
木槿开花过程中花色素的变化规律及其影响因素
2023-07-05 10:36:36李浙浙张一丹王郑昊程蓓蓓
植物研究 2023年4期
李浙浙 张一丹 王 波 王郑昊 王 璐 程蓓蓓*
(1.河北科技师范学院,秦皇岛 066000;2.河北省特色园艺种质资源挖掘与创新利用重点实验室,秦皇岛 066000;3.泰安市时代园林科技开发有限公司,泰安 271000;4.泰安市徂徕山林场,泰安 271000)
花色是观赏植物重要的观赏特征和表型性状,也是品种分类的重要依据[1]。植物在开花过程中,花色常发生变化,虽然有较多的研究关注了花色变化现象[2],但较少对花色变化产生的内在机制进行深入探究[3-4]。花色素的种类是花色形成的物质基础,种类繁多且复杂,主要包括类黄酮[5]、类胡萝卜素[6]和生物碱[7]3类。除花色素的种类和含量外,pH、金属离子等也是影响花色呈色的重要因素[8-9]。大量研究表明,不同植物、同种植物不同花色品种花色素的组成和含量具有较大差异,而这种差异与花色多样性密切相关[10-11]。同时花色多样性与植物品种间丰富的遗传多样性密切相关[12],并受到特定基因调控[13-14]。
木槿(Hibiscus syriacus)为锦葵科(Malvaceae),木槿属(Hibiscus)多年生灌木或小乔木,是我国北方广泛种植的夏季重要木本花卉之一。木槿花色丰富,盛开时满树繁花,极具观赏价值。大部分木槿花在开放过程中,花色通常会发生变化。陈含等[15]对不同花色木槿的抗氧化性进行探究,结果表明总黄酮是影响木槿抗氧化性的主要因素。赵艺璇等[16]利用SRAP 法,在分子水平上对39 个木槿品种进行鉴定,结果表明不同木槿品种间亲缘关系较近。目前关于木槿花色的研究相对较少,主要集中在花色分离模式[17]等方面。在分子层面上,陈家龙等[18]对于白花品种‘点雪’和粉花品种‘粉红巨人’的花瓣进行了转录组测序和分析,确定了与花色调控相关的基因。而在木槿开花过程中花色变化的物质基础和影响因素却鲜见深入的研究,这在一定程度上影响了关于木槿花色变化的基础研究。
为进一步探究木槿开花过程中花色变化规律和影响木槿花色变化的因素,本研究以‘千丝绊’和‘白色雪纺’木槿为试验材料,通过对开花过程中花色素的含量和影响花色因素的探究,欲解决以下问题:(1)开花过程中不同花色的木槿花色变化的规律;(2)影响木槿花色变化的理化因素;(3)木槿开花过程中花色变化与色素成分的变化是否存在相关性。
1 材料与方法
1.1 试验材料与试剂
试验材料为2021 年7 月,采摘于河北科技师范学院昌黎校区的‘千丝绊’(H.syriacus‘Qian⁃siban’)和‘白色雪纺’(H.syriacus‘White Chiffon’)木槿,裂开期(S1)、初开前期(S2)、初开期(S3)、盛开期(S4)、谢花期(S5)共5个开花时期的花瓣。采摘后,立即带回实验室进行试验。各时期的划分标准参考杨晨[19]在牡丹(Paeonia×suffruticosa)开花过程中的时期划分,分别为:S1,花苞处于发育阶段,花苞小,萼片半包;S2,花苞处于发育后期,花苞大、未开放;S3,花苞未完全展开,呈半开放状;S4,花苞完全绽放;S5,花朵闭合,颜色加深,易从枝上掉落。
标准品为矢车菊素和芦丁(纯度≥98%),均购自上海源叶生物科技有限公司。
1.2 试验方法
1.2.1 花色表型测定
利用色彩色差仪(SP62,美国)测定‘千丝绊’和‘白色雪纺’不同时期的木槿花瓣的内表皮中央部位明度L*值,色相a*和b*值[20]。根据公式C*=(a*2+b*2)1/2计算彩度C*值。每个时期测定3 个样本,每个样本测定3次。
1.2.2 特征显色反应
取0.1 g的‘千丝绊’和‘白色雪纺’不同时期的木槿花瓣(去除丹心雌雄蕊)鲜样,在液氮下研磨成粉末后放入10 mL 离心管,分别加入石油醚、10%盐酸和30%氨水,观察溶液颜色变化。试验重复3次[21-24]。
1.2.3 含水量测定
采摘新鲜的‘千丝绊’和‘白色雪纺’不同时期的木槿花瓣(仅花冠部分)各6 朵,称量,将其放入干燥箱中,在105 ℃下,烘干12 h至恒质量后,再次称量,计算‘千丝绊’和‘白色雪纺’不同时期的木槿花瓣的含水量。试验重复3次。
1.2.4 金属离子含量测定
取烘干的‘千丝绊’和‘白色雪纺’不同 时期的木槿花瓣(仅花冠部分)0.2 g,使用消解液(V(盐酸)∶V(硝酸)∶V(高氯酸)=3∶1∶1)20 mL 进行消解后,采用电感耦合等离子体发射光谱仪(Opti⁃ma2100DV,日本)测定钙、镁、锌、铁、铜、锰等离子质量分数。
1.2.5 过氧化物酶(POD)、多酚氧化酶(PPO)含量测定
称取0.5 g‘千丝绊’和‘白色雪纺’不同时期的木槿花瓣(仅花冠部分)鲜样于研钵中,加入3 mL经4 ℃预冷的磷酸缓冲液(0.05 mol·L−1,pH=7.8)研磨成匀浆,转移到5 mL 离心管中,定容。4 ℃下12 000 r·min−1离心20 min,收集上清液待测,POD采用愈创木酚法、PPO 采用儿茶酚法测定[24]。试验重复3次。计算公式如下:
其中:OD值为470 nm 下的吸光度,m为样品鲜质量(g),t为反应时间(min),V1为提取时酶液体积(mL),V2为反应时酶液体积(mL)。
其中:ΔOD为反应时间内OD值的变化,t为反应时间(min),Vt为样液总体积(mL),Vs为测定时样品的体积。
1.2.6 pH测定
根据Zhao 等[25]的方法,取0.1 g 新鲜的‘千丝绊’和‘白色雪纺’不同时期的木槿花瓣(仅花冠部分),加入5 mL的去离子水,在研钵中研磨呈匀浆,转移到10 mL离心管中,在4 ℃下,12 000 r·min−1离心5 min,将上清液转入新的10 mL离心管中,使用pH 计(数显PHS-25,上海)测定‘千丝绊’和‘白色雪纺’不同时期的木槿花瓣的pH。试验重复3次。
1.2.7 花色苷总含量测定
花色苷总含量参考洪燕红[26]的方法。取不同时期的‘千丝绊’和‘白色雪纺’木槿(仅花冠部分)鲜样各0.1 g,分别用液氮研磨成粉末后转移到10 mL 离心管中,加入5 mL 1%盐酸甲醇溶液,振荡摇匀后于冰箱中4 ℃黑暗条件下充分浸提24 h(期间多次震荡)后离心15 min(4 ℃,13 000 r·min−1),取上清液测定在A530 和A657 处的吸光值。花色苷总含量以A530-0.25·A657 表示。试验重复5 次。
1.2.8 花色素组分和含量测定
采用液相色谱−质谱联用进行黄酮类物质定性定量测定[19,27]。样品前处理同1.2.7。
液相条件:柱温40 ℃,进样量10.0 μL,流速0.50 mL·min−1,350、520 nm 波长下扫描吸收光谱。流动相参数:A 为体积分数为1%的甲酸水溶液,B 为色谱级乙腈。梯度洗脱程序:0 min,95%A+5%B;40 min,60%A+40%B;45 min,95%A+5%B;分析时间80 min。
质谱条件:采用正离子模式,电喷雾离子源,干燥器温度400 ℃,离子源温度120 ℃,干燥气流速度10 L·min−1,喷雾气压力50 psi,毛细管裂解电压3.5 kV,锥孔电压40 V,质量数扫描范围:100~1 200 m/z。
以标准矢车菊素和芦丁为对照,以浓度为横坐标,以峰面积为纵坐标,制作标准曲线,计算其花色苷和黄酮类物质的含量。其中,矢车菊素含量标准方程为:y=528.57x−9.676 2,R2=0.998 9;芦丁含量标准方程为:y=534.58x+499.02,R2=0.996 4。
1.3 数据分析
试验结果采用Excel 2019、SPSS 26.0、DPS 9.01软件进行数据处理,采用Duncan’s 新复极差法对不同时期各指标数据进行差异显著性分析、采用皮尔逊系数对花色表型和影响因素进行相关性分析。
2 结果与分析
2.1 花色变化
如图1 所示,‘千丝绊’木槿S1 期花色为紫红色,随着花朵的开放,颜色逐渐变为粉紫色,最后变为紫色;‘白色雪纺’木槿在S1期和S2期为淡黄色,从S3期一直到S5期都为白色。由表1可知,在‘白色雪纺’木槿开花过程中,其亮度L*在逐渐升高,在S5 期达到最高,且各时期的亮度L*有着显著性差异(P<0.05);而粉色品种‘千丝绊’的亮度L*值整体上是升高的,同样在S5 期达到最大值。2 个品种的色彩度C*在开花过程中呈现逐渐下降,表明其色彩度C*越来越不丰富,颜色越来越单一。

表1 ‘千丝绊’和‘白色雪纺’木槿开花过程中花色表型特征Table 1 Floral phenotype statistics of H. syriacus ‘Qiansiban' and ‘White Chiffon' during flowering

图1 ‘千丝绊’和‘白色雪纺’木槿开花过程中花色变化Fig.1 Flower color changes of H. syriacus ‘Qiansiban’and H. syriacus ‘White Chiffon’during flowering
2.2 特征显色反应
由图2 可知,经10%盐酸处理后(图2a),‘千丝绊’溶液显粉红色,说明‘千丝绊’花瓣中含有花色苷;‘白色雪纺’溶液呈无色,说明其中不含花色苷。在经石油醚处理后(图2b),两个溶液都无色,说明两个品种的木槿花瓣中不含类胡萝卜素。在经30%氨水处理后(图2c),两个溶液呈现不同程度的黄色,说明两个品种的木槿花瓣中含有类黄酮。

图2 木槿花的特征显色反应左侧为‘千丝绊’右侧为‘白色雪纺’;a.10%盐酸溶液;b.石油醚溶液;c.30%氨溶液Fig.2 Characteristic color reaction of flowers of H. syriacus On the left was H. syriacu ‘Qiansiban’and on the right was H. syriacus ‘White Chiffon’;a.10% hydrochloric acid solution;b.Petroleum ether solu⁃tion;c.30% ammonia solution
2.3 花色素的组成与含量
2.3.1 花青素的组成及含量
在520 nm的波长下,分析‘千丝绊’木槿和‘白色雪纺’木槿5 个时期花瓣提取液,共检测到5 种花色苷,具体结果见表2。

表2 ‘千丝绊’木槿花瓣中花色苷的液质结果Table 2 Results of anthocyanins in the petals of H. syriacus ‘Qiansiban’
由表2 可知,‘千丝绊’木槿5 个时期花瓣中共检测出5种花色苷,通过对质谱分子离子和碎片离子分析,结合参考文献[28-30],共推测出5 种花色苷物质,分别为:飞燕草素-3-O-葡萄糖苷(De3G)、矢车菊素-3-O-葡萄糖苷(Cy3G)、牵牛花素-3-O-葡萄糖苷(Pe3G)、天竺葵素-3-O-葡萄糖苷(Pg3G)、锦葵素-3-O-葡萄糖苷(Ma3G)。
由图3可知,‘千丝绊’木槿不同时期花色苷的组成未发生变化,但含量却发生了明显的变化。不同时期的‘千丝绊’木槿花中锦葵素-3-O-葡萄糖苷都是最高的,其次是牵牛花素-3-O-葡萄糖苷。这5种花色苷的含量随着开花过程都在逐渐减少,S1~S5期,分别减少了46%、52%、40%、64%和39%。

图3 ‘千丝绊’木槿花瓣中花色苷各成分质量分数Fig.3 Mass fraction of anthocyanins in the petals of H.syriacus ‘Qiansiban’
2.3.2 黄酮类物质的组成
在350 nm 波长下,分析‘千丝绊’和‘白色雪纺’木槿5 个时期花瓣提取液,共检测到10 种黄酮类物质,具体结果见表3。

表3 ‘千丝绊’和‘白色雪纺’木槿花瓣中黄酮类物质的液质结果Table 3 Results of flavonoids of H. syriacus ‘Qiansiban’and ‘White Chiffon’
由表3 可知,在‘千丝绊’和‘白色雪纺’木槿5个时期花瓣中共检测出6 种苷元,10 种黄酮类物质。通过对各组分的质谱分子离子和碎片离子分析,结合参考文献[19,27,31],共推测出6 种苷元,分别为:槲皮素、木犀草素、圣草酚、山奈酚、芹菜素和牡荆苷。确定了10种黄酮类物质分别为:槲皮素-3-葡萄糖苷、木犀草素-7-O-葡萄糖苷、圣草酚-7-O-葡萄糖苷、山奈酚-O-六碳糖-C-六碳糖苷、芹菜素-C-二葡萄糖苷、木犀草素-3,7-二葡萄糖苷、芹菜素-7-O-芸香糖苷、牡荆苷、山奈酚-3-葡萄糖苷和山奈酚-C-六碳糖苷。
由表4可知,‘千丝绊’木槿的黄酮类物质种类比‘白色雪纺’木槿丰富,其中芹菜素-C-二葡萄糖苷和芹菜素-7-O-芸香糖苷是两种木槿所共有的且含量较高的黄酮类物质,这两者在‘白色雪纺’木槿中的含量高于在‘千丝绊’木槿中的含量。在黄酮类物质种类方面,‘千丝绊’木槿的种类多于‘白色雪纺’木槿的种类。‘白色雪纺’木槿中除山奈酚-3-葡萄糖苷外,其余4 种黄酮类物质均在S1 期达到最大值。
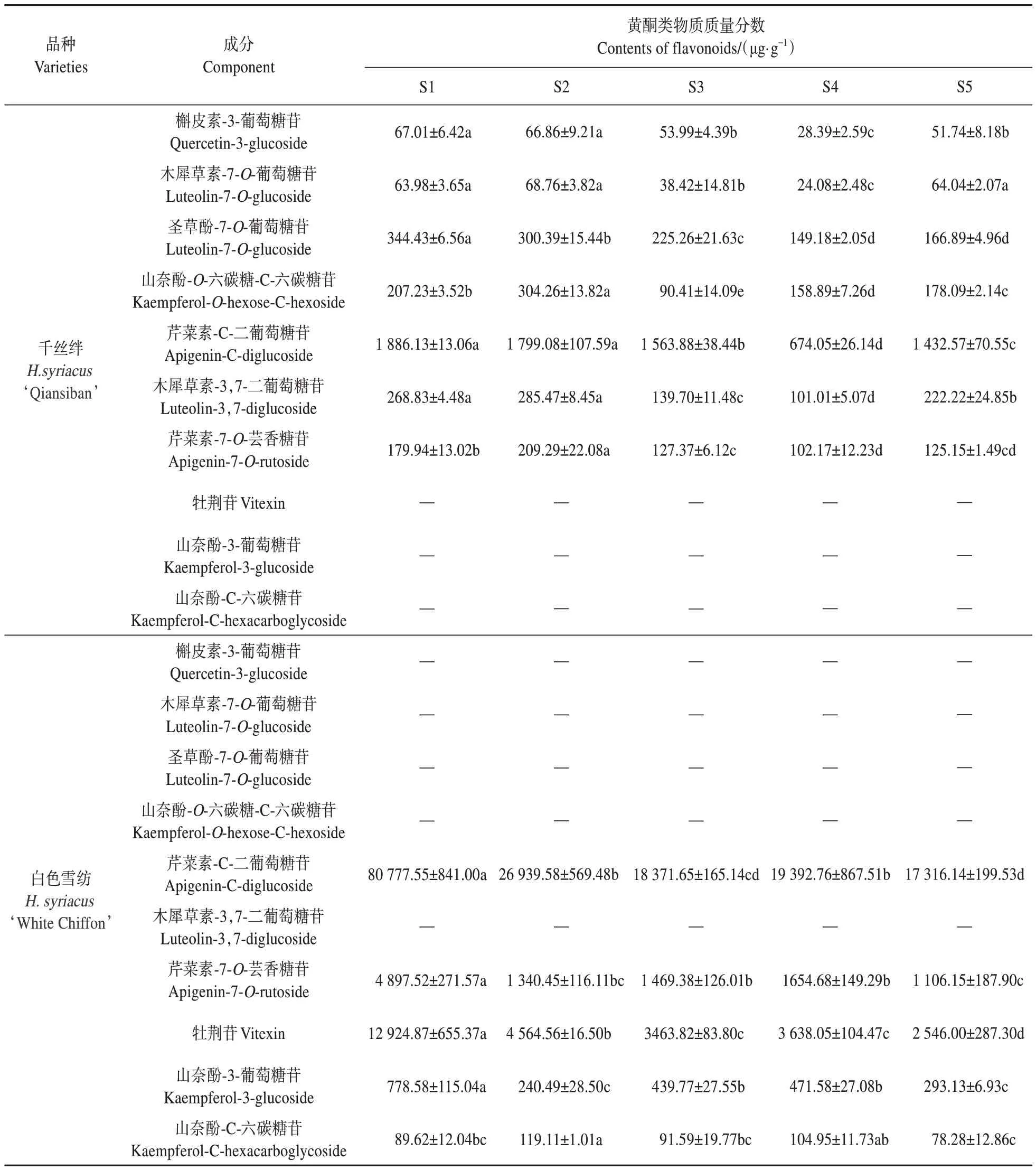
表4 ‘千丝绊’和‘白色雪纺’木槿花瓣中黄酮类物质的质量分数Table 4 Contents of flavonoids of petals in H. syriacus ‘Qiansiban’and ‘White Chiffon’
2.4 理化因子对木槿开花过程颜色变化的影响
木槿花色受多种因素的共同影响,由表5 可知,‘千丝绊’和‘白色雪纺’木槿的含水量在80%~95%,且后者高于前者。两个品种木槿花瓣的pH差异性不显著,均表现酸性,介于5.50~5.80。两个品种木槿的POD和PPO活性均表现显著性差异,其活性值分别介于0.20~1.50 和2.00~17.00 U·g−1·min−1,POD 活性在‘千丝绊’木槿中均高于‘白色雪纺’木槿,而PPO 活性则相反,PPO 的活性在两类木槿中均在S3 期到达最大值,分别为5.47 和3.63 U·g−1·min−1,而POD 活性在S5 期达到最大值,分别为0.61 和1.62 U·g−1·min−1。‘白色雪纺’木槿不含有花色苷,‘千丝绊’木槿花色苷总含量在S1~S5期,含量逐渐下降,结合表1,发现木槿花在开放过程中,色彩越来越单一。在金属离子含量方面,钙离子和镁离子是木槿花瓣中含量最高的金属离子,铜、铁、锰、锌等金属离子在木槿花瓣中含量相对较低。在‘白色雪纺’木槿中,钙离子、镁离子含量均在S5 期达到最高,分别为2 222.16 和1 126.01 mg·kg−1;在‘千丝绊’木槿中,钙离子、镁离子在S3 期达到最大值,分别为4 186.26 和1 263.73 mg·kg−1。铁、锌、铜和锰离子含量在两个品种的木槿中‘千丝绊’木槿要高于‘白色雪纺’木槿,其中在S1~S4 期,均为在‘千丝绊’木槿的含量高于‘白色雪纺’木槿,在S5 期则刚好相反。在钙离子含量方面,‘白色雪纺’木槿整体上高于‘千丝绊’木槿中钙离子的含量,而在S3 期,‘千丝绊’木槿中钙离子的含量高于‘白色雪纺’木槿中的钙离子含量。

表5 影响木槿开花过程的理化因子Table 5 Physicochemical factors affecting the blossoming of H. syriacus
2.5 相关性分析
木槿开花过程中花色表型和影响因素进行相关性分析见表6。由表6 可知,在‘千丝绊’木槿开花过程中,色相a*与色彩度C*呈极显著正相关(P<0.05),随着a*值的增大,C*值也增大,说明色相a*是影响‘千丝绊’木槿色彩度的主要因素。在金属离子方面,钙离子、铁离子和锰离子都与色彩度C*呈显著性正相关(P<0.01),表明随着这3 种金属离子的含量越高,‘千丝绊’木槿的C*值越大,色彩度越丰富。‘千丝绊’木槿花色表型与POD、PPO 活性、含水量、pH 无显著相关性,说明其不是影响‘千丝绊’木槿花色表型的主要因素。在‘白色雪纺’木槿开花过程中,色彩度C*与色相b*和PPO 活性呈极显著正相关(P<0.01),与色相a*呈极显著负相关(P<0.01),说明b*值越大,C*值也就越大,PPO 活性越强,C*值也越大。色相a*、色相b*和PPO 活性呈极显著负相关(P<0.01),表明a*值随着b*值的增大和PPO 活性的增强,逐渐变小。‘白色雪纺’木槿花色表型与POD 活性、pH、金属离子含量和含水量均无显著相关性,说明其不是影响‘白色雪纺’花色表型的主要因素。

表6 影响因素与木槿开花过程中花色表型的相关性Table 6 Correlation analysis between influencing factors and flower color phenotype of H. syriacus during flowering
3 讨论
在开花过程中,花色的变化常与花瓣中的色素成分和含量密切相关。本研究表明在‘千丝绊’木槿开花过程中,花色苷含量的降低是造成花色由紫红色变为粉紫色直至紫色的主要原因。如杨琴等[20]对两个牡丹(Paeonia×suffruticosa)品种的研究发现,在开花过程中其花瓣从红色向黄色方向变化,其色素种类并未发生变化,主要是由于花青素含量的剧烈降低造成的;周兵等[3]在对紫茉莉(Mirabilis jalapa)的研究中发现,紫红色的紫茉莉花由花蕾期的淡绿色变为盛开期的紫红色,主要与叶绿素含量的降低和花青素含量的升高密切相关。以上研究结果在一定程度上支持了本研究结论。黄酮类物质在花中主要作为助色素,辅助花色素的呈现,但其本身呈现淡黄色。邓娇[32]发现黄酮和黄酮醇在黄色莲花(Nelumbo lutea)形成中起重要作用,除此以外,木犀草素、芹菜素-7-O-葡萄糖苷、山奈酚-3-O-葡萄糖苷等物质在金花茶(Camellia petelotii)[33]、菊花(Chrysanthemum×mori⁃folium)[34]等植物中高表达,而在‘白色雪纺’木槿中,其黄酮和黄酮醇类物质含量减少,花瓣颜色从淡黄色变成白色,推测其可能是因为黄酮类物质含量的变化造成的,但具体原因还需要进一步研究,才能确定其花色变化的具体原因。本研究发现pH 和POD 活性对于两个品种木槿开花过程中花色变化没有影响,但PPO 活性与‘白色雪纺’木槿花色表型相关。在陈小红等[35]对鸳鸯茉莉(Brunfelsia latifolia)开花过程中花色变化的生理机制,发现pH 和POD 活性对于植物花色的变化也起着一定的作用。研究发现,随着pH、POD 活性的逐渐升高,花色苷逐渐降解或酚类物质被氧化为醌,花色会朝着白色转变。在本研究中,不同时期木槿花瓣的pH 相差不大,推断pH 不是影响木槿木槿花色变化的原因。另外对于PPO 的研究[36]表明,PPO 可以氧化酚类物质,从而使花色苷降解。但PPO 活性是否与‘白色雪纺’木槿花瓣从淡黄色变成白色有关,还需要进一步研究,才能确定其影响因素。
在金属离子含量方面,本研究发现钙、铜、铁和锰离子与‘千丝绊’木槿花色变化相关。李芹梅等[8]对滇水金凤花(Impatiens uliginosa)花色变化的原因进行了探究,结果发现除花色苷的含量外,铜离子含量也是影响滇水金凤花花色变化的原因,铜离子主要通过影响控制合成花色苷的基础代谢物的含量来影响滇水金凤花的花色。除此以外,在绣球花(Hydrangea macrophylla)[37]、红色新几内亚凤仙花(Impatiens hawkeri)[38]、威氏绿绒蒿(Meconopsis wilsonii)[39]中都验证了铜、铝等金属离子含量会对植物花色产生影响,造成花色的变化。在木槿花中,金属离子是否是通过控制合成花色苷的基础代谢物的含量或者与花色苷产生络合物,进而来影响‘千丝绊’木槿的花色,还需要进一步探究。
木槿花色丰富,大部分木槿花在开花过程中都会发生花色变化的现象,本试验结果表明,色素的组成、含量及金属离子和酶活性与木槿花色的变化有着密切的关系,其中花色素的组成和含量起着决定性作用,但本研究仅分析了部分影响因素与花色之间的关系,今后还可以从解剖结构、分子遗传等方面对木槿花色变化机制进行深入的研究。本研究在一定程度上为揭示木槿花的呈色机制,为探究木槿花色变化过程中的相关调控基因奠定了基础,也为木槿的分子育种和扩大木槿的种质资源的利用提供参考。
猜你喜欢
语文世界(初中版)(2023年8期)2023-09-09 23:48:01
天津文学(2023年8期)2023-04-29 16:28:26
安徽文学(2023年4期)2023-04-11 03:49:44
青年文学家(2022年35期)2022-02-13 23:54:51
中老年保健(2018年3期)2018-07-12 03:26:20
现代家庭·生活版(2017年4期)2017-04-20 21:51:04
诗林(2016年5期)2016-10-25 06:07:54
小资CHIC!ELEGANCE(2016年12期)2016-06-17 20:11:34
少年文艺(2015年6期)2015-05-28 04:45:25
流行色(2014年3期)2014-06-23 08:37:06
