
单宁酶在黑曲霉中的高效表达及其酶学性质研究
2023-08-15 01:10仝丹心何洪彬
食品工业科技 2023年15期
仝丹心,李 杰,何洪彬 ,张 会,
(1.东北农业大学生命科学学院,黑龙江哈尔滨 150030;2.山东师范大学生命科学学院,山东济南 250014)
单宁酶(单宁酰基水解酶,EC3.1.1.20)也称鞣酸酶,可在水相中水解没食子酸单宁、没食子酸脂类的酯键、缩酚酸酯键和缩酚羧键,生成葡萄糖、没食子酸和其他醇类化合物[1−4]。自然界中的单宁酶广泛存在于动物、植物和微生物体内,尤其是真菌类的曲霉属、青霉属和根霉属[5−7]。单宁酶已被列入食品工业的安全添加剂,并在食品、饲料等领域广泛应用[8−9]。可降低茶汤中单宁酸含量,解决茶饮料“冷后浑”问题[10];可降低单宁含量,解决果汁类饮料除涩、抗氧化等问题[11−14];可降低饲料中单宁酸含量,提高营养物质利用率等[15−17]。
不同来源的单宁酶酶学性质差异较大,多数单宁酶的最适温度在35~50 ℃范围内,最适pH 在5.5~7.0 范围内[18−20]。王喆丽[21]发现野生型黑曲霉中单宁酶的酶活力为0.782 U·mL−1,优化后酶活力增加至2.686 U·mL−1。林健辉等[22]以β-环糊精为碳源优化发酵培养基后的单宁酶酶活力为116.51 U·mL−1。徐璐等[23]在土壤中筛选出产单宁酶的黑曲霉Lys117高活性菌株,并对其发酵条件进行优化,优化后酶活达到233.78 U·mL−1。
黑曲霉是规模化生产重组蛋白的理想系统,被誉为“理想的真核蛋白细胞工厂”,是目前和未来工业蛋白质表达系统研究的热点[24−25]。目前单宁酶仍存在产量低、生产成本高等问题。本研究以糖化酶黑曲霉工业生产菌为宿主菌,该菌株的糖化酶基因位点为转录活跃位点,因此,以内源高表达的糖化酶基因glaA位点为基因整合的靶位点,并使用glaA 六拷贝强启动子和信号肽来提高单宁酶的表达量,并对其酶学性质进行研究,以期使单宁酶的生产性能达到国内领先水平。
1 材料与方法
1.1 材料与仪器
野生型糖化酶生产菌黑曲霉TH-2 由肇东日成酶制剂有限公司赠予;大肠杆菌感受态细胞DH5α
购于北京全式金生物技术有限公司;农杆菌AGL1
由东北农业大学真菌遗传工程研究室保存;载体pSZHG6R-AnTan、黑曲霉菌株A1(ΔglaA:AnTan)
为本研究构建;克隆载体pMD-19T、Es Taq Master Mix、限制性内切酶、DNA Maker、T4 DNA 连接酶 均购于TaKaRa 生物公司;利福平(Rifamycin)、头孢霉素(Cefalexin)、乙酰丁香酮(Acetylsyringone)、氨苄青霉素(Ampicillin)、卡那霉素(Kanamycin)、潮霉素B(Hygromycin B)均购于Sangon 公司;Protein Maker 购于Thermo 公司;质粒提取试剂盒、DNA 凝胶回收试剂盒、植物DNA 基因组提取试剂盒 均购于康为世纪生物科技有限公司;罗丹宁 购于上海aladinn 生化科技股份有限公司;柠檬酸 购于天津市东丽区天大化学试剂厂;甲醇、磷酸氢二钠 均购于天津市天力化学试剂有限公司;没食子酸正丙酯 购于南京都莱生物公司;没食子酸购于北京Solarbio 科技有限公司;葡萄糖、麦芽糖均购于西安天茂化工有限公司;糊精 购于山东萍聚生物科技有限公司;FeSO4·7H2O 购于武汉北国奉化工有限公司;ZnSO4·7H2O 购于上海铺振生物科技有限公司;CuSO4·5H2O 购于重庆名宏化工材料有限公司;MgSO4·7H2O 购于长沙锐启化工产品有限公司;MnCl2·4H2O 购于武汉吉业升化有限公司;CaCl2购于山东皓宇新材料科技有限公司;K2SO4购于湖北兴银河化工有限公司;EDTA 购于陕西晨明生物科技有限公司;SDS 购于无锡市亚泰联合化工有限公司;玉米浆 购于天津市利发隆化工科技有限公司。
DK-8D 型三温三控水槽 购于上海百典仪器有限公司;SW-CJ-1G 型超净工作台 购于上海鼎科科学仪器有限公司;DTC-3G 型PCR 仪 购于西安天隆科技有限公司;GELDoc Go 型凝胶成像系统、1658033 型蛋白质电泳系统 购于BIO-RAD 公司;UV-5100 型紫外可见分光光度计 购于上海元析仪器有限公司;DYCZ-24 型蛋白质电泳系统 购于北京六一仪器厂;A6 型匀浆器 购于上海欧河机械设备有限公司。
1.2 实验方法
1.2.1 培养基的配制 a.YEB(Yeast extract beef medium):由1 g 酵母提取物,5 g 蛋白胨,0.493 g 硫酸镁,5 g 蔗糖,5 g 牛肉膏定容至1 L,pH 调至7.0,固体培养基加入10% 琼脂粉,121 ℃高压灭菌1 h;b.PDA 培养基:由200 g 土豆(称量200 g 土豆,去皮后切成小块,加入1 L 蒸馏水,电磁炉煮30 min,纱布过滤留滤液),20 g 葡萄糖,3 g 磷酸二氢钾,1.5 g硫酸镁。固体培养基按100 mL 加入15 g 琼脂粉;c.液体发酵培养基:由10 g 葡萄糖,2 mL 玉米浆(浓度52%,蛋白质含量47%),2 g 豆饼粉定容至100 mL,pH5.0~6.0,固体培养基加入10%琼脂粉,121 ℃高压灭菌1 h。
1.2.2 引物设计 根据GenBank 单宁酶序列(Gene ID: 4990850),单宁酶基因AnTan的上下游引物P1、P2 是通过Primer5.0 软件设计而来。P3 和P4为鉴定同源重组菌株所用引物(如表1 所示)。

表1 所需引物序列Table 1 The desired prime sequence
1.2.3 基因克隆 以黑曲霉TH-2 基因组DNA 为模板,以P1 和P2 为引物,通过PCR 技术扩增出大小为1700 bp(PCR 扩增条件:预变性95 ℃ 5 min,(变性95 ℃ 30 s、退火56 ℃ 45 s、延伸72 ℃ 90 s)共30 个循环,终延伸72 ℃ 10 min,4 ℃ 2 h)的目的基因片段AnTan。基因片段AnTan和克隆载体pMD-19T 进行连接反应,将获得的质粒送到测序公司进行测序,选择Blast结果为100%的质粒进行下一步实验。
1.2.4 单宁酶基因AnTan表达载体的构建 用Xba I/Hind III 双酶切克隆载体质粒pT-AnTan,回收约1700 bp 的目的基因片段。用Nhe I/Hind III 双酶切表达载体质粒pSZHG6R,回收约15000 bp 的表达载体片段pSZHG6R,将回收的目的基因片段AnTan和表达载体片段pSZHG6R 用T4 DNA Ligase 连接,构建重组表达载体pSZHG6R-AnTan(ΔglaA:AnTan),质粒图如图1 所示。
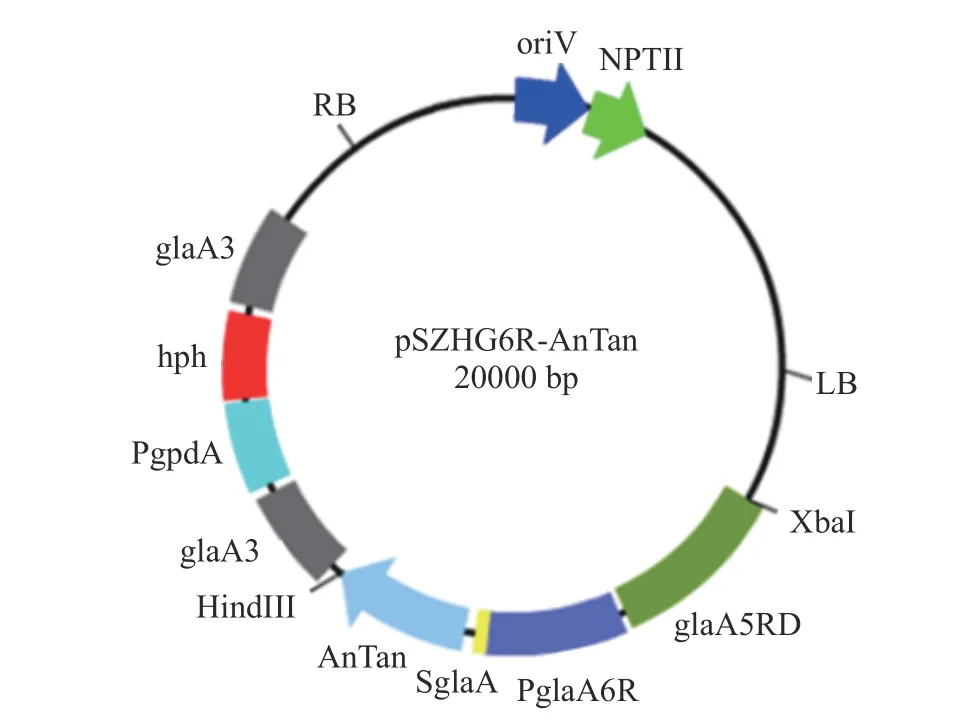
图1 pSZHG6R-AnTan 表达载体质粒图Fig.1 Plasmid map of pSZHG6R-AnTan expression vector
1.2.5 单宁酶基因AnTan黑曲霉共培养及转化子的筛选、鉴定 采用冻融法将重组表达载体pSZHG6RAnTan 转入农杆菌感受态细胞中。向新鲜灭菌的液体YEB 培养基(含100 mg·L−1Rif,100 mg·L−1km+)中加入鉴定正确的农杆菌转化子进行初次活化,28 ℃,200 r·min−1振荡培养12~14 h;另取一瓶新鲜灭菌的液体YEB 培养基(含100 mg·L−1Rif,100 mg·L−1km+和200 μmol·L−1As)接入初次活化的菌液(按1:10 的比例),28 ℃,200 r·min−1振荡培养4~5 h;待菌液摇起后吸取200~300 μL 菌液置于EP 管中,12000 r·min−1,离心1 min,留取沉淀以备使用;取1 mL 的黑曲霉TH-2 菌液,匀浆器打碎后静置15 min,取75 μL 上清和75 μL 菌丝于农杆菌沉淀的EP 管中混匀,取50 μL 的混合菌液涂布于铺有玻璃纸的PDA 固体培养基(含200 μmol·L−1As)上,放置于30 ℃恒温箱,培养2 d;取新鲜灭菌的PDA 固体培养基(含200 mmol·L−1头孢,200 mmol·L−1Hygromycin B)将覆有菌丝的玻璃纸倒置其中,30 ℃恒温箱进行培养,培养时间为12 h,玻璃纸揭去后继续培养6~8 d 直至长出黑曲霉单菌落。向液体PDA 培养基(含200 mmol·L−1头孢,200 mmol·L−1Hygromycin B)中加入长势良好的黑曲霉单菌落,培养2~3 d;待菌丝大量生长后,吸取1 mL 菌丝液于EP 管中进行基因组DNA 的提取,提取方法参考基因组提取试剂盒。以提取的基因组DNA 为模板,将P3 和P4 作为上下游引物进行PCR 鉴定,以水为阴性对照,重组表达载体质粒pSZHG6R-AnTan 为阳性对照,筛选出单宁酶基因AnTan黑曲霉纯合重组菌,标记为A1。
1.2.6 单宁酶基因AnTan黑曲霉纯合重组菌株的SDS-PAGE 检测 向装有100 mL 液体发酵培养基的锥形瓶中分别接入10 mL 的野生型菌株TH-2 和单宁酶黑曲霉重组菌株A1,然后将锥形瓶置于30 ℃270 r·min−1的培养箱里振荡培养,吸取第4 d~第11 d的发酵液上清进行SDS-PAGE 检测。
1.2.7 单宁酶酶活力测定 参考文献[26]方法,单宁酶酶活定义:30 ℃条件下,每分钟产生1 μmol 没食子酸所需单宁酶的量为一个酶活力单位,计算公式如下:
式中:U:单宁酶酶活力(μmol·mL−1),X:没食子酸浓度(μmol·L−1),D:发酵液上清总稀释倍数,t:酶解反应时间(min)。
1.2.8 重组单宁酶的酶学性质分析 将摇瓶发酵培养的100 mL 培养液于第11 d 取出,收集上清。对上清液进行抽滤,去除杂质。
1.2.8.1 最适温度 取8 份A1 发酵液上清,用pH7.0的0.2 mol·L−1磷酸氢二钠-柠檬酸缓冲液进行稀释,加入0.05 mol·L−1甲醇饶丹宁底物,分别在20、30、40、50、60、70、80、90 ℃的条件下反应30 min 后测定A1 的酶活力,计算各组酶活数值与最大值的比值为相对酶活力,绘制相对酶活力与温度曲线关系,以最大酶活力为100%[27]。
1.2.8.2 温度稳定性 取9 份A1 发酵液上清,其中8 份分别在20、30、40、50、60、70、80、90 ℃水浴锅中保温15、30、45、60 min,之后在pH 为7.0,温度为50 ℃的条件下反应30 min 后测定A1 的酶活力;另一份酶液作为对照组不进行保温处理,并定义其酶活力为100%[27],计算处理组与对照组酶活比值为残余酶活力并绘制曲线关系。
1.2.8.3 最适pH 取7 份A1 发酵液上清,分别用pH 为3.0、4.0、5.0、6.0、7.0、8.0、9.0 的0.2 mol·L−1磷酸氢二钠-柠檬酸缓冲液进行稀释,加入0.05 mol·L−1甲醇饶丹宁底物,50 ℃条件下反应30 min 后测定A1 的酶活力,以最大酶活力为100%[27]。
1.2.8.4 pH 稳定性 取8 份A1 发酵液上清,其中7 份分别用0.2 mol·L−1磷酸氢二钠-柠檬酸缓冲液调成pH 为3.0、4.0、5.0、6.0、7.0、8.0、9.0 的酶液混合液,37 ℃处理30、60、90、120 min。用pH5.0 的缓冲液稀释处理后的样品,加入0.05 mol·L−1甲醇饶丹宁底物,50 ℃条件下反应30 min 后测定A1 的酶活力,另一份酶液作为对照组未经 pH 处理,并定义其酶活力为100%[27],计算处理组与对照组比值为残余酶活力并绘制曲线关系。
1.2.8.5 金属离子和化学试剂对A1 活性的影响取9 份A1 发酵液上清,其中8 份分别添加终浓度为5 mmol·L−1KCl、MnSO4、ZnSO4、MgSO4、CaCl2、CuSO4、FeSO4、EDTA,在50 ℃、pH5.0 条件下处理1 h,研究不同金属离子和化学试剂对A1 酶活力的影响,另一份酶液作为对照组不加任何金属离子,并定义其酶活力为100%[27],计算处理组与对照组酶活比值为残余酶活力并绘制曲线关系。
1.3 数据处理
在酶活性相关实验中,每组数据处理重复三次,计算得到的酶活数据利用SPSS 软件进行单因素方差分析,以确定组间显著差异,使用Graphpad Prism v 8.0 软件绘图。
2 结果与分析
2.1 单宁酶AnTan 基因黑曲霉重组表达载体的构建
以实验室构建并保存的单宁酶基因载体质粒pSZHG-tan 为模板,以单宁酶基因引物P1 和P2 为引物进行PCR 扩增,得到大小约1700 bp 的基因片段AnTan,结果如图2 所示。将测序后的序列在GenBanK上进行Blast 序列比对分析,结果该序列与登录号XM_001401772 相似性达100%,表明单宁酶基因片段AnTan克隆成功。
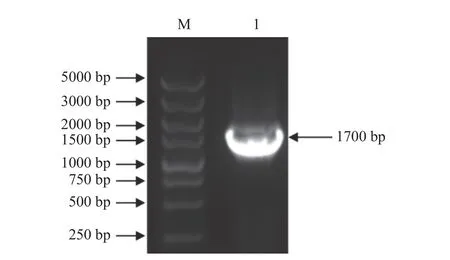
图2 AnTan 基因PCR 扩增结果Fig.2 The amplification of AnTan by PCR
将回收的目的基因片段AnTan和表达载体片段pSZHG6R 用T4 DNA Ligase 连接,构建重组表达载体pSZHG6R-AnTan,Xba I/Hind III 双酶切重组质粒,获得大小约15000 和3300 bp 的片段,结果如图3 所示,说明重组表达载体pSZHG6R-AnTan 构建成功。
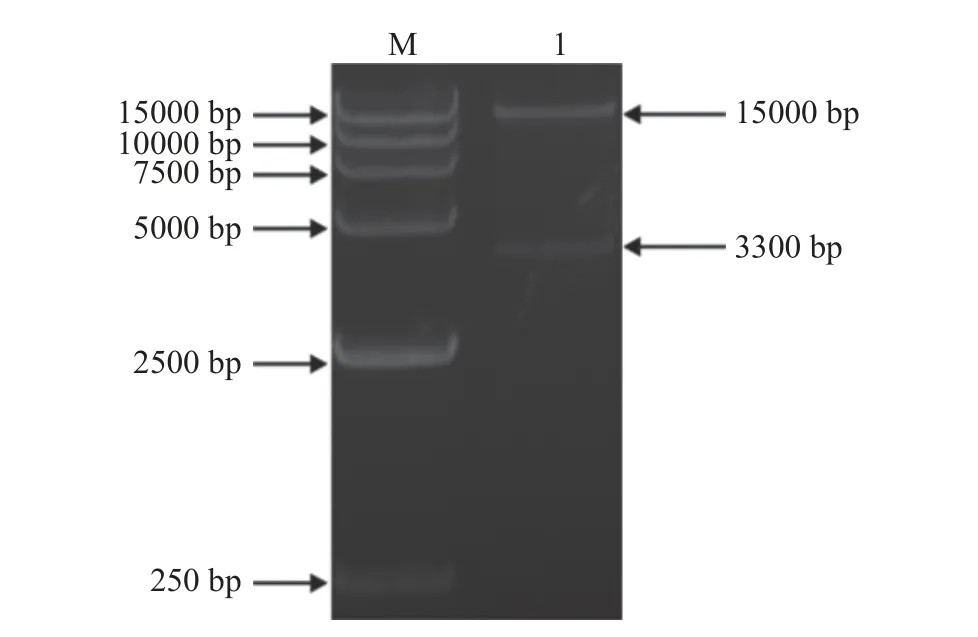
图3 pSZHG6R-AnTan 的酶切鉴定结果(Xba I/Hind III)Fig.3 Enzyme digestion of pSZHG6R-AnTan by Xba I/Hind III
2.2 单宁酶AnTan 基因农杆菌转化子的筛选及鉴定
采用冻融法将单宁酶黑曲霉重组表达载体质粒pSZHG6R-AnTan 转入农杆菌感受态AGL1 中,28 ℃培养3~4 d 至长出单菌落,挑取菌落为模板,水为阴性对照,表达载体质粒pSZHG6R-AnTan 为阳性对照,用单宁酶基因特异引物P1、P2 进行菌落PCR 鉴定。结果如图4 所示,菌落1 扩增出与阳性对照相同大小的片段,证明单宁酶黑曲霉重组表达载体pSZHG6R-AnTan 成功转入农杆菌AGL1。

图4 农杆菌转化子菌落PCR 鉴定Fig.4 Identification of Agrobacterium tumefaciens transformants by PCR
2.3 单宁酶AnTan 基因黑曲霉纯合重组菌株的筛选及鉴定
用同源臂引物P3 和P4 对重组菌株进行基因组PCR 鉴定,野生型菌株黑曲霉TH-2 和重组菌株均能扩增出约4500 bp 条带,因此,通过PCR 方法不能直接鉴定出转化子。进一步将PCR 产物用Hind III 单酶切,结果如图5 所示,泳道0 为野生型菌株黑曲霉TH-2,糖化酶基因表达框上无Hind III 酶切位点,因此切不开;泳道0+为单宁酶黑曲霉重组表达载体质粒,可将扩增出的单宁酶基因表达框切成3300和1200 bp 条带;泳道1 为重组菌株,能切出与阳性质粒一样大小的条带,而无野生型菌株4500 bp 条带,证明其为纯合的单宁酶黑曲霉重组菌株,记为A1。
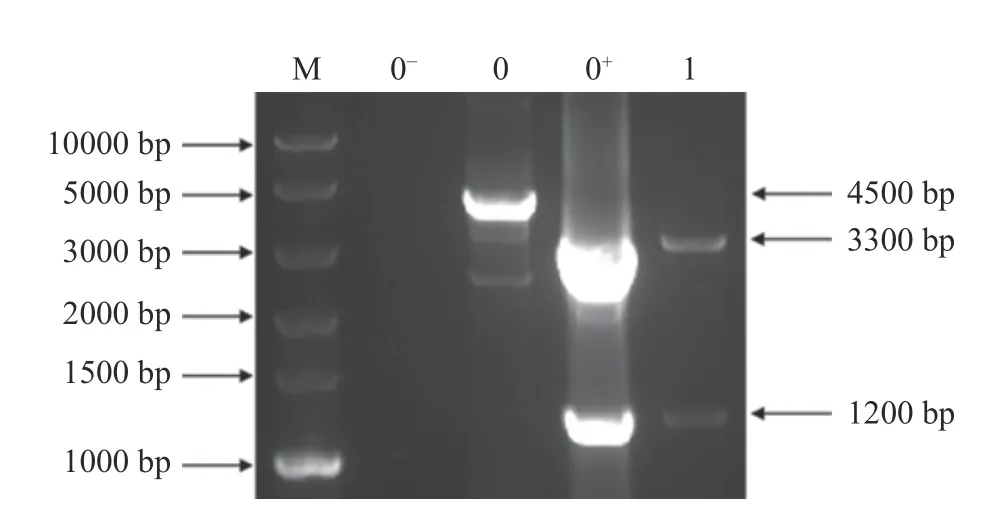
图5 黑曲霉纯合重组菌株的筛选和鉴定(Hind III)Fig.5 Screening and identification of Aspergillus niger homozygous recombinant strains(Hind III)
2.4 单宁酶黑曲霉重组菌株A1 的SDS-PAGE 检测
不改变培养基其他成分的条件下,分别以10%葡萄糖、10%糊精及10%麦芽糖为碳源,按10%接种比例分别将野生型菌株黑曲霉TH-2 和单宁酶黑曲霉重组菌株A1 进行摇瓶发酵培养,取第4~11 d的发酵上清液进行SDS-PAGE 检测,经CalMolWt 软件预测,A1 蛋白条带大小约为76 kDa,结果如图6 所示。实验室前期对野生型菌株黑曲霉TH-2 进行转录组测序和分泌蛋白质组测序分析表明,图中位置1 为糖化酶,位置2 为酸稳定的ɑ-淀粉酶,位置3 为ɑ-淀粉酶,本研究将单宁酶基因定点整合到位置1 糖化酶基因位点,所以重组菌株失去了糖化酶基因活性,与野生型菌株黑曲霉TH-2 相比,在位置4 处观察到一条明显的蛋白条带,大小约为76 kDa,与预估值相似,且没有位置1 的糖化酶蛋白带,证明重组菌株A1 为纯合同源重组菌株。

图6 重组菌株A1 的SDS-PAGE 蛋白表达电泳图谱Fig.6 SDS-PAGE detection of recombinant strain A1
2.5 单宁酶黑曲霉重组菌株A1 的酶活测定
取第4~11 d 的发酵上清液进行单宁酶活性检测,野生型菌株A.nigerTH-2 单宁酶酶活力最高为0.7 U·mL−1。重组菌株A1 单宁酶酶活力分别为75.23 U·mL−1(10%葡萄糖),134.36 U·mL−1(10%糊精),88.97 U·mL−1(10%麦芽糖),最高约为野生型菌株的192 倍,具有明确的产业化价值。以葡萄糖为基准,碳源为糊精时,单宁酶酶活力提高约78.60%,碳源为麦芽糖时,单宁酶酶活力提高约18.26%,因此,糊精是单宁酶黑曲霉重组菌株A1 的最佳碳源,结果如图7 所示。
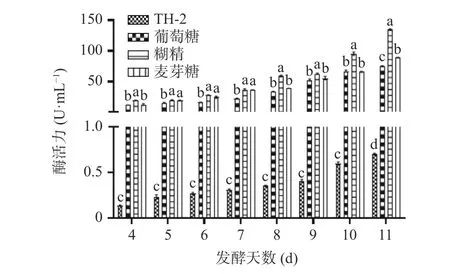
图7 A1 重组菌株酶活测定结果Fig.7 Emyze activity of recombinant strain A1
2.6 单宁酶黑曲霉重组菌株A1 中单宁酶酶学性质分析
2.6.1 温度对重组酶活性的影响及其稳定性 不同温度(20~90 ℃)下,单宁酶的活性变化差异显著,结果如图8 所示,当温度处于40~70 ℃范围内时,酶活性相对较高,说明单宁酶适宜的温度范围较广;在50 ℃时,酶活达到最大值。热稳定性结果如图9 所示,不同温度(20~90 ℃)条件下处理60 min 之后,酶在20~50 ℃范围内相对稳定,残余酶活力仍然在50%以上;60 ℃时残余酶活力维持在40%左右;酶在80~90 ℃范围内时,残余酶活不足10%。
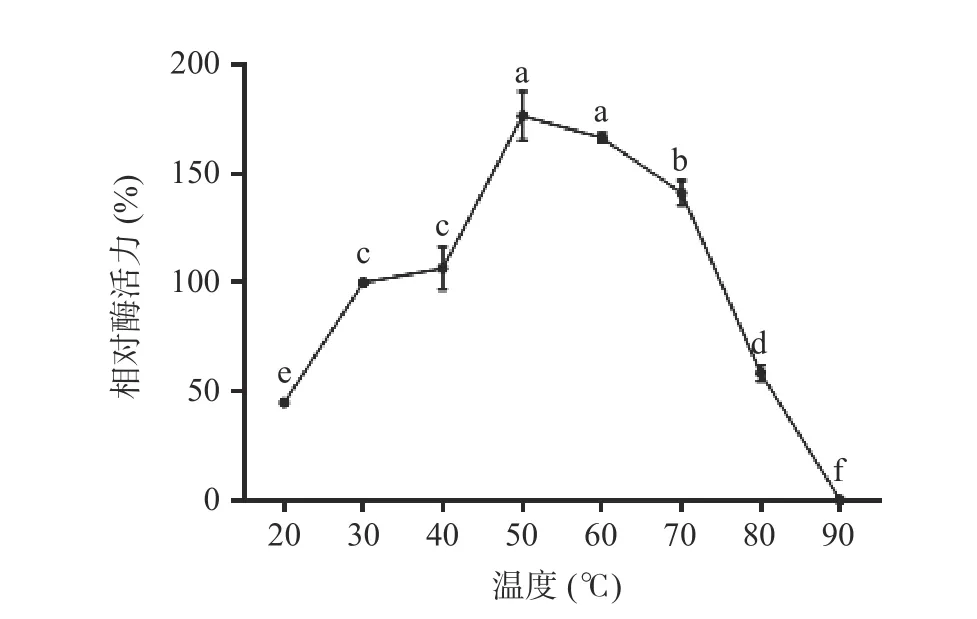
图8 不同温度下重组菌株A1 的酶活测定结果Fig.8 Enzyme activity of recombinant strain A1 at different temperature
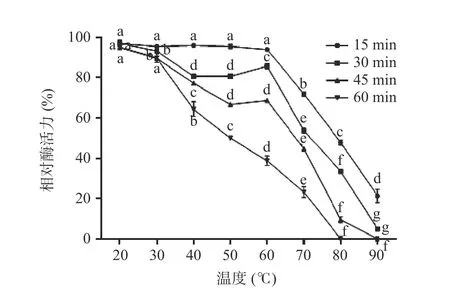
图9 不同温度对重组菌株A1 的酶活稳定性的影响Fig.9 Effect of different temperature on enzyme activity stability of recombinant strain A1
2.6.2 pH 对重组酶活性的影响及其稳定性 不同pH(pH3.0~9.0)条件下,单宁酶的活性变化差异显著,结果如图10 所示。pH 在3.0~7.0 之间酶活性呈上升趋势,并在7.0 时酶活达到最大,说明单宁酶适宜的pH 范围较广,pH 在7.0~9.0 时,其酶活呈现下降趋势。pH 稳定性结果如图11 所示,结果表明不同pH 条件下处理120 min 后,酶在pH6.0~8.0 条件下,残余酶活总体仍然在50%以上,稳定性相对较高;在pH3.0~5.0 条件下,残余酶活在40%左右;在pH 在9.0 条件下,残余酶活几乎为0。

图10 不同pH 下重组菌株A1 的酶活测定结果Fig.10 Enzyme activity of recombinant strain A1 at different pH
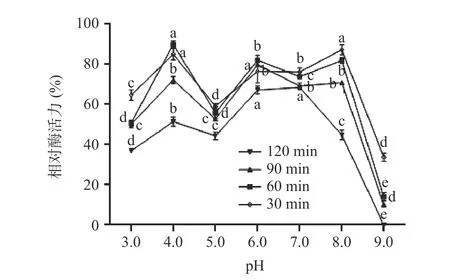
图11 不同pH 对重组菌株A1 的酶活稳定性的影响Fig.11 Effect of different pH on enzyme activity stability of recombinant strain A1
2.6.3 金属离子和化学试剂对重组酶活性的影响向重组菌株A1 的发酵培养基中加入不同的金属离子和化合物(浓度为5 mmol·L−1),结果如图12 所示。其中,检测结果表明,K+和Ca2+对重组菌株A1的酶活力没有影响,Mg2+对重组菌株A1 的酶活力有微弱的促进作用,Zn2+对重组菌株A1 的酶活力有明显的促进作用,Cu2+和Fe2+对重组菌株A1 的酶活力有微弱的抑制作用,Mn2+和化学试剂EDTA 对重组菌株A1 的酶活力有明显的抑制作用,不同的二价金属离子对酶的作用并不一致,其机理与原因有待进一步研究。

图12 不同金属离子和化合物对重组菌株A1 的酶活力影响Fig.12 Effects of different metal ions and compounds on enzyme activity of recombinant strain A1
3 结论
本研究以黑曲霉糖化酶工业生产菌为宿主菌,以内源高表达的glaA基因位点为整合靶位点,并使用内源高表达的glaA 六拷贝强启动子PglaA6R 和信号肽SglaA,获得单宁酶黑曲霉纯合重组菌株A1,最高酶活为134.36 U·mL−1,约是野生型菌株的192 倍,具有明确的产业化价值。获得的重组单宁酶最适作用温度是50 ℃,最适作用pH 为7.0。Zn2+对重组酶有明显的促进作用,Mn2+和EDTA 对重组酶有明显的抑制作用。未来可进一步通过更换信号肽、基因融合、密码子优化、增加基因拷贝数、优化培养条件及使用蛋白酶缺陷菌株等来提高重组蛋白的产量。通过多重策略的组合使用解除重组蛋白在转录和翻译水平的限制,以期进一步提高单宁酶的表达量,实现单宁酶的工业化生产。
猜你喜欢
——“单宁”
山东国资(2020年6期)2020-07-09
食品与生物技术学报(2019年12期)2019-02-15
食品工业科技(2018年21期)2018-11-27
中国酿造(2016年12期)2016-03-01
中国酿造(2016年12期)2016-03-01
大连工业大学学报(2015年4期)2015-12-11
应用化工(2014年11期)2014-08-16
中国酿造(2014年9期)2014-03-11
中国酿造(2013年10期)2013-04-23
食品科学(2013年17期)2013-03-11
