃、奔跑和行走等行为。
2.4.3 繁殖行为
繁殖行为(reproductive behavior):黑叶猴个体为繁衍后代进行的一系列生理活动的行为,如产仔和哺乳等行为。
2.4.4 稀有行为
异常行为(abnormal behavior):与在自然环境下相比,黑叶猴个体在人造环境中做出的不正常行为,包括晃动头和摇晃身体2种行为。
其他行为(miscellaneous behavior):黑叶猴个体之间为寻求舒适而发生的一些频次较低的行为,如挠痒、自我清洁等行为。
3 讨论
PAE 行为分类编码系统的研究已在部分非人灵长类物种中开展,包括黔金丝猴(Rhinopithecus brelichi)[11]、川金丝猴(Rh.roxellana)[12]、藏酋猴(Macaca thibetana)[38]、褐狐猴(Lemur fulvus)[14]和普通狨猴(Callithrix jacchus jacchus)[15]等。这些研究表明非人灵长类的行为发生频次与年龄、性别等有关,不同物种均具有社会行为、生存行为、个体行为及繁殖行为等典型行为,但在分类的细节上,每个物种又具有特有的行为,这可能是动物在特定的环境条件下形成的特殊行为,相同的动作模式也因长时间适应环境和后天学习而产生了特有的行为表达[12],如岷山藏酋猴有游泳、溢精等行为[38]。本研究中笼养黑叶猴的PAE行为谱与上述非人灵长类动物相比,具有一些特有行为,如蹭痒、摇晃身体等行为,因此,有必要构建不同物种的行为数据库,通过PAE行为编码分类系统构建PAE行为谱,可实现种间行为数据的比较。
有研究显示,动物的行为表达受活动空间大小影响,当生存空间受限时,刚性行为(如休息、排遗和摄食行为等)仍将表现出来,但弹性行为(如亲密、通讯和玩耍行为等)的表达有较大差异[6]。本研究中笼养黑叶猴个体均表现出的刚性行为包括摄食行为、排遗行为、调温行为、休息行为和运动行为,表现出的较大的弹性行为包括聚群行为、通讯行为、亲密行为、繁殖行为、竞争行为、异常行为和其他行为。本研究3 个繁殖群体的个体数量、性比和年龄结构不同(表1),但3 个繁殖群体的弹性行为出现比例无显著差异(p>0.05),A3 繁殖群的繁殖行为比例比B1、B2 繁殖群低约4.23、2.62 个百分点(表5),可能是研究期间A3 繁殖群能够繁殖的只有1 对成年雌雄个体,其余个体未能参与到繁殖行为中,社会结构较简单。笼养黑叶猴表现出较大的弹性行为,可能是该中心黑叶猴繁殖单元的个体数量和性比合理,笼舍面积大小合适,黑叶猴有足够的空间活动,个体之间的社会交往行为能够充分表达[3],也可能是笼舍有足够的丰容设施,如树桩、轮胎和铁链等可供黑叶猴玩耍[39]。此外,笼养黑叶猴的饲料丰富,有新鲜的嫩树叶、坚果和糕料等,丰富的丰容设施和食物使笼养黑叶猴更多地保持和表现自然行为[40]。
野外黑叶猴的行为划分为觅食、休息、移动、饮水、出洞、入洞、舔食石壁、社会行为和晒太阳9 大类[31],根据其生理生态目的,前7 类为刚性行为,后2 类为弹性行为[6]。目前未见关于野外黑叶猴系统的PAE 行为谱研究,且野外黑叶猴行为研究的定义[27,31]与本研究也有差异,但一些行为仍具一定的可比性。笼养黑叶猴个体从内室到外室的行为与野外出入洞行为相似;晒太阳行为(调温行为)是野外黑叶猴在低温条件下为保持体温而表现的行为[41-42],与秋冬两季(室内外温度相差约10 ℃)笼养个体进入内室的调温板取暖行为相似;舔食石壁是野外黑叶猴为获取生存所需的矿质元素[25]所表现的行为,与笼养个体表现出的舔网、舔墙壁和舔水泥地面等行为相似。野外黑叶猴的社会行为包括玩耍、理毛和拥坐,反映出个体的序位和亲缘关系,同时具有清洁和调温等作用。笼养黑叶猴的社会行为(表6)与野外黑叶猴相似,如亲密行为和玩耍、理毛行为相似,聚群行为和拥坐行为相似。野外黑叶猴行为划分中并没有明显的竞争行为,可能是野生环境中,黑叶猴群拥有一定的家域面积,不同群体间的相遇机会少,使群体间的竞争行为较少被观察到[28]及群体内的等级序位降低了群内竞争行为的发生[30]所导致。野外黑叶猴通讯行为较少被观察到的原因可能是群体移动或休息时个体间的距离较近,且通讯行为多发生在不同猴群遭遇或发现天敌时,此行为常被归入预警通讯行为中,而未明确划分到野外黑叶猴的行为类型中[43]。由此可见,在弹性行为与刚性行为的表现上,笼养个体与野外个体又存在一定差异。

有研究表明,采用相同PAE行为定义,野生个体较同种笼养个体表现的行为多样性更高,如利用红外相机观察野生川金丝猴,共统计到17 种姿势、84 种动作和116 种行为,包括摄食、调温、休息和运动等刚性较大的行为,以及交配、母性和幼儿、亲密、作威、威胁、攻击、屈服、聚群和其他等弹性较大的行为[12];而笼养条件下共统计到20 种姿势、76 种动作和49 种行为,包括取食、排遗、休息和运动等刚性较大的行为,以及交配、育幼、梳理、拥抱、游戏、睡眠和其他等弹性较大的行为[44]。这可能是野外空间大,野生川金丝猴个体有足够的空间表达各种各样的行为[12],因此,即使不能获得笼养条件下所观察到的一些行为(如分娩、通讯和排遗行为等),野外个体统计的动作和行为仍多于笼养个体。但笼养个体的姿势多于野生个体,可能是前者对圈养环境的适应产生了一些后者未统计到的姿势。本研究中的笼养黑叶猴与野外环境的远缘非人灵长类物种[12,44]相比,发现姿势数量相差不大,但动作更多,行为少,这可能是种间差异[7,9-11,36],也可能是生存环境不同使行为表达的强度不同。若按照刚性行为与弹性行为来比较,本研究中的黑叶猴和其他灵长类物种的研究结果[9]相似。
值得注意的是,除了完整地表达刚性行为和较丰富的弹性行为外,笼养个体间也表现出行为的差异。在本研究中,笼养黑叶猴的食物以树叶为主,糕料为辅,而2 只少年个体经常啃咬树皮,可能是为了摄取更多的食物[45],同时其他个体存在舔网、舔墙壁和舔水泥地面等行为,可能是为了摄取部分身体所需元素,如钙质和铁元素[32]。另外,笼养个体的食物配给方式可能会引发一些异常行为[46-47]。本研究发现3 只婴幼猴、2 只少年猴和1 只成年雌性猴出现摇晃身体的异常行为,2 只成年雄性猴、5 只成年雌性猴、1 只少年猴和2 只婴幼猴出现晃动头的异常行为,可能是笼养个体由饲养员按时投喂而不需要付出时间和能量去寻找食物,大量空余时间使笼养黑叶猴产生了异常行为[17]。此外,由于个体难以融入猴群社会,也会出现机械和呆板的、以固定频率重复和无任何功能效果的刻板行为[40]。黑叶猴在长期圈养过程中,自然群体结构较难维持,混养的方式造成异常行为增加,可能导致较高的死亡率[48]。在圈养混养蜂猴(Nycticebus coucang)中,也出现了摇晃身体、晃动头等异常行为,笼舍面积、丰容设施及不合理的性别配对和混养方式均可能是其影响原因[46]。因此,基于PAE 行为分类编码系统编制的黑叶猴PAE 行为谱可见,目前笼养黑叶猴虽然能充分表达其行为,但仍需考虑个体的差异及笼养个体的配对问题。
动物行为多样性是生物多样性的基础问题,与濒危动物的野外放归及保护相关[16],因此,笼舍面积和丰容设施的设置,对笼养个体行为的多样性表达,特别是一些弹性行为的表达,对今后提高野化放归个体的野外生存能力有重要意义[49]。此外,利用相同的方法进行野外黑叶猴行为谱的构建,并与笼养条件下黑叶猴的行为谱对比,能真正评估笼养黑叶猴行为表达空间是否足够,为提高黑叶猴的迁地保护提供更科学的依据。
猜你喜欢
四川轻化工大学学报(自然科学版)(2021年1期)2021-06-09
汉字汉语研究(2020年2期)2020-08-13
文苑(2020年5期)2020-06-16
小学生学习指导(低年级)(2020年3期)2020-06-02
电子制作(2019年22期)2020-01-14
劳动保护(2019年7期)2019-08-27
中国生殖健康(2019年10期)2019-01-07
疯狂英语·新读写(2018年3期)2018-11-29
学习月刊(2015年22期)2015-07-09
中学科技(2015年1期)2015-04-28
