
水杉凋落物水浸提液对其种子萌发和生长的化感作用
2022-12-30 07:19徐来仙周大寨郭秋菊夏煜轩
广西植物 2022年11期
徐来仙, 姚 兰, 周大寨, 郭秋菊, 朱 江, 邓 楚, 艾 鑫, 夏煜轩
(湖北民族大学 林学园艺学院, 湖北 恩施 445000 )
化感物质主要是通过植物分解、根际分泌、雨水淋溶和挥发等方式释放到环境中的水溶性物质(Rice, 1984;Kimura et al., 2015),其中凋落物是林木化感物质的重要来源之一,其在分解过程中释放内含物质或转化为次生物,从而对种子萌发和幼苗生长产生影响,进而成为林木天然更新障碍的关键因素之一(Caviers et al., 2007;Muturi et al., 2017;Aguilera et al., 2017)。凋落物化感作用的浓度效应主要表现为对植物种子发芽及幼苗生长的“低促高抑”双重效应(李登武等,2010;庄正等,2017),或随浓度升高而抑制增强的“抑制效应”,从而导致其天然更新出现障碍(张珊珊等,2016;郭晓燕等,2019)。不同部位、分解阶段凋落物也具有不同的化感效应(Fernandez et al., 2006;Huang et al., 2019)。
水杉(Metasequoiaglyptostroboides)为柏科(Cupressaceae)水杉属(Metasequoia)孑遗植物,是植物界的“活化石”,为中国特有,属国家Ⅰ级保护树种。现存水杉原生母树仅分布于湖北省利川市、湖南省龙山县和重庆市石柱县之间狭窄的三角区域内,仅存5 696株。水杉原生母树周边林下及利川市小河水杉母树管理站、利川市林业科学研究所人工营造的35~42 a生水杉无性系种子园中,均很难发现有天然更新的水杉幼苗及幼树存在(林勇等,2017)。这一现象引起学术界对水杉天然更新障碍及更新繁育的极大关注。许多专家学者从水杉生长环境与干扰(胡先骕和郑万钧,1948;林勇等,2017;陈俊等,2020)、种群与遗传结构(黄小等,2020;刘小红,2020)、交配与扩散格局(陈文文,2016)等方面进行了探索,另外也从水杉更新繁育方面进行了大量研究,如水杉原生母树个体及种子种源差异(吴漫玲等,2020)、温度(辛霞等,2004)、光照(郭秋菊等,2018)、水分(Fan et al., 2020)、硒元素(郭秋菊等,2018)、整地覆土(李淑娴等,2012)等对水杉种子萌发的影响。但是,目前水杉天然更新困难的机制仍不完全明确。
通过对水杉母树主要分布区调查发现,水杉林下有大量凋落物,仅偶见1~2株水杉幼苗。木本植物种群的天然更新是在自然力作用下,新个体不断产生而使种群得以繁衍和扩大的重要方式,而种子萌发和早期成苗阶段是植物天然更新最关键的两个阶段,这两阶段又受林下凋落物产生化感作用的影响(Aliskan et al., 2015),并且化感效应在不同阶段随着释放的化感成分变化而变化(Fernandez et al., 2008)。水杉母树周围累积的凋落物所产生的化感作用是否会成为水杉天然更新障碍的因素,为此,本研究针对不同类型水杉母树凋落物设置不同浓度的水浸提液对水杉种子进行萌发试验,以探究水杉凋落物化感作用对水杉种子萌发与生长的影响,为进一步查明水杉天然更新困难障碍提供科学依据。
1 材料与方法
1.1 种子和凋落物的收集与处理
试验所用水杉种子为2020年11月从水杉原生母树1 076号(树龄118 a,树高31 m,胸径87.6 cm,生长地理位置为108°35′45.1″ E、30°07′20.5″ N,海拔1 114 m)采集。种子含水量为9.78%± 1.13%,千粒重为(2.57±0.17) g。
为保证收集凋落物的代表性,在湖北省利川市林业科学研究所水杉种子园内按东南西北中五个方向分30个区域随机布设凋落物收集框采集。2020年10月末,采集水杉母树上自然脱落的当季凋落物为新鲜凋落物,即尚未进入分解过程,包括叶、枝、果、皮等混合物;2020年11月末,收集水杉林内地表的凋落物为自然凋落物,包括未分解层、半分解层和全分解层枝叶果皮等的混合物。所有采集回来的凋落物通过室温风干后,分别进行相应处理。
1.2 方法
1.2.1 凋落物水浸提液制备 首先,将新鲜凋落物和自然凋落物分别风干、粉碎(过100目筛),以蒸馏水为溶剂,按料液比1∶5加入蒸馏水,于室温条件下振荡48 h;然后,10 000 r·min-1离心10 min,其上清液即为200 g·L-1的母液,分别为新鲜凋落物浸提液(标记为FL)、自然凋落物浸提液(标记为NL);最后,均放入冰箱-18 ℃中保存。
1.2.2 种子萌发和生长试验 对FL、NL均设置8种浓度:200、100、50、20、10、5、2、1 g·L-1,以蒸馏水为对照(CK),共17个处理,每处理3次重复。取籽粒饱满的水杉种子均匀排列于铺有2层滤纸的培养皿(Φ 9 cm,使用前于180 ℃消毒2 h)中,每皿50粒,分别加入4 mL相应浸提液,置20 ℃(参考国家气象科学数据中心利川市四、五月的平均气温,为20 ℃)全黑暗(辛霞等,2004;郭秋菊等,2018)的SPX-30085H-Ⅱ生化培养箱,每3 d更换滤纸并添加4 mL的对应处理液以保证浸提液浓度。自水杉种子置床开始,每隔 24 h 观察记录发芽情况,以胚根出现为萌发标志。待种子发芽后在每个试验单元随机抽取10粒种子测量芽长,记录测量直至所有种子发芽完成,约20 d后测定其胚轴(子叶着生点到主根的一段距离)长度和主根(由胚根发育而成的根)长度(强胜,2005)。根据发芽情况计算种子发芽率、发芽势和发芽指数,根据种子发芽以及芽、根的生长情况计算化感效应敏感指数(allelopathy sensitivity index,RI)和化感综合效应指数(Synthetical effect of allelopathy index,SE),其计算公式如下。
式中:Ga为正常发芽的种子数;Gt为供试种子数。
式中:Gmax为发芽数最多日的发芽种子数;Gt为供试种子数。
式中:Gp为与Gd相对应的每天发芽种子数;Gd为发芽天数。
(4) 化感效应敏感指数(RI)(Williamson & Richardson,1988):
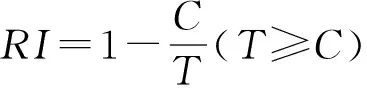
式中:C为对照值;T为处理值;RI>0为促进,RI<0为抑制,RI绝对值的大小与作用强度一致。
(5) 化感综合效应指数(SE)反映化感效应的强弱,是指同一处理下对同一受体各测试项目RI的算术平均值,即
SE=(发芽率RI+发芽势RI+发芽指数RI+芽长RI+胚轴RI+主根RI)/ 6。
1.2.3 数据处理 对不同浓度FL、NL处理下水杉种子萌发和生长进行方差分析及多重比较(Duncan);使用SPSS 18.0软件对不同处理下水杉种子芽长的生长变化进行Logistic模型拟合和计算生长参数(杨志玲等,2011;吴漫玲等,2020),即最大线性生长速率(maximum linear growth rate,MGR,即连日生长量最大时的生长速率)、线性生长速率(linear growth rate,LGR,即线性生长期内的平均生长速率)、线性生长量(total linear growth,TLG,即线性生长期内的生长量)和线性生长量占总线性生长量的百分率。具体计算公式如下。
Logistic模型拟合方程:
式中:y为芽长累计生长量;k表示拟合芽长的生长极限值;t为生长时间;a和b为待定系数。
2 结果与分析
2.1 凋落物浸提液对水杉种子萌发的影响
由图1可知,不同类型的凋落物浸提液处理后水杉种子萌发指标呈现一定差异性。不同浓度FL对种子发芽指数有显著影响(P<0.05),对发芽率和发芽势则影响不显著(P>0.05)。FL处理后,种子发芽率、发芽势和发芽指数随着浓度升高呈现波动变化且均在50 g·L-1处理时最高;而在1 g·L-1处理时发芽率和发芽指数显著比CK分别减少36.842%、32.677%。不同浓度NL对发芽率、发芽势和发芽指数影响不显著(P>0.05),在1 g·L-1处理下,发芽率、发芽势与发芽指数均高于CK,其中发芽势(36%)显著比10 g·L-1高2.349倍。总体来看,在FL浓度为50 g·L-1时对水杉种子萌发有一定促进作用,而在1、100、200 g·L-1时则有一定抑制作用;NL处理下,当浓度为1 g·L-1时显示一定促进作用,而5、10、200 g·L-1则抑制种子萌发。
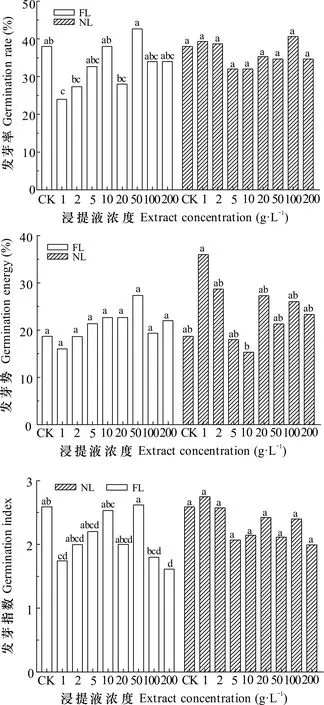
同一类型凋落物的不同小写字母表示差异显著 (P<0.05)。下同。Different lowercase letters of the same type of litter indicate significant differences (P<0.05). The same below.图 1 水杉凋落物浸提液对自身种子萌发的影响Fig. 1 Influence of Metasequoia glyptostroboides litter extract on its own seed germination
2.2 凋落物浸提液对水杉种子生长的影响
2.2.1 凋落物浸提液对3个种子生长指标的影响 由图2可知,不同浓度FL对芽长、胚轴长和主根长影响极显著(P<0.01),并且所有经过FL处理的种子生长的3个指标值均小于CK。当浓度高于10 g·L-1时,随着浓度的升高,各指标值下降程度越大;当低于此浓度时,3个种子生长指标值呈现波动变化。不同浓度NL对芽长和主根长有极显著影响(P<0.01),对胚轴长影响显著(P<0.05),并且这3个种子生长指标均在CK处理时为最大值(45.000、11.767、28.867 mm),其次为10 g·L-1,并且浓度高于10 g·L-1时,各生长指标值随浓度升高逐步降低;在1~5 g·L-1范围内,芽长和胚轴长呈现逐渐降低的趋势,主根长则表现为先降低再升高,这表明随着浸提液浓度的增加,种子生长表现出一定的浓度梯度抑制作用。经过FL和NL处理后,不同类型不同浓度的凋落物浸提液均对水杉种子萌发后的生长呈现出一定的抑制作用,当浓度高于20 g·L-1时,其抑制程度尤为显著。

图 2 水杉凋落物浸提液对自身种子生长的影响Fig. 2 Influence of Metasequoia glyptostroboides litter extract on its own seed growth
2.2.2 凋落物浸提液对芽生长进程的影响 水杉种子的芽长在试验期间的生长表现为开始较为缓慢,之后较迅速,达到一定值后,又趋于缓慢,直至生长停止。可见,芽长的生长变化符合“S”型生长曲线特点,故采用Logistic模型拟合。由表1可知,FL和NL处理下种子芽长Logistic拟合方程的决定系数为0.988~0.998,均达到了极显著相关水平。随着2种凋落物浸提液浓度的增加,芽长的生长曲线越平缓,芽长进入线性生长期的时间越晚(图3)。当浸提液浓度达到100、200 g·L-1时,FL处理下芽长的生长曲线明显低于NL处理下的生长曲线,说明高浓度下FL对芽长生长进程的抑制作用更明显。由表2可知,FL和NL处理后,水杉种子芽长MGR和LGR均随着浓度的增加逐渐下降,1~5 g·L-1时高于CK,而当浓度增加到50 g·L-1及以上时,MGR和LGR均小于CK;TLG均小于CK,除2、5 g·L-1处理外,其余处理均随着浓度的增加而逐渐降低;线性生长量占总生长量的百分率为55.263%~64.683%。
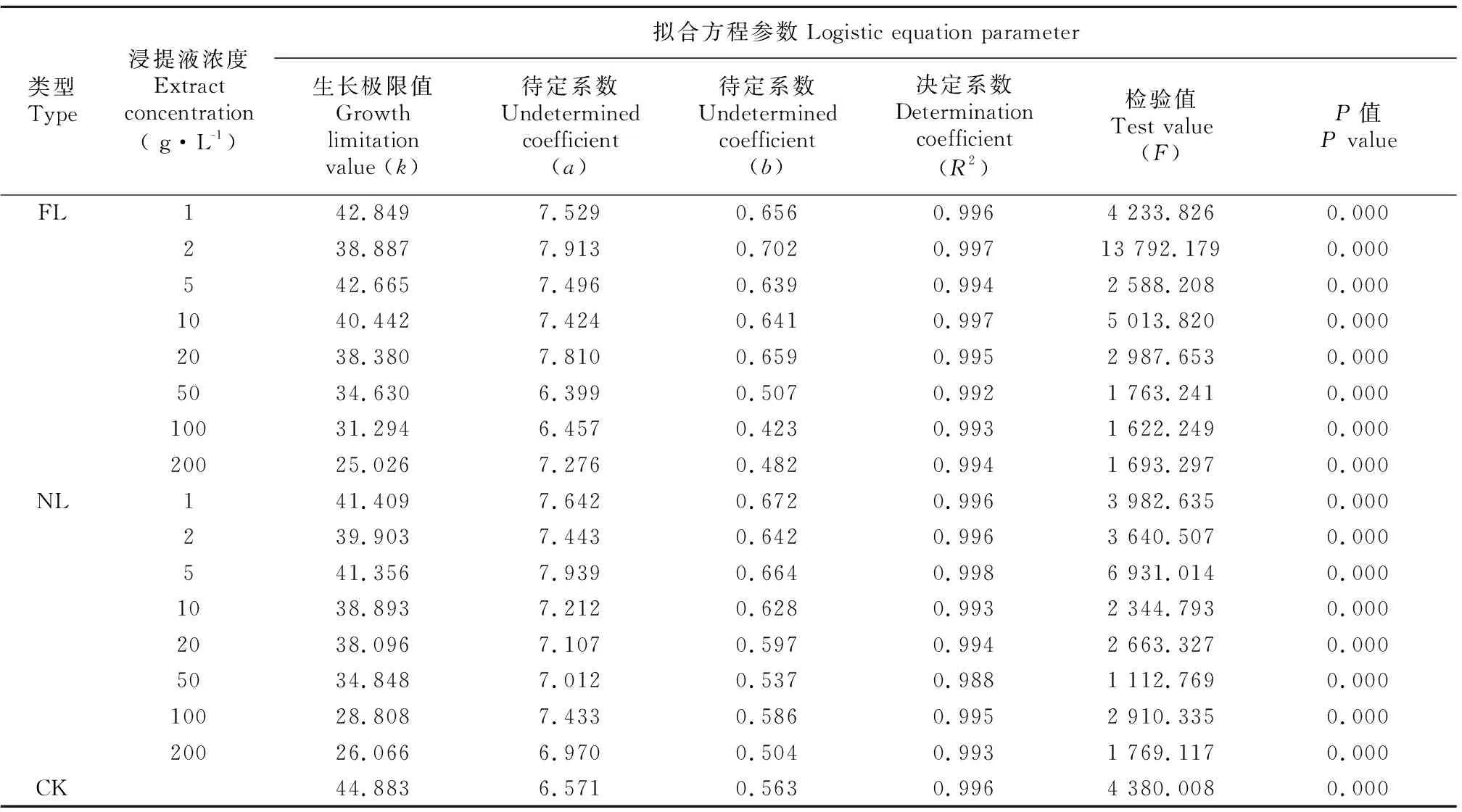
表 1 不同浓度凋落物浸提液处理水杉种子芽长生长曲线方程的拟合参数Table 1 Fitting parameters of shoot length growth curves equation of Metasequoia glyptostroboides seeds treated with different concentrations of litter extract
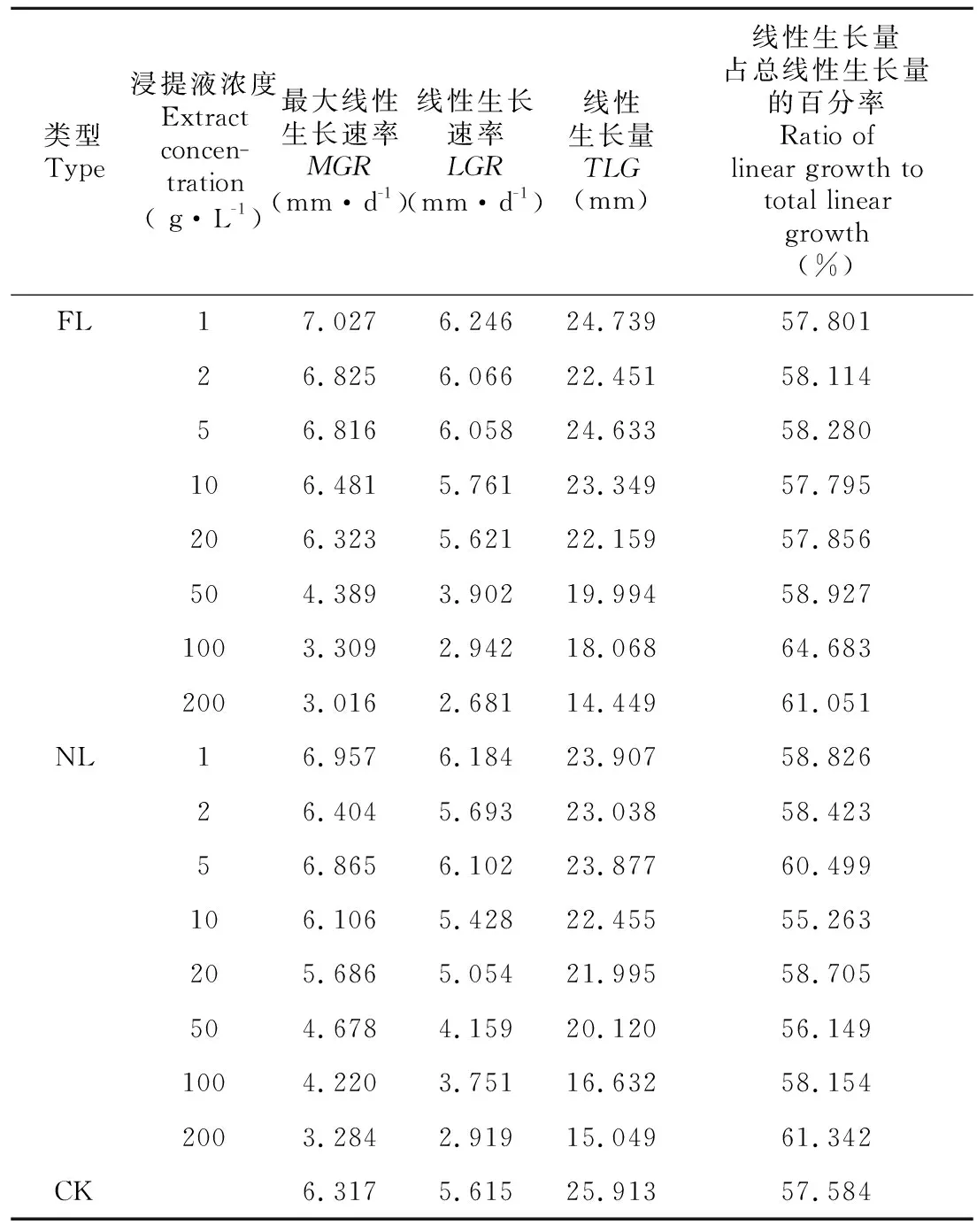
表 2 不同浓度凋落物浸提液处理水杉种子芽长的生长参数Table 2 Growth parameters of shoot length of Metasequoia glyptostroboides seeds treated with different concentrations of litter extracts
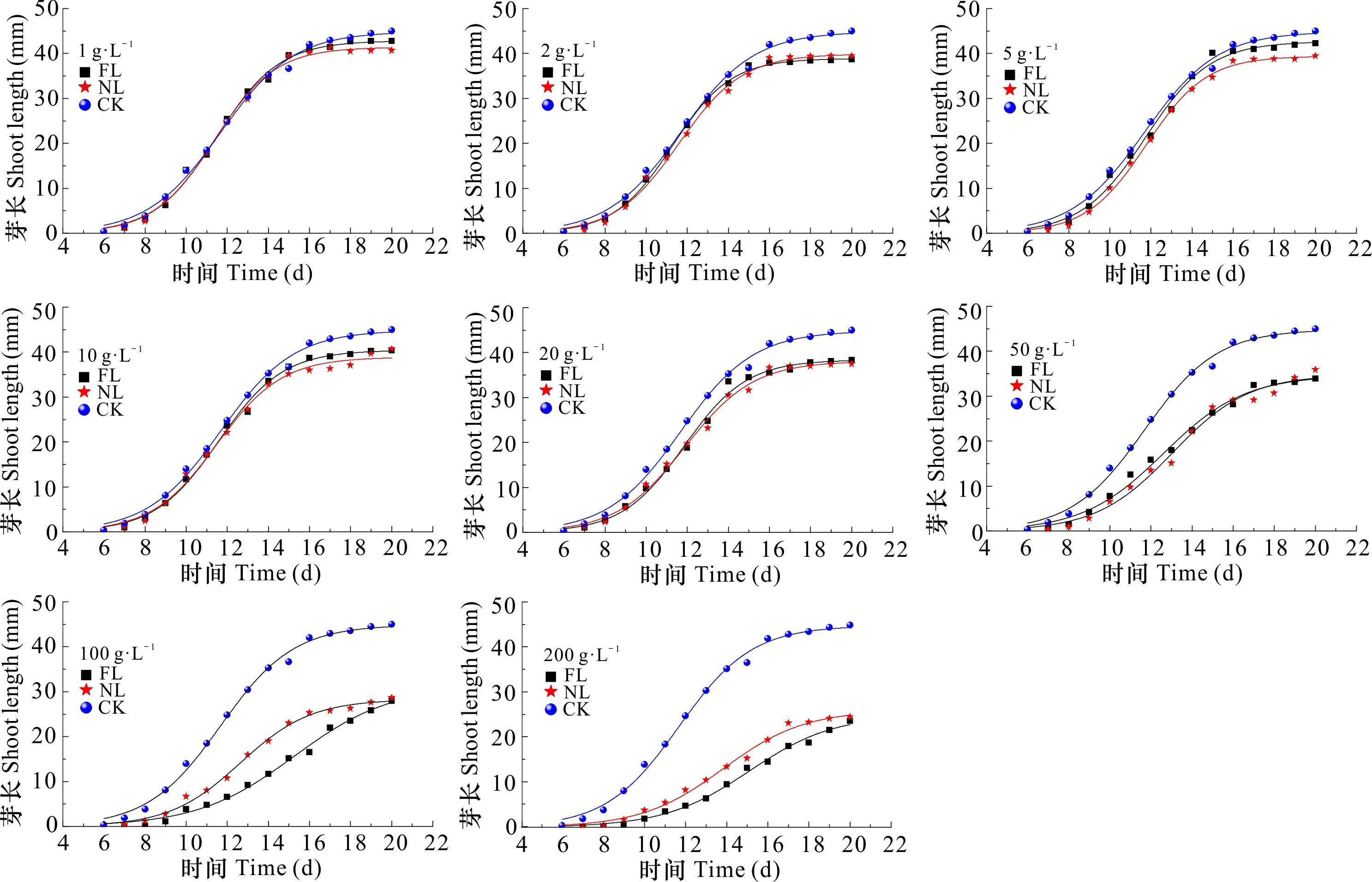
图 3 不同浓度凋落物浸提液对水杉种子芽长生长曲线的影响Fig. 3 Effects of different concentrations of litter extracts on shoot length growth curves of Metasequoia glyptostroboides seeds
2.3 凋落物浸提液对水杉的化感效应评价
由表3可知,FL处理后,种子萌发指标的化感效应敏感指数(RI)在1~5 g·L-1和100~200 g·L-1处理时为负数,即表现为抑制作用,其中1 g·L-1对种子发芽率和发芽势的抑制最强,200 g·L-1对发芽指数抑制最强,而10~50 g·L-1有不显著的促进作用。NL在1、2 g·L-1浓度处理时,种子萌发指标的RI为正值,即有一定促进作用,在此之后,随着浓度的升高,表现为无显著促进作用和抑制作用。FL和NL处理对水杉种子芽长、胚轴长和主根长均有抑制作用,在浓度≥10 g·L-1时,浓度越高抑制作用越强,同时对主根长的抑制作用高于胚轴长和芽长。
为综合分析凋落物浸提液对水杉种子萌发和生长的化感效应,进行化感效应综合指数(SE)计算,结果见表3。FL各浓度均抑制种子萌发和生长(-0.001~0.562),抑制作用表现为1~10 g·L-1范围内逐渐下降,10~50 g·L-1则为先上升再下降的趋势,100~200 g·L-1抑制最强;NL对水杉种子萌发和生长的影响为“低促高抑”的双重效应,1 g·L-1时为一定促进作用(0.044),浓度高于1 g·L-1时为抑制作用,并且浓度越高抑制越强。综合来看,FL对水杉种子萌发和生长的抑制强度大于NL,并且不同浓度呈现出不同的抑制效果。
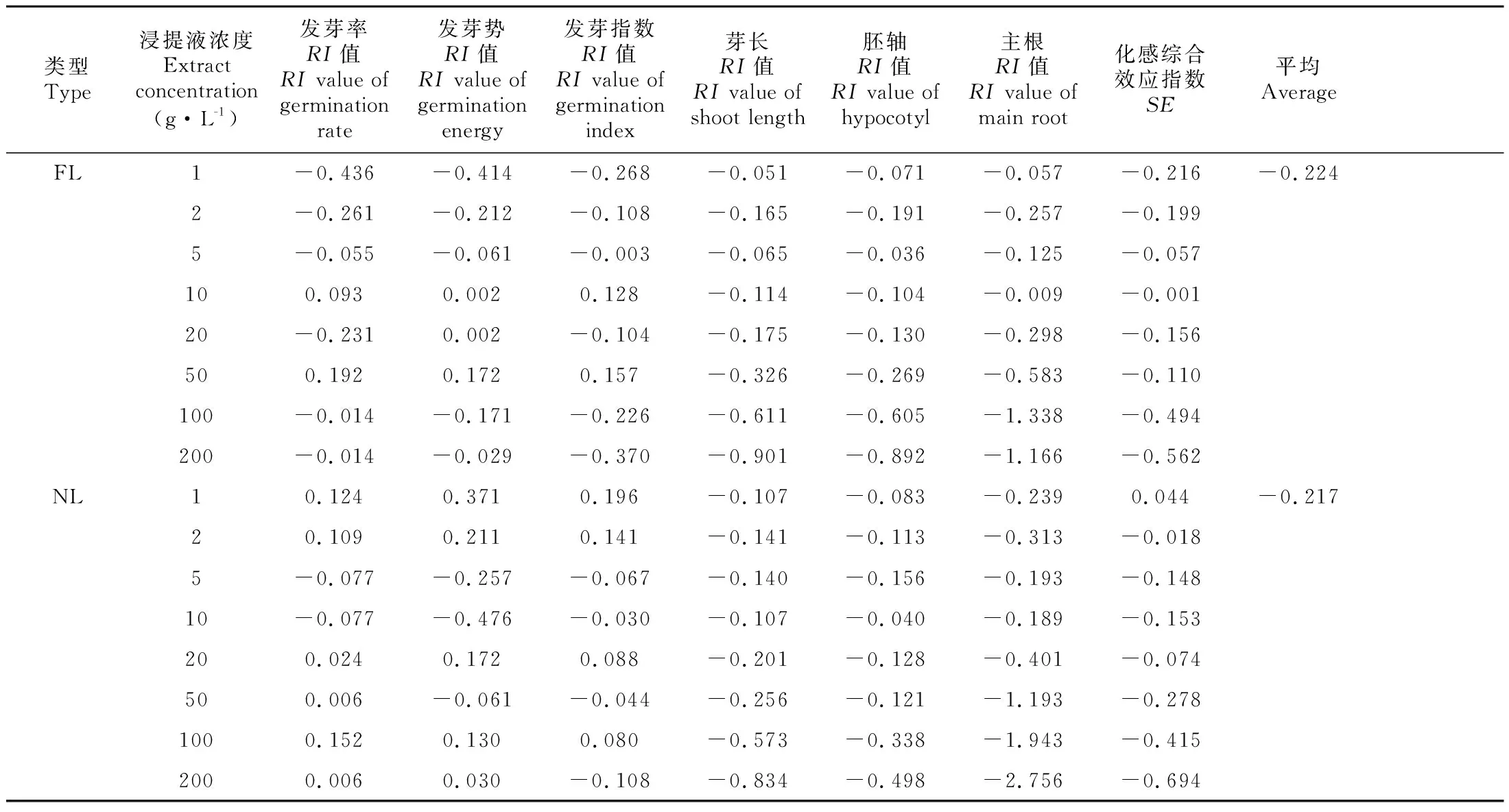
表 3 水杉凋落物浸提液对自身种子萌发和生长的化感效应敏感指数和化感综合效应指数的影响Table 3 Allelopathy sensitivity index and synthetical effect of allelopathy index of Metasequoia glyptostroboides litter extracts on its own seed germination and growth
3 讨论与结论
种子萌发是植物生活史的重要阶段,并且显著地受到凋落物的化感效应影响,从而影响种群的建立和更新(刘芳黎等,2017)。本研究证实水杉凋落物对水杉种子的萌发与生长有一定程度的抑制作用, 说明水杉母树凋落物对该种群的天然更新存在一定化感效应。新鲜凋落物浸提液在极低浓度(1 g·L-1)时阻碍种子发芽,这与Macias(1995)的研究一致,可能是新鲜凋落物中含有萘类、酯类、醇类等化感物质,干扰种子内部物质的代谢,抑制细胞分裂、伸长及膜渗透性等,进而阻碍种子萌发,因此表现出低浓度对种子萌发具有强烈抑制作用。但是,究竟是哪种化感物质起作用还有待进一步研究。高浓度(100、200 g·L-1)处理的新鲜凋落物浸提液对水杉种子萌发和生长均有一定抑制作用,这与晋梦然等(2020)研究格氏栲(Castanopsiskawakami)未分解层浸提液在高浓度时抑制杉木(Cunninghamialanceolata)种子发芽的结果一致。出现这种现象的原因是,随着化感物质在高浓度下积累到一定程度,会导致细胞器受到破坏(Mahdavikia et al., 2017),从而影响植物对水分和矿物质吸收利用,致使种子萌发和生长受到限制。水杉自然凋落物浸提液对水杉种子发芽率、发芽势和发芽指数影响不显著,除了浓度为1 g·L-1时呈现一定的促进作用外,其余均为抑制作用,并且在试验中发现高浓度的自然凋落物浸提液会延迟种子萌发。Garnett等(2004)研究发现新泽西州刚松、美洲越橘(Gaylussaciabacatta)和美国白栎(Quercusalba)3种主要树种的凋落物均对新泽西州刚松的种子发芽无影响,这主要是因为自然凋落物受土壤中微生物、风化、淋溶等作用的影响,导致其化感物质活性有一定降低(Araniti et al., 2016),化感作用也随之减弱。
幼苗阶段在植物整个生活史中至关重要,而芽是从种子到早期幼苗建成的最初阶段,其会受到众多因素的影响(蔺吉祥等,2011)。水杉新鲜和自然凋落物浸提液均抑制芽长、胚轴长和主根长,尤其是浓度高于20 g·L-1时极不利于种子萌发后的生长,这与毛红椿(Toonaciliatavar.pubescens)凋落叶水浸液对毛红椿幼苗生长抑制作用相同(郭晓燕等,2019)。植物凋落物释放的化感物质能够破坏内根组织、抑制根毛形成(Aguilera et al., 2017),降低对水分和营养物质的吸收,阻碍有机物质的迅速积累,从而导致水杉种子生长受到明显抑制,最终影响水杉幼苗在种群中的存活力和竞争能力,这在一定程度上阻碍了水杉种群天然更新。本研究发现,水杉凋落物浸提液浓度对主根长的抑制作用高于胚轴长和芽长的抑制作用,这也进一步证实了Chon等(2002)的研究结果,即苜蓿(Medicagosativa)根系比种子萌发率和胚轴长对化感自毒物质的反应更敏感。由于种子发芽后由主根直接向外界汲取生长所需能量,因此根是最先受到影响且影响程度比其他部分更加显著(张志忠等,2013)。
“S”型生长曲线描述某一种群受空间约束的生长过程(卢恩双等,2002),本研究发现,水杉种子芽长的生长动态变化也符合“S”型生长曲线(R2≥0.988),并且低浓度(≤20 g·L-1)凋落物浸提液在一定程度上促进芽的生长速率,而中高浓度则抑制芽的生长。这是由凋落物化感作用的浓度依赖性造成的,低浓度作用下诱导植物产生耐受蛋白、采取快速生长等方式以应对不良环境,当浓度超过植物自我调节能力的阈值时,其受到的化感抑制作用更明显 (Craine & Dybzinski, 2013)。
此外,芽长在试验生长周期内的线性生长量占总生长量的百分率在55%以上,进一步证实无论是在何种条件下线性生长期都是水杉生长过程中的关键时期(吴漫玲等,2020)。
化感效应敏感指数是衡量化感作用强度的重要指标,本研究发现新鲜和自然凋落物浸提液对种子萌发后生长的影响大于对种子萌发的影响。一方面是因为芽与化感物质直接接触,且接触的时间最长,化感抑制作用表现明显;另一方面,由于种子拥有种皮、种翅等保护作用,导致其对化感作用的响应有一定程度的滞后(Devaney et al., 2018)。不同类型不同浓度的凋落物浸提液对林木种子萌发和生长的影响不同,化感综合效应指数显示水杉新鲜凋落物浸提液对水杉种子萌发和生长的抑制强度大于自然凋落物浸提液,这与郭晓燕等(2018)和刘芳黎等(2017)的研究结果一致,均表明新鲜凋落物抑制作用大于自然凋落物,出现这种现象可能是凋落物在不同分解阶段产生的化感物质种类或含量不同,从而造成化感效应的差异化。本研究所有结果是在室内实验条件下得出,存在一定局限性,需要结合田间试验进一步探讨、验证水杉凋落物对水杉天然萌发与更新的影响机制。此外,天然更新受到复杂自然环境(温度、光照、水分、凋落物等)、人为因素(采种、干扰等)和物种自身特性(种子活力、种源、繁殖特性、遗传种群结构等)等多因素影响,如何有效促进水杉天然更新还需进一步综合研究。
种子萌发和早期生长是天然更新的重要阶段,本研究发现水杉新鲜和自然凋落物水浸提液均对自身种子萌发和生长产生了化感作用,并且总体表现为一定的抑制作用,验证了水杉凋落物的化感作用是影响水杉天然更新的障碍因素之一。由于水杉每年落叶时间刚好在球果成熟前后,因此今后在水杉母树保护与繁育管理中,建议在种子雨高峰期前适当清理水杉凋落物,以避免化感物质富积,为种子萌发和幼苗生长提供适宜环境,从而促进水杉种群天然更新。
猜你喜欢
山东林业科技(2021年4期)2021-08-31
江西农业大学学报(2021年3期)2021-07-22
福建林业科技(2020年3期)2020-10-25
现代园艺(2020年17期)2020-08-31
福建农林大学学报(自然科学版)(2020年4期)2020-07-15
现代园艺(2020年7期)2020-04-22
华南农业大学学报(2020年1期)2020-01-18
湖北农业科学(2019年20期)2019-12-11
西南农业学报(2019年9期)2019-11-11
热带作物学报(2019年1期)2019-06-11
