
日粮中补充木聚糖酶对藏羊肉品质与肌肉发育相关基因表达的影响
2023-11-22 04:46吴振岭侯生珍桂林生
江西农业大学学报 2023年5期
周 力,吴振岭,张 宇,侯生珍,桂林生
(青海大学 农牧学院,青海 西宁 810016)
【研究意义】藏羊(Ovis aries)作为青藏高原数量最多的牲畜,已经超过5 000万只,不仅为当地牧民提供羊毛、毛皮和肉类等产品,同时对于维护高原生态系统以及藏族文化发挥着至关重要的作用[1-2]。藏羊肉的品质与整个畜牧业的发展密切相关。藏羊肉因其风味独特、味道鲜美、营养价值优越而深受消费者的青睐[3]。尽管我国藏羊的存栏量、产肉量持续上升,但仍无法满足市场的消费需求,所以如何生产更多优质的羊肉便成为当前高原畜牧业中研究的热点。【前人研究进展】小麦作为可溶性蛋白质的中间产物,可消化能量及价格都与玉米十分接近,被认为是替代玉米的适宜产品[4]。在日粮添加40%小麦能够提高育肥猪的饲料效率,降低肌肉蒸煮损失,且对养分表观消化率产生不利影响[5]。完全用小麦替代玉米不会对育肥羔羊的生长性能产生不利影响,其中以75%的小麦替代玉米可以提高其生长性能[6]。但小麦中含有较多的阿拉伯木聚糖、β-葡聚糖等抗营养因子,能够影响养分的消化吸收[7]。因此,如何消除谷物原料中的不利影响因素对于推动畜禽产业的发展尤为重要。在实践生产中,酶制剂作为饲料添加剂被广泛应用,能够达到提高饲料中养分消化率以及降低生产成本的目的[8]。其中木聚糖酶(xylanase)是一类降解木聚糖重要的酶,能够有效地分解谷物中(尤其是麦类)的阿拉伯木聚糖,从而产生分子质量大小不同的低聚木糖[9]。日粮中添加木聚糖酶可以提高羊的采食量、中性洗涤纤维消化率以及瘤胃发酵[10]。在含有高比例百慕草青贮饲料的饲粮中添加木聚糖酶可提高奶牛干物质、酸性洗涤纤维和中性洗涤纤维的体外降解率[11]。肌苷酸是肉类重要的呈鲜味物质,是评定肉品风味和新鲜度的常用指标,其含量越高,解产物越少,表明肉鲜味和新鲜度越好[12]。矿物元素在维持身体酸碱平衡,参与机体的能量代谢以及在维持神经肌肉的正常功能方面扮演着重要的作用,可以通过增强机体超氧化物歧化酶活性,减少机体自由基对肉品质的影响,从而改善肉品质[13]。另外,氨基酸的含量和种类可直接决定肌肉的营养价值[14]。【本研究切入点】本团队前期研究[15]发现,小麦型饲料中添加0.2%木聚糖酶不仅能促进藏羊胃肠道形态发育,同时还能提高其消化酶活性和抗氧化能力。此外,根据木聚糖酶产品的活性以及藏羊日粮中小麦的含量,笔者推测0.2%含量的木聚糖酶在饲喂效果以及成本控制方面为最优选择。但是有关木聚糖酶调控绵羊(尤其是藏羊)肉质性状的研究较少,仍有待进一步探讨。【拟解决的关键问题】鉴于此,本研究拟以高原型藏羊为研究对象,探索小麦-玉米型基础日粮中补充0.2%木聚糖酶能否改善其肉质特性,为外源性酶在绵羊生产中的应用提供理论依据。
1 材料与方法
1.1 试验材料
试验动物:选取初始体质量相近(19.35±2.18)kg且体况良好的2~3月龄高原型藏羊60只,试验地点:青海香咔梅朵牧业有限公司。木聚糖酶购自湖北武汉新华扬生物股份有限公司,活性为100 000 U∕g。日粮中精料补充料为粉料,粗饲料由燕麦青干草和燕麦青贮组成(按DM 基础1∶1 计),均由青海省海北州共和县香咔梅朵牧业饲料厂提供,其组成及营养水平见表1。

表1 日粮组成及营养水平(干物质基础)Tab.1 Composition and nutrient levels of the diet(DM basis) %
1.2 试验设计
将高原型藏羊公羔60 只随机分为两组,每组5 个重复,每个重复6 只羊。对照组(control group,CG)饲喂小麦型日粮,试验组(test group,TG)饲喂小麦型日粮中添加0.2%木聚糖酶。预饲期10 d 正试期90 d。正试期共分为2个阶段,第1~30天,精粗比为6∶4;第31~90天,精粗比为7∶3。
1.3 饲养管理
试验采取全舍饲单栏饲养,试验羊进行编号和驱虫,羊舍定期清理和消毒,每天饲喂2 次(08:30 和16:30),饲喂方式为人工将精料与粗料搅拌后饲喂,所有试验羊自由采食和自由饮水,每日准确记录采食量和剩料量,并清理食槽和水槽1次,管理模式和环境条件一致[16]。
1.4 样品采集
试验正试期结束后,从各组中随机选择5 只试验羊进行屠宰,参考《家畜解剖学及组织胚胎学》中方法[17],采集左侧胴体第12~13肋骨间背最长肌,剥离肉眼可见的局部脂肪与筋膜,然后置于-80 ℃,另取一份肌肉用于4%多聚甲醛固定不少于48 h。
1.5 测定指标与方法
1.5.1 肌纤维观察 将固定好的肌肉样品取出,置于冰冻切片机中,保持温度为-25 ℃,将其切成约7 µm 厚薄片,采用苏木精-伊红(hematoxylin&eosin staining,H&E.)染色法对肌纤维进行染色[18]。应用Eclipse Ci-L 正置白光拍照显微镜(Nikon,日本)在40×10 倍下观察,并通过Image-Pro Plus 6.0(media cybemetics,美国)图像软件分析肌纤维组织形态学。每个样本制作3 张切片,每张片子随机选择3~5 个组织完整的视野进行拍照。肌纤维横截面最长两点距离与最短两点距离之和的平均值,作为肌纤维直径。
1.5.2 氨基酸检测 氨基酸(amino acid,AA)含量的测定参考GB 5009.124—2016《食品安全国家标准食品中氨基酸的测定》。具体步骤如下:称取肌肉冻干粉碎样品50 mg(精确到0.1 mg)置于消解管,再加入6 mol∕L的盐酸10 mL,110 ℃烘箱内水解24 h,定容至100 mL的容量瓶中,吸取0.5 mL上述液体经氮气吹干,加入1 mL 0.02 mol∕L盐酸溶解沉淀,样品过0.22 µm滤膜进行过滤,将滤液收集于进样瓶中,置于4 ℃保存,待上机测定(日立L-8900全自动氨基酸分析仪)。
1.5.3 矿物元素测定 钙(Ca)含量的测定参考GB 5009.92—2016《食品安全国家标准食品中钙的测定》;磷(P)含量的测定参考(GB 5009.87—2016)《食品安全国家标准食品中磷的测定》;铁(Fe)含量的测定参考(GB 5009.90—2016)《食品安全国家标准食品中铁的测定》;铜(Cu)含量的测定参考(GB 5009.13—2017)《食品安全国家标准食品中铜的测定》;锌(Zn)含量的测定参考(GB 5009.14—2017)《食品安全国家标准食品中锌的测定》;硒(Se)含量的测定参考(GB 5009.93—2017)《食品安全国家标准食品中硒的测定》。
1.5.4 鲜味物质含量的测定 肌苷酸(inosine monophosphate,IMP)和肌苷(inosine,I)含量的测定采用高效液相色谱法:精确称取1 g 肉样置于离心管,加入4 mL 体积分数3.5%高氯酸后匀浆,于4 ℃条件下,4 000 r∕min离心5 min,吸取上清液至容量瓶中作为提取液,调节pH值至6.5,转移至25 mL棕色容量瓶定容,并用0.45 µm 滤膜过滤,用于上机分析。色谱条件:C18 色谱柱,流速为1 mL∕min,柱温25 ℃,进样量20 µL,检测波长254 nm。
1.5.5 肌肉发育基因表达量检测 按照TRIzol法对肌肉样品总RNA进行提取,使用核酸浓度检测仪、凝胶电泳法分别检测其浓度、纯度以及完整性,样品经质检合格后,参照TaKaRa反转录试剂盒(南京)说明书将总RNA 反转录为cDNA。采用RT-qPCR 技术检测肌肉发育相关基因的表达。以甘油醛-3-磷酸脱氢酶(glyceraldehyde-3-phosphate dehydrogenase,GAPDH)为内参基因,肌细胞生成素(myogenin,MyoG)、(growth hormone,GH)、生长激素释放激素(growth hormone releasing hormone,GHRH)和胰岛素样生长因子1(insulin like growth factor 1,IGF1)为目的基因。采用2-ΔΔCt法计算TG 和CG 组中肌肉发育基因相对表达量。条件为:94 ℃预变性30 s,94 ℃变性5 s,60 ℃退火15 s,72 ℃延伸15 s,循环40次。试验所有引物均由北京奥科鼎盛生物科技有限公司合成,其引物序列见表2。

表2 荧光定量RT-PCR引物Tab.2 Primer sequence for qRT-PCR
1.6 数据处理分析
原始数据均使用Excel 2010 整理汇总,利用SPSS 21.0 软件进行独立样本t检验,结果用平均值和标准误表示,*表示差异显著(P<0.05),ns 表示差异不显著(P>0.05),并应用Graphpad Prism 8.0.2 软件进行绘图。
2 结果与分析
2.1 日粮中添加0.2%木聚糖酶对藏羊背最长肌肌纤维直径的影响
使用H&E法对藏羊背最长肌的石蜡切片进行染色。如图1所示,应用Eclipse Ci-L正置白光拍照显微镜在40×10 倍下观察,结果发现肌肉组织学的结构较为完整,然后通过对切片图像进行统计分析,结果表明两组间肌纤维直径未达到统计学差异,但CG组比TG组高出了0.98%(P>0.05)。

图1 木聚糖酶对藏羊肌纤维直径的影响Fig.1 Effect of Xylanase on muscle fiber diameter of Tibetan sheep
2.2 日粮中添加0.2%木聚糖酶对藏羊背最长肌氨基酸含量及组成的影响
使用日立L-8900 全自动氨基酸分析仪从藏羊肌肉中共检测出17 种氨基酸。如表3 所示,CG 组中必需氨基酸、赖氨酸、蛋氨酸、缬氨酸、异亮氨酸、亮氨酸、苏氨酸及苯丙氨酸的含量均高于TG 组,但均未达到显著差异水平(P>0.05)。除了半胱氨酸外,TG 组的非必需氨基酸、天冬氨酸、谷氨酸、甘氨酸、丝氨酸、脯氨酸、丙氨酸、组氨酸、精氨酸、酪氨酸含量均高于CG 组,但差异不显著(P>0.05)。此外,各组间总氨基酸、鲜味氨基酸、必需氨基酸∕非必需氨基酸及必需氨基酸∕总氨基酸含量差异亦不显著(P>0.05)。
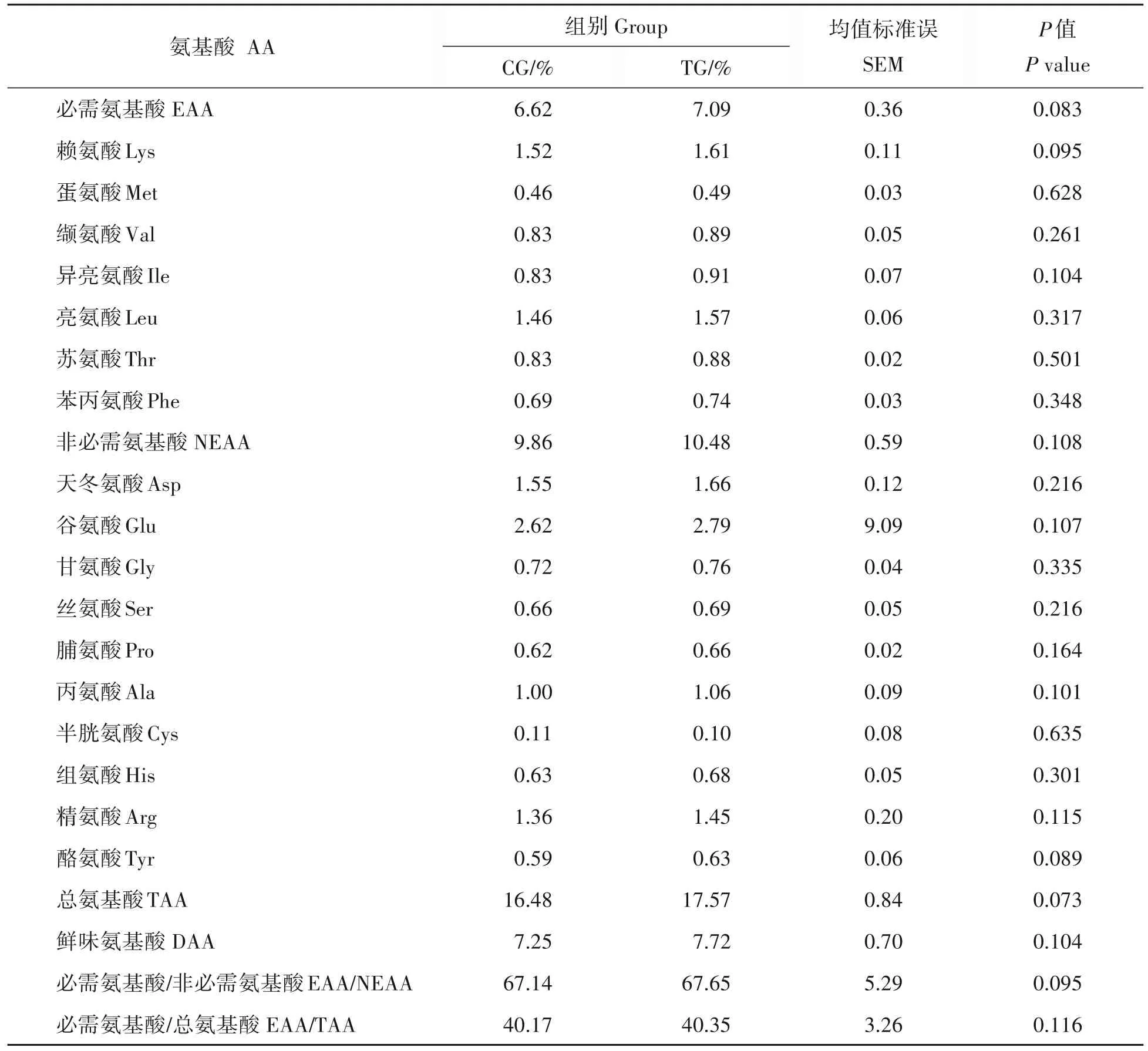
表3 木聚糖酶对藏羊肌肉中氨基酸含量及组成的影响Tab.3 Effect of Xylanase on amino acid content and composition in Tibetan sheep muscle
2.3 日粮中添加0.2%木聚糖酶对藏羊背最长肌矿物质含量的影响
如表4 所示,在微量元素中,相较于CG 组,TG 组的Fe 含量显著提高了17.84%(P<0.05)。TG 组的锌、钙含量大于CG组,而铜、硒及磷含量则呈相反的趋势,但均未达到统计学差异水平(P>0.05)。
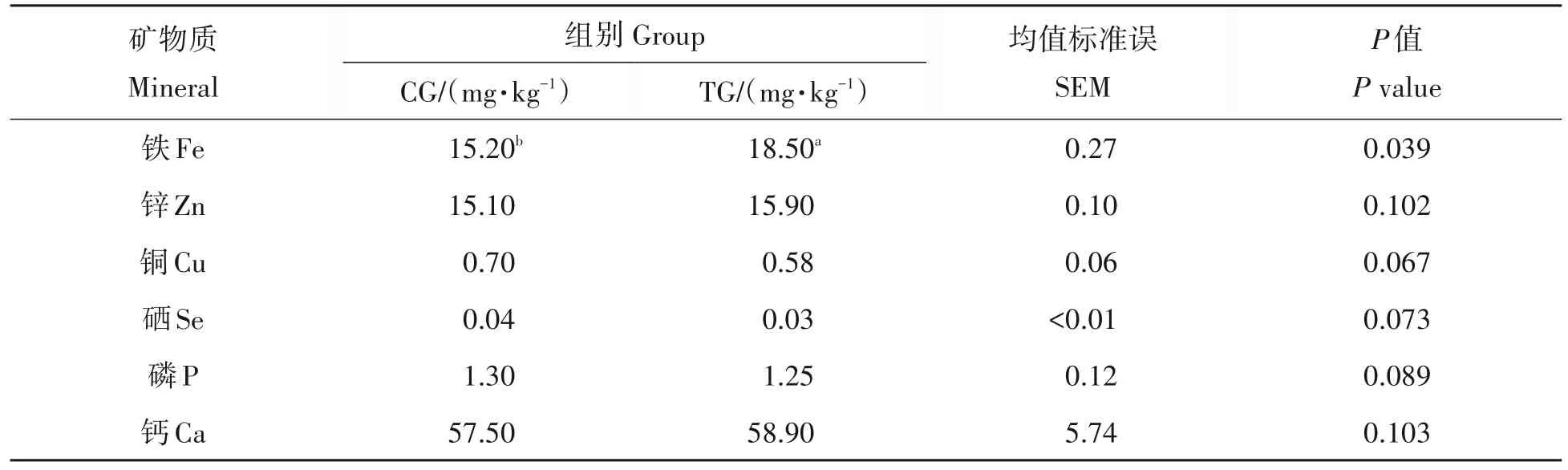
表4 木聚糖酶对藏羊肌肉中矿物质含量的影响Tab.4 Effect of Xylanase on mineral content in muscle of Tibetan sheep
2.4 日粮中添加0.2%木聚糖酶对藏羊背最长肌肌苷酸和肌苷的影响
采用高效液相色谱法检测藏羊肌肉中肌苷酸和肌苷的含量。如图2 所示,结果表明相较于CG 组,TG组的肌苷酸(IMP)含量显著提高(P<0.05)。两组间肌苷(I)含量未达到统计学差异水平(P>0.05)。

图2 木聚糖酶对藏羊肌肉中鲜味物质含量的影响Fig.2 Effect of Xylanase on the content of flavor substances in Tibetan sheep muscle
2.5 日粮中添加0.2%木聚糖酶对藏羊背最长肌基因表达量的影响
采用RT-qPCR 技术对藏羊背最长肌发育相关基因表达情况进行检测。由图3 可知,日粮中添加0.2%木聚糖酶对藏羊背最长肌中的肌细胞生成素(MyoG)、生长激素(GH)、生长激素释放激素(GHRH)以及胰岛素样生长因子1(IGF1)基因相对表达水平差异均不显著(P>0.05)。
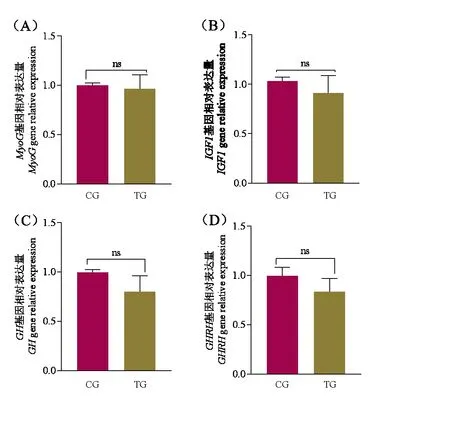
图3 木聚糖酶对藏羊肌肉发育相关基因表达量的影响Fig.3 Effect of Xylanase on the expression of muscle development related genes in Tibetan sheep
3 讨论与结论
3.1 日粮中添加0.2%木聚糖酶对藏羊背最长肌肌纤维直径的影响
肌纤维是构成骨骼肌的基本结构单位,其纤维类型受遗传、品种、营养和内部调节因子等因素相互转化[19]。据有关报道,肌纤维的组成及类型直接影响肌肉的生长发育[20]。因此,可以通过改变肌纤维类型组成来提高羊肉品质。肌纤维数目在动物出生前就已固定,此后的生长发育主要依赖于直径的增加以及类型的转化来实现[21]。研究表明,肌纤维特性与肌肉品质直接相关,如嫩度、pH 值和肉色等[22]。其中嫩度不仅是决定肌肉感官品质的重要因素,同时也是评价其多汁性的重要指标[23]。而肌纤维直径越大,说明剪切力越大,肉质嫩度也越差,反之则嫩度越佳[24]。本试验结果说明,与CG组相比,TG组的肌纤维直径有所下降。推测可能是木聚糖酶含量不足以影响肌细胞发育相关基因的表达,从而使肌纤维直径发生明显的变化。迄今为止,关于国内外在小麦-玉米型饲料中补充外源性酶制与反刍动物肌纤维特性的报道较少,还需进一步探索。
3.2 日粮中添加0.2%木聚糖酶对藏羊背最长肌氨基酸含量及组成的影响
羊肉品质指标主要包括食用品质、营养成分、氨基酸、脂肪酸等,其品质受到品种、性别、年龄及饲养方式等因素影响[25]。目前有关外源酶制剂调控羊肉氨基酸组成及含量的研究较少,所以在小麦-玉米型日粮中添加0.2%木聚糖酶对藏羊肌肉品质的影响,值得深入研究。在本试验中,两组藏羊肌肉中均检测出17种氨基酸,但CG组的TAA 含量略低于TG组。预示着木聚糖酶在调控藏羊肌肉中TAA 含量具有一定的促进作用。根据FAO∕WTO 的理想模式,组成理想蛋白质的氨基酸中EAA∕TAA 应在40%左右,EAA∕NEAA 应不低于60%[26]。在本试验中,两组间EAA∕TAA、EAA∕NEAA 虽差异不显著,均高于FAO∕WTO的标准,且以TG组含量较高。说明在舍饲条件下,两组的藏羊肉质均符合国家标准,但在日粮中补充木聚糖酶后肌肉的蛋白质量更佳。EAA不能从头合成,或者合成速率不能充分满足机体的需要,必须从膳食中获得才能满足新陈代谢的需求[27]。研究[28]证实,EAA 的含量制约着肌肉蛋白质的品质,其含量越高,说明营养价值越高。此外,羊肉的鲜美程度主要取决于Glu、Ala、Gly、Asp和Arg等鲜味氨基酸的含量[29]。其中,Glu 在决定羊肉鲜味中起主导作用[30]。本试验结果发现,两组间EAA、DAA 及Glu 含量差异不显著。
3.3 日粮中添加0.2%木聚糖酶对藏羊背最长肌矿物元素的影响
矿物元素是构成器官和组织的重要组成部分,其浓度能够衡量肌肉的营养价值。孟凡珠等[31]研究表明,年龄能对苏尼特羊不同部位的肌肉矿物质含量存在显著影响。其他研究也证实,公羔臂三头肌中多数微量元素含量、臀肌的多数常量元素与微量元素含量显著高于母羔[32]。Ca 作为骨骼的重要组成成分,能够促进骨骼的生长发育以及参与机体内多种生理作用[33]。本试验中,两组间Ca 和P 含量差异均不显著。表明木聚糖酶对藏羊肌肉中Ca 和P 含量均无显著影响。Fe 是人体内不可缺少的微量元素,能够输送氧气、促进酶合成、能量产生以及免疫调节等功能[34]。于此同时,Fe 还可促进胎儿大脑的发育和维持神经的连接[35]。在本试验中,小麦-玉米型日粮中添加0.2%木聚糖酶,明显提高了藏羊肌肉的Fe 含量,从15.20 mg∕kg 提高到18.50 mg∕kg。由此可见,木聚糖酶对藏羊肌肉中铁的沉积率的提高具有明显作用。究其原因可能是:木聚糖酶破坏饲料细胞壁,将被包裹其中的养分释放出来,供机体消化、吸收和利用所致,因相关内容较少,其调控机理还有待进一步深入研究。Cu 和Zn 在辅酶、蛋白质和脂质中起重要作用,同时还可建立肌肉、器官、血细胞和部分软组织[36]。本试验结果表明,与CG 组相比,TG 组肌肉的Cu、Zn 含量均无显著差异。说明木聚糖酶对Cu、Zn 元素在肌肉中沉积率无明显影响。另据报道,硒不仅能够在体内发挥抗氧化功能,同时还可阻止多不饱和脂肪酸过氧化,进而改善肉品质[37]。因此,从营养价值角度来说,小麦-玉米型饲料中添加0.2%木聚糖酶饲喂后藏羊的肉品质较佳。
3.4 日粮中添加0.2%木聚糖酶对藏羊背最长肌肌苷和肌苷酸的影响
随着高原畜牧业工作的持续推进,对于肉品质,特别是风味特性的研究越来越重视。肌苷酸(IMP)作为肉质风、鲜味的主要成分,已被作为评估肌肉鲜味程度的一项重要指标[38]。研究表明,日粮中添加百里香从不同程度上提高了滩羊肌肉中肌苷酸的含量,其影响效果与添加的剂量有关[39]。因此,从风味物质IMP 入手,掌握其在畜禽肌肉中合成、降解规律,对于改善羊肉质性状具有重要的理论和实践意义。在三磷酸腺苷酶的作用下,三磷酸腺苷(adenosine triphosphate,ATP)在肌肉的成熟过程中分解形成IMP[40]。研究表明,IMP 的提鲜能力是谷氨酸钠(味精)的40 倍,并且与谷氨酸钠具有很强的协同作用[41]。本试验中,TG 组的IMP 含量显著高于CG 组。推测其可能原因是:首先日粮中添加木聚糖酶被机体吸收后,转化更多的ATP,进而增加了肌肉IMP 含量;其次添加的木聚糖酶影响了IMP 代谢过程中相关酶的活性,从而提高了IMP 含量。此外,肌苷(I)是肌苷酸(IMP)在磷酸脂酶作用下分解产生的,具有一定的苦味,能对肉品风味产生不良的影响。研究发现,与基础日粮相比,中草药添加剂能够显著提高巴美羊肉中肌苷含量,这与本试验结果不一致[42]。在本试验中,相较于CG 组,TG 组的肌肉中I 含量差异不显著。进一步表明在小麦-玉米型日粮中添加0.2%木聚糖酶对藏羊肌苷含量无不利影响。
3.5 日粮中添加0.2%木聚糖酶对藏羊背最长肌基因表达的影响
肌肉发育是一个十分复杂的过程,除了受营养、遗传及环境等因素调控外,更多取决于基因表达的控制[43]。据有关报道,肉质性状的相关基因进行筛选及功能验证都可以很好的实现肉质改善[44-45]。因此,可以从基因表达水平可以进一步阐明畜禽肌肉发育的潜在分子机理。其中肌细胞生成素(MyoG)是唯一表达于所有骨骼肌细胞的因子,参与调控肌纤维的融合[46],同时也负责集中控制肌细胞分化过程,能够直接影响动物的产肉量[47]。本试验表明,两组间MyoG基因的相对表达量有所下降,但未达到统计学差异显著水平,这与肌纤维HE 染色结果相一致。IGF1作为IGF 家族中最重要的因子之一,具有多种促生长和代谢活性[48]。另外,IGF1对于调节动物产后生长至关重要,并且与肌肉生长抑制素(MSTN)共同调节肌肉的大小[49]。本试验中,与CG 组相比,TG 组的IGF1基因相对表达量差异不显著。可能是因为小麦型日粮中添加木聚糖酶含量过低,不足以消除谷物饲料中众多的抗氧化因子所致。生长激素(GH)基因调控肉的生长,能被用作肉类生长性状遗传标记的候选基因[50]。GH 的外显子4 和5 以及生长激素受体(GHR)外显子10 的基因多态性被认为与水牛的产肉量有关[51]。GHRH作为GH的正调节因子,主要促进垂体合成和分泌GH[52]。同样可以控制肌肉生长和发育,被认为是调节动物肉质性状的候选基因[53]。从本试验结果发现,两组间GH和GHRH基因的相对表达水平差异不显著。这说明小麦-玉米型日粮中添加0.2%木聚糖酶对藏羊肌肉发育相关基因表达水平影响较小。
3.6 结论
小麦-玉米型日粮中添加0.2%木聚糖酶可提高青海藏羊肌肉中Fe、IMP 含量,可改善藏羊肌肉品质。
猜你喜欢
林产化学与工业(2022年4期)2022-09-13
中国临床医学影像杂志(2022年6期)2022-07-26
中国临床医学影像杂志(2022年5期)2022-07-26
粮食与饲料工业(2021年2期)2021-04-28
国际放射医学核医学杂志(2021年10期)2021-02-28
西藏农业科技(2019年3期)2019-11-04
湖南饲料(2019年5期)2019-10-15
佛山科学技术学院学报(自然科学版)(2019年2期)2019-05-05
家禽科学(2018年9期)2018-01-16
天津农学院学报(2016年2期)2016-12-01
