
不同树龄侧柏古树扦插生根过程中的生理响应
2023-07-05 10:36:32常二梅刘建锋贾子瑞赵秀莲
植物研究 2023年4期
常二梅 董 遥 刘建锋 贾子瑞 李 香 赵秀莲 郭 伟,4*
(1.林木遗传育种国家重点实验室,国家林业和草原局林木培育重点实验室,中国林业科学研究院林业研究所,北京 100091;2.中国林业科学研究院森林生态环境与自然保护研究所,国家林业和草原局森林生态环境重点实验室,北京 100091;3.北京市绿地养护管理事务中心,北京 102211;4.泰安市泰山林业科学研究院,泰安 271000)
古树能够保持旺盛的生命力通常具有抗逆或长寿等特性,可以作为林木抗逆的候选基因材料。但随着树龄的增长古树长势渐弱,甚至死亡[1],因此保存古树的优良种质资源已迫在眉睫。扦插操作简单、效率高是古树无性繁殖的主要方式,但是部分古树如侧柏(Platycladus orientalis)等扦插生根率极低[2]。研究古树的扦插不定根形成过程中的生理机制对提高扦插生根率有重要作用。
碳水化合物、含氮化合物、激素等为插穗的生根提供能量和营养[3-4]。银杏(Ginkgo biloba)母树的树龄越大,总糖含量和生根率越低[5]。日本落叶松(Larix kaempferi)难生根的无性系总氮含量少[6],营养物质在调节细胞生长和分化中有重要功能,其含量在不定根发生过程中调节植物的形态建成[7]。因此研究古树插穗总糖、可溶性蛋白等物质并针对性地补充营养物质为古树插穗生根提供能量保障。
环境胁迫产生过量的活性氧(ROS),而过氧化物酶能清除有害的ROS,保证植物组织正常生长[8-9]。其中超氧化物歧化酶(SOD)是一种与衰老有密切关系的抗氧化剂,SOD 活性上升诱导糙叶杜鹃(Rhododendron scabrifolium)的插穗愈伤组织形成[10]。酚氧化酶(PPO)是促进形成生根辅助因子“IAA-酚酸复合物”的氧化还原酶,也是不定根形态建成的重要影响因子[11]。过氧化物酶(POD)是木质素合成过程中的关键酶,并且是不定根发生和伸长的重要影响因子[12]。悬铃木(Platanus orientalis)和柚木(Tectona grandis)的插穗POD 活性随着母树树龄增加而下降[8,13],因此氧化酶活性能清除插穗基部过量ROS促进古树扦插生根。
内源吲哚乙酸(IAA)对根原基诱导、愈伤组织形成及根伸长均起着重要作用[14]。随着白皮松(Pinus bungeana)树龄的增加,扦插生根率显著降低,插穗的内源脱落酸(ABA)含量升高,而IAA、玉米素核苷(ZR)、赤霉素(GA)含量降低[15]。白桦插穗中的ω(IAA)/ω(ABA)的比值随着母树树龄的增加而降低[16],而麻楝(Chukrasia tabularis)、菲油果(Acca sellowiana)的ABA 含量过高抑制了根原基发育和不定根的伸长[17-18]。创伤胁迫引起的GA含量变化可触发局部细胞变化启动再生[19-20]。因此了解古树在扦插过程中的激素含量变化有助于提高古树扦插生根率。
侧柏的寿命长达几百甚至几千年,蕴藏着长寿等优良基因。因此保存侧柏古树的种质资源能极大地发挥古树的生态和经济价值[2]。然而侧柏古树树龄越大,扦插过程中插穗越容易出现腐烂、愈伤组织形成困难、生根率低等问题[21]。因此侧柏古树扦插过程中插穗的营养物质、抗氧化酶、内源激素含量对生根率影响的机制急需研究。本研究通过树龄为5、100、300、700 年侧柏母树插穗扦插过程中的关键时间点取材,分析插穗的生理生化变化从而探讨生根机制,为提高侧柏古树扦插生根率提供理论依据。
1 材料与方法
1.1 试验材料
试验材料采自位于中国林业科学研究院和北京植物园内的侧柏母树,树龄分别为5、100、300、700 年左右(根据古树树龄记载),每个年龄选择9 株进行采样。其中5 年生侧柏插穗作为对照试验材料。6月份采集生长健壮、无病虫害的距地面3~5 m 的侧枝上1~3 年生新梢。截成10~15 cm 插穗,在离芽1.0~1.5 cm的基部斜切,备用。
1.2 试验设计及取样方法
插穗用m(NAA)∶m(IBA)=1∶1 质量浓度为1 000 mg·L−1浸泡处理1 min。采用完全随机区组设计,每个处理25 个插穗、3 次重复。扦插试验在中国林科院内的全光照喷雾下进行,管理参照杜常健等[22]进行操作。基质选用体积比是2∶8 的泥炭和珍珠岩的混合物的轻基质网袋容器。根据前期实验观察到的扦插过程中的形态变化,将5、100、300、700 年生侧柏母树插穗中的不定根形成过程分为4 个阶段:扦插试验当天设为第0 天,在0 d(S1,扦插前)、35 d(S2,愈伤组织形成期)、65 d(S3,不定根形成期)、95 d(S4,不定根伸长期),对每个重复随机选取6个插穗进行样本取材。拔出插穗后,用蒸馏水冲洗干净,用滤纸擦干,迅速剥取插穗基部2~3 cm韧皮部,剪碎混匀后保存在液氮中。
1.3 测定方法
1.3.1 营养物质测定
称取0.1 g 新鲜样本,加入1 mL 提取液研磨匀浆,离心取上清。采用生理指标试剂盒(苏州科铭生物技术有限公司),利用酶标仪,根据试剂盒说明书和参考杜常健等[22]进行操作在540、620 nm 波长下测定不同树龄扦插过程中的总糖、总蛋白吸光值。根据试剂盒的计算公式计算总糖、总蛋白的含量。
1.3.2 酶活性测定
称取0.1 g 新鲜样本,加入1 mL 提取液,进行低温研磨匀浆,离心取上清。采用生理指标试剂盒(苏州科铭生物技术有限公司)测定酶活性并计算。根据试剂盒说明书操作,PPO 活性的测定原理是PPO 催化邻苯二酚产生醌,利用酶标仪,调节波长至525 nm 进行测定;SOD 活性测定采用NBT法,通过黄嘌呤及黄嘌呤氧化酶反应系统产生超氧阴离子(O·−2),O·−2可还原氮蓝四唑生成蓝色甲臜,后者在560 nm 处有吸收;SOD 可清除O·−2,从而抑制甲臜的形成,在560 nm处测定吸光值;POD的测定原理是POD 催化H2O2氧化特定底物,在470 nm测定吸光值;采用硫代巴比妥酸−分光光度法,测定丙二醛(MDA)含量,利用MDA 与硫代巴比妥酸缩合,生成红色产物,在532 nm处有最大吸收峰,进行比色后可估测样品中过氧化脂质的含量,同时测定600 nm下的吸光度,利用532、600 nm下的吸光度的差值计算MDA的含量。
1.3.3 内源激素含量测定
称取0.2 g 样品,用预冷的含10 μg·L−1抗氧化剂BHT 的80%冷甲醇溶液浸提4 h,过滤后,将处理液用冰盒送到中国农业大学作物化控中心,采用酶联免疫法(ELISA)进行内源激素含量测定,测定结果采用Logit曲线方法进行计算。
1.4 数据处理
采用SPSS 22.0 软件的单因素方差分析(Oneway ANOVA)对试验数据开展单因素方差分析,采用Turkey法比较不同组间的差异性。
2 结果与分析
2.1 不同树龄侧柏插穗扦插生根情况
为了探讨母树树龄对扦插生根率的影响,对不同树龄的侧柏开展扦插试验。结果显示5 年生侧柏插穗的生根率和生根数量分别为63.5%和6.12,显著高于古树扦插的生根率和生根数量(P<0.05)。100、300、700 年生侧柏插穗生根率分别为20.32%、18.55%、13.74%(图1A),生根数量分别为3.26、2.81、2.24(图1B)。结果表明不同树龄的扦插生根率和生根数量没有显著差异,而幼树扦插生根能力显著高于古树。
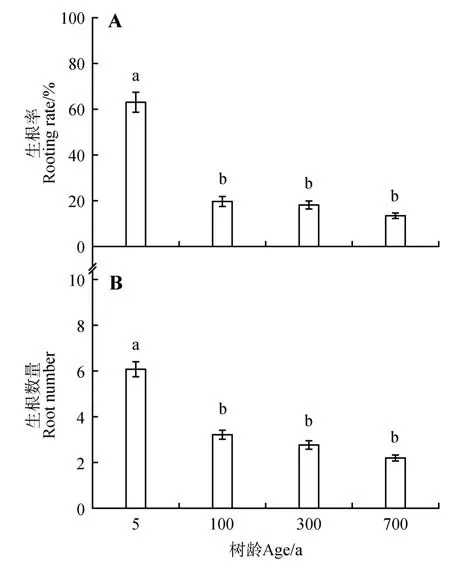
图1 侧柏树龄对插穗生根的影响不同小写字母代表处理之间在P<0.05水平存在显著性差异;数值为平均值±标准差Fig.1 Effects of age on rooting and cuttings of P. orientail Different lowercase letters in the same column represented significant difference at P<0.05 level;Data presented as mean±SD
2.2 不同树龄侧柏插穗扦插过程中营养物质含量变化
营养物质为插穗在生根过程中提供能量。结果表明,营养物质包括总糖和总蛋白含量都随着插穗母树树龄的增加而减少。5 年生侧柏插穗的营养物质显著高于古树(P<0.05)(图2A),说明5年生的插穗能供给较多的营养物质促进不定根形成。100、300、700 年生侧柏插穗(P>0.05)(图2B)营养物质含量差别不显著,说明树木到一定树龄后生理状态差异不显著。
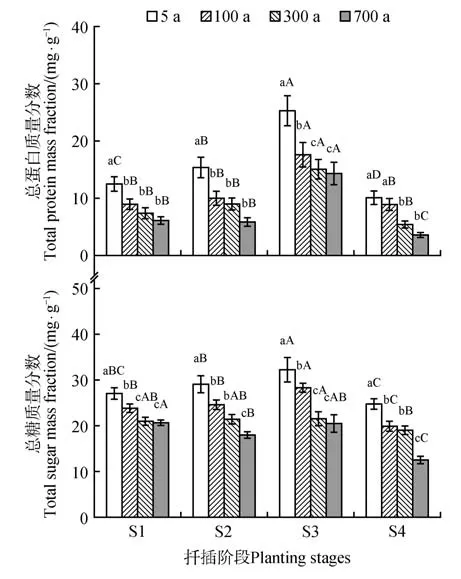
图2 扦插过程中总糖和总蛋白质量分数变化不同大写字母表示不同生根阶段在同一树龄的差异显著(P<0.05),不同小写字母表示不同树龄在同一生根阶段差异显著(P<0.05);下同Fig.2 Changes of total sugar and total protein mass frac⁃tion during the cutting rooting process Different capital letters indicated significant difference among differ⁃ent rooting stages in the same tree age(P<0.05);Different lowercase letters indicated significant difference among different tree ages in the same stage(P<0.05);the same as below
侧柏插穗的总糖和总蛋白含量随着扦插进程(S1~S4)呈先升高后降低的趋势。在S2 时不同树龄插穗的总糖和总蛋白含量呈上升趋势,S3 不定根形成促使插穗中的总糖和总蛋白增强。这可能是因为插穗地上部组织的光合作用加强,产生的营养物质运输到插穗基部,使插穗基部的营养物质积累。S4时侧柏插穗的总糖和总蛋白含量呈现下降趋势,可能是因为根系在生长过程中消耗的总糖多于叶片光合作用合成的营养物质。
2.3 不同树龄侧柏插穗扦插过程中抗氧化酶活性变化
抗氧化酶能清除有害物质提高抗性,辅助不定根形成。结果显示,5年生侧柏的抗氧化酶包括PPO、SOD、POD 活性增加显著高于古树(P<0.05)(图3)。插穗的抗氧化酶活性随着侧柏母树树龄的增加而下降。不同树龄侧柏插穗在S1 和S2 时PPO 活性呈现降低的趋势(图3A),5、100 年生的侧柏插穗的PPO 活性在各阶段变化趋势相似,呈现出先降低后升高的趋势,而300、700年生侧柏插穗的PPO 活性的变化趋势相似,呈现出持续下降的趋势。在S4 时5 年生侧柏插穗的PPO 活性大幅度上升。在不定根形成期和不定根伸长期,PPO活性的升高有利于“IAA-酚酸复合物”的形成,从而促进不定根的形成和伸长。

图3 扦插过程中抗氧化酶活性变化Fig.3 Changes of activities of antioxidative enzymes dur⁃ing the cutting rooting process
所有树龄侧柏插穗随着扦插生根的进程SOD活性上升(图3B),其中5 年生侧柏插穗的SOD 活性上升的幅度显著高于古树(P<0.05),在S2~S4显著大于S1。S4 时所有侧柏母树树龄插穗的SOD活性达到峰值,说明在扦插过程中插穗的SOD 活性升高,起到了促进插穗抗逆和不定根发生的作用。
POD 能消除体内有害的自由基,利于愈伤组织和不定根的形成。在S1~S2 时,5、100 年生侧柏插穗的POD 活性随生根的进程显著上升(P<0.05)(图3C),S3 和S4 时所有插穗的POD 活性上升,5、100 年生侧柏插穗的POD 活性显著高于300、700年生(P<0.05),这说明POD 活性上升有助于氧化IAA促进不定根形成。
2.4 不同树龄侧柏插穗扦插过程中MDA含量变化
MDA 是膜脂过氧化的产物,含量越高表明膜脂受到的伤害越严重。侧柏插穗的MDA 含量随着插穗母树树龄的增高呈现升高的趋势(图4)。在扦插过程中,MDA 含量呈现先上升后下降的趋势。S3 时5 年生侧柏插穗的MDA 含量继续升高,而其余树龄插穗的MDA 含量下降,其中300 年生侧柏插穗MDA含量下降的幅度较大。S4时5年生侧柏插穗的MDA含量降低,侧柏古树插穗MDA含量升高。这可能是因为插穗在扦插初期的机械损伤,MDA含量上升,在愈伤组织形成和不定根发生时,插穗的生理状态逐步恢复,并且对环境的适应能力开始增强,导致MDA含量降低。
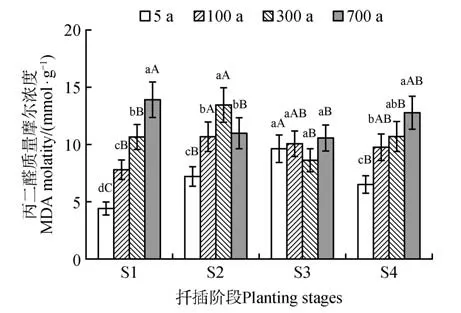
图4 扦插过程中MDA质量摩尔浓度变化Fig.4 Changes of MDA during the cutting rooting process
2.5 不同树龄侧柏插穗扦插过程中内源激素含量变化
侧柏插穗的激素含量随着插穗母树树龄的增加而呈现出不同的变化趋势(图5A)。不同树龄侧柏插穗之间IAA 含量差异显著(P<0.05)。来自5、100、300 年生侧柏插穗的IAA 含量在扦插4 个阶段,呈先上升后下降的趋势,S2时来源于不同树龄侧柏插穗的IAA 含量上升,促使内源IAA 向插穗基部积累;在S3 时IAA 含量达到峰值。S4 时5、100、300 年生插穗的IAA 含量却下降,可能是因为不定根在伸长过程中消耗了大量IAA。另外不同树龄侧柏插穗的ZR 含量在扦插4 个阶段,总体呈现上升的趋势(图5B)。在扦插过程中侧柏插穗的ZR 含量随着插穗母树树龄的增加而下降。在S2~S4 时5 年生侧柏插穗的ZR 含量显著大于古树插穗的ZR含量(P<0.05),这可能是较高浓度的ZR促进了细胞分裂。

图5 扦插过程中激素质量分数变化Fig.5 Changes of hormones mass fraction in the cutting rooting process
在扦插过程的4 个阶段GA 含量呈现降低—升高—降低的趋势,S2 时所有树龄侧柏母株插穗的GA含量都呈现下降趋势;S3时各树龄侧柏插穗GA 含量均呈上升趋势,因此GA 在一定程度上促进侧柏不定根的发生(图5C)。
随着侧柏母树树龄增长,插穗ABA 含量呈现增加的趋势(图5D),而在整个扦插过程中,5 年生侧柏插穗的ABA 含量显著低于古树插穗的ABA含量(P<0.05),说明侧柏古树插穗ABA 含量较高从而抑制扦插生根。在不定根形成期和不定根伸长期,所有树龄侧柏插穗的ABA 含量在S3 时最低,这说明ABA 含量的降低有利于扦插过程中插穗的不定根形成和伸长。
2.6 不同树龄侧柏插穗扦插过程中ω(IAA)/ω(ABA)、ω(ZR)/ω(ABA)比值的变化
不同树龄侧柏插穗的ω(IAA)/ω(ABA)、ω(ZR)/ω(ABA)趋势相似,比值总体呈上升趋势(图6)。在S1 时各树龄侧柏差异较小,在S2~S4 时5 年生侧柏插穗的ω(IAA)/ω(ABA)、ω(ZR)/ω(ABA)比值大幅上升。且上升幅度显著大于100、300、700 年生侧柏插穗的ω(IAA)/ω(ABA)、ω(ZR)/ω(ABA)比值(P<0.05)。在S4 时5 年生侧柏插穗的ω(IAA)/ω(ABA)比值上升幅度变小,而100、300、700 年生侧柏插穗的ω(IAA)/ω(ABA)比值却呈显著下降趋势(P<0.05)。通常情况下,ω(IAA)/ω(ABA)、ω(ZR)/ω(ABA)比值越高,插穗越容易生根。在此扦插试验的过程中,5 年生侧柏插穗的ω(IAA)/ω(ABA)比值最高,这促进了5 年生侧柏插穗生根,而700 年生侧柏插穗的ω(IAA)/ω(ABA)比值最小,因此,其生根率较低,这也是各种激素之间相互作用的结果。

图6 扦插过程中IAA/ABA、ZR/ABA比值变化Fig.6 Changes of ratios of IAA/ABA、ZR/ABA during the cutting rooting process
3 讨论
古树随着树龄的增长扦插生根率骤降,许多古树如:甘蒙柽柳(Tamarix austromongolica)、银杏等扦插生根率极低[23-24],内源生长物质减少是降低扦插生根能力的主要原因之一。插穗内部充足的营养物质是根原基形成以及快速生根的基础[25]。本研究发现侧柏古树随着母树树龄的增加插穗的营养物质含量呈现下降趋势;这与白桦、长白落叶松(Larix olgensis)树龄增长扦插生根率降低的结果[16]相似。在对秤锤树(Sinojackia xylocarpa)扦插繁殖研究中发现,生根过程消耗插穗内部大量的糖和蛋白[26],这与本研究侧柏插穗总糖含量提高有利于愈伤组织形成和不定根发生的结果一致。有研究发现来自5、100、300、700 年生侧柏插穗在愈伤组织形成期,可溶性糖含量上升,并且5 年生侧柏插穗的上升幅度最大[21],与本研究的结果一致。插穗内营养物质为插穗不定根的形成提供了物质保障,而且正是这些变化推动了插穗的生根进程。侧柏古树扦插生根率低可能是因为营养物质含量不能满足不定根形成和伸长。
抗氧化酶能消除自由基等抑制生根的物质[27]。插穗的SOD、PPO、POD 活性随着插穗母树树龄的增加而降低。在侧柏扦插过程中(S1~S4),PPO、SOD、POD活性呈先上升后下降的趋势。5年生侧柏插穗的抗氧化酶活性显著高于古树,这与扦插生根率趋势一致,因此抗氧化酶活性高对扦插生根具有促进作用。POD 酶活性峰值的出现对马尾松(Pinus massoniana)不定根的发生起到了明显的促进作用[28]。扦插前期高活性的SOD 有利于增强百香果(Passiflora edulis)插穗抗性,促进插穗切口愈合和愈伤组织的形成[29],扦插后期SOD 的活性越高对杜仲(Castanea mollissima)不定根的形成越有利[30],这两项结果均与本研究的试验结果相一致。另外,栓皮栎(Quercus variabilis)根原基的PPO 活性高有利于诱导不定根的形成[27]。MDA值越大表示电解质的渗漏量越多,细胞膜受害程度越重,侧柏古树插穗抗氧化酶活性低而MDA 含量高说明其各种不良环境抵抗能力弱导致生根困难。
IAA 能够促进插穗内部营养物质的重新分配促进插穗生根[31]。本研究中5、100、300 年生侧柏插穗的IAA 含量呈先上升后下降的趋势,在S3 时IAA含量达到峰值,说明IAA对于不定根的诱导和伸长具有促进作用。此结果与山杏(Armeniaca si⁃birica)和北美香柏(Thuja occidentalis)的扦插试验结果[32-33]一致。ZR 对不定根的发生有促进作用,ZR 含量随着侧柏插穗母树树龄的增加而下降。在整个扦插过程中,4 个树龄段的侧柏插穗的ZR含量呈现上升的趋势,这与四倍体刺槐(tetraploidRobiniapseudoacacia)和白皮松(Pinus bungeana)扦插试验结果[15,34]相一致。IAA 和ZR 保证了细胞分裂、分化、形态建成过程中所需的物质。
GA 和ABA 含量与扦插过程中的机械损伤有关,插穗的ABA 含量增加以增强插穗的抗逆性。本研究结果显示5 年生侧柏插穗的GA 含量显著低于侧柏古树,因此含量低可能对不定根的形成有促进作用。这与波叶金桂(Osmanthus fragrans)和兴安圆柏(Sabina davurica)扦插时低浓度的GA有利于低树龄母树插穗不定根发生的试验结果[7,35]一致。在S3 时GA 含量高说明创伤能够诱导器官的再生和防御[19-20]。在不定根形成期和不定根伸长期,侧柏古树插穗的ABA 含量在S3 时达到最高值,说明ABA 对于不定根的诱导和伸长具有一定的抑制作用。这与ABA 含量过高抑制山杏(Armeniaca sibirica)、菲油果根原基发育和不定根伸长的结果[32,18]一致。而杂交鹅掌楸(Lirioden⁃dron chinese×L.tulipifera)IAA/ABA 升高时有利于插穗不定根的形成[36]与侧柏古树的结果相似,说明各种激素在侧柏古树扦插过程中相互影响,适合适量的激素促进不定根生成。
4 结论
扦插保存侧柏古树优良种质资源发挥古树的生态和经济价值具有重要作用。通过研究不同树龄侧柏古树扦插生根关键时间点的生理生化变化,发现有利于生根的物质如:营养物质(总糖、总蛋白)、抗氧化酶(PPO、POD、SOD 活性)、激素含量(IAA、ZR)在随着树龄的增长各个扦插生根阶段逐渐降低。MDA、GA和ABA呈现先升高后降低的趋势,这些物质在不定根形成阶段含量最高致使侧柏古树扦插生根难。说明有利于生根的营养物质、抗氧化酶、激素在侧柏古树插穗中含量较低,而有害物质较多。因此,在古树扦插过程中施加适量的营养物质(总糖、总蛋白)、激素(IAA)有助于提高古树生根率。
猜你喜欢
花卉(2024年1期)2024-01-16 11:33:50
林业科学研究(2023年4期)2023-04-29 00:44:03
作物研究(2022年1期)2022-11-27 23:34:38
新农民(2022年22期)2022-11-07 23:38:06
作物学报(2022年10期)2022-07-21 03:13:58
现代农业研究(2021年10期)2021-11-02 14:40:24
山东林业科技(2021年4期)2021-08-31 05:55:04
星星·诗歌原创(2021年3期)2021-06-01 09:13:23
天然产物研究与开发(2018年4期)2018-05-07 06:48:03
福建林业科技(2018年1期)2018-04-09 00:56:41
