
引导编辑:突破碱基编辑类型的新技术
2022-11-29 09:15刘尧周先辉黄舒泓王小龙
遗传 2022年11期
刘尧,周先辉,黄舒泓,王小龙
综 述
引导编辑:突破碱基编辑类型的新技术
刘尧,周先辉,黄舒泓,王小龙
西北农林科技大学,动物科技学院/农业农村部动物生物育种国际联合研究中心/陕西省动物遗传育种与繁殖重点实验室,杨凌 712100
引导编辑技术(prime editing)是一种基于CRISPR/Cas系统的新型基因编辑技术。引导编辑器(prime editor, PE)的效应蛋白是逆转录酶与具有单链切割活性的nCas9(H840A)的融合蛋白,被称为PE2;其向导RNA是通过在sgRNA的3′末端增加逆转录模板和引物结合位点序列构成,被称为pegRNA。PE系统可以实现所有12种类型的碱基突变,还可以实现短的插入删除编辑,及多种编辑类型的组合。自2019年被开发以来,因其编辑类型多样化及高特异性等优势,PE已经被成功地应用在多种动植物及细菌中,同时在基因治疗和农业育种等领域也展现出良好的应用前景。本文对PE系统的开发过程、特点、优化、应用、安全风险等方面进行了系统介绍,并对其发展前景进行了展望,以期有助于相关领域研究人员对PE系统的了解与应用。
引导编辑器;CRISPR/Cas9;优化;应用
经过近10年的发展,基于CRISPR的基因编辑技术因其精准性和高效性已经被广泛应用于基因组的修饰[1~3]。CRISPR/Cas9系统通过在基因组上诱导双链断裂损伤(double strand break, DSB)来实现靶位点的编辑[1]。细胞修复DSB的方式主要有两种:非同源末端连接(non-homologous end joining, NHEJ)和同源重组(homology directed repair, HDR)[4]。细胞通过NHEJ修复DSB会随机产生碱基的插入或缺失,造成基因的移码突变,从而实现基因敲除[1]。细胞通过HDR修复时需要DNA片段作为修复模板,此时可以提供人为设计的外源DNA模板,从而在基因组上引入包括点突变、插入、删除等在内的基因修饰[1]。细胞倾向于利用NHEJ来修复DSB,造成HDR的效率偏低,因此CRISPR/Cas9系统无法高效地诱导点突变等编辑类型[4]。
为此,美国哈佛大学David R. Liu课题组于2016年首次开发出胞嘧啶碱基编辑器(cytosine base editor, CBE)[2]。他们将大鼠胞嘧啶脱氨酶、nCas9(D10A)和尿嘧啶糖基化酶抑制剂(uracil glycosylase inhibitor, UGI)进行融合表达,该融合蛋白可以在sgRNA的引导下靶向基因组目标位点,催化其解旋形成R-loop结构,从而暴露出单链DNA,胞嘧啶脱氨酶催化一定窗口内单链DNA上的胞嘧啶(C)脱氨形成尿嘧啶(U),随着DNA的复制或修复,最终转化为胸腺嘧啶(T)。基于类似的思路,David R. Liu课题组又于2017年开发了腺嘌呤碱基编辑器(adenosine base editor, ABE),这一工具可以完成腺嘌呤(A)向鸟嘌呤(G)的转换[3]。CBE和ABE可以实现碱基之间的转换,但还不能实现碱基间的颠换[2,3,5];同时,小片段的插入删除编辑也缺乏高效编辑工具,更为全面有效的基因编辑工具的开发成为必然。
2019年,引导编辑器(prime editor, PE)的出现将精准基因组编辑推到新的高度[6]。PE系统通过对靶位点的“搜索”和“替换”,可以实现4种碱基之间的任意变换及小片段的精准插入和删除,克服了CRISPR/Cas9系统介导的HDR效率低和碱基编辑系统不能实现碱基颠换的弊端,具有明显的优势[6,7]。PE系统目前已被广泛地应用到农业、基因治疗、疾病模型构建等多个研究领域[8~11]。2019年,发表PE系统的论文被杂志评为年度十大杰出论文。本文将对PE系统的开发过程、特点、优化、应用、安全风险等进行系统介绍,并对其发展前景进行展望,从而增进人们对PE系统的了解,促进其应用。
1 PE系统的开发及特点
1.1 PE系统的开发
PE系统的核心包括效应蛋白及pegRNA两部分[6]。效应蛋白是逆转录酶与具有单链切割活性的nCas9 (H840A)的融合蛋白,被命名为PE2蛋白[6]。pegRNA是在sgRNA的基础上,对其3′端进行延长,增加逆转录模板(reverse transcriptase templates, RTT)和引物结合位点(primer binding site, PBS)。RTT序列携带人为设计的遗传信息,而PBS序列是逆转录引物的结合位点[6]。引导编辑的核心原理可分为3步:第一步是PE2蛋白的靶向及切割,即PE2蛋白在pegRNA的引导下,靶向基因组目标位点,催化其解旋,形成R-loop结构,从而暴露出单链DNA,随后nCas9在PAM上游第3个碱基处切割DNA单链使DNA游离;第二步是逆转录反应,即游离的单链DNA与PBS碱基互补,起始逆转录反应,逆转录酶以RTT为模板,合成新的单链DNA[6];第三步是DNA的修复,即新合成的单链DNA形成3′-flap,与原有的5′-flap进行竞争,细胞中的DNA修复机制倾向切除5′-flap,随后,3′-flap在DNA连接酶的作用下,被整合进基因组,形成带有错配的DNA双链;内源DNA修复机制识别此错配并以被编辑链为模板,修复未编辑链,从而实现双链的编辑[6](图1)。
2019年,David R. Liu课题组首次建立了3个版本的PE系统[6]。第一代PE系统PE1是由野生型的M-MLV逆转录酶与具有单链切割活性的nCas9 (H840A)组成[6]。PE1对点突变的编辑效率最高可达到5.5%,对于短的插入删除的编辑效率在4%~17%之间[6]。为了提高PE系统的编辑效率,David R. Liu课题组在野生型逆转录酶上引入了5处突变来提高逆转录酶的热稳定性、反应的持续性、与DNA:RNA底物的亲和性及抑制逆转录酶的RNaseH结构域的活性,开发了第二代PE系统PE2[6]。相比于PE1,PE2将点突变的编辑编辑效率提高了1.6~5.1倍,同时对于短的插入删除编辑也有了非常大的提高[6]。在PE2介导的基因编辑过程中,会存在一条链被编辑,一条链未被编辑的异源双链状态。在以往的碱基编辑系统开发过程中,为了提高编辑效率,David R. Liu课题组利用Cas9的单链切割活性在未编辑链上引入一个切口来促使DNA修复机制以编辑链为模板修复未编辑链,从而实现DNA的双链编辑。基于此思路,David R. Liu课题组通过额外添加一条sgRNA来引入此切口,并将此三元系统命名为PE3[6]。如果额外添加的sgRNA在作用过程中其spacer序列与被编辑后的序列互补,则称该系统为PE3b[6],PE3和PE3b共同构成了第三代PE系统。由于成功被编辑的链才能被sgRNA靶向,所以PE3b可以降低在DNA链上同时出现两个切口的概率,从而减少DSB的形成和indels的产生。与PE2相比,PE3和PE3b将编辑效率提高了约3倍[6]。
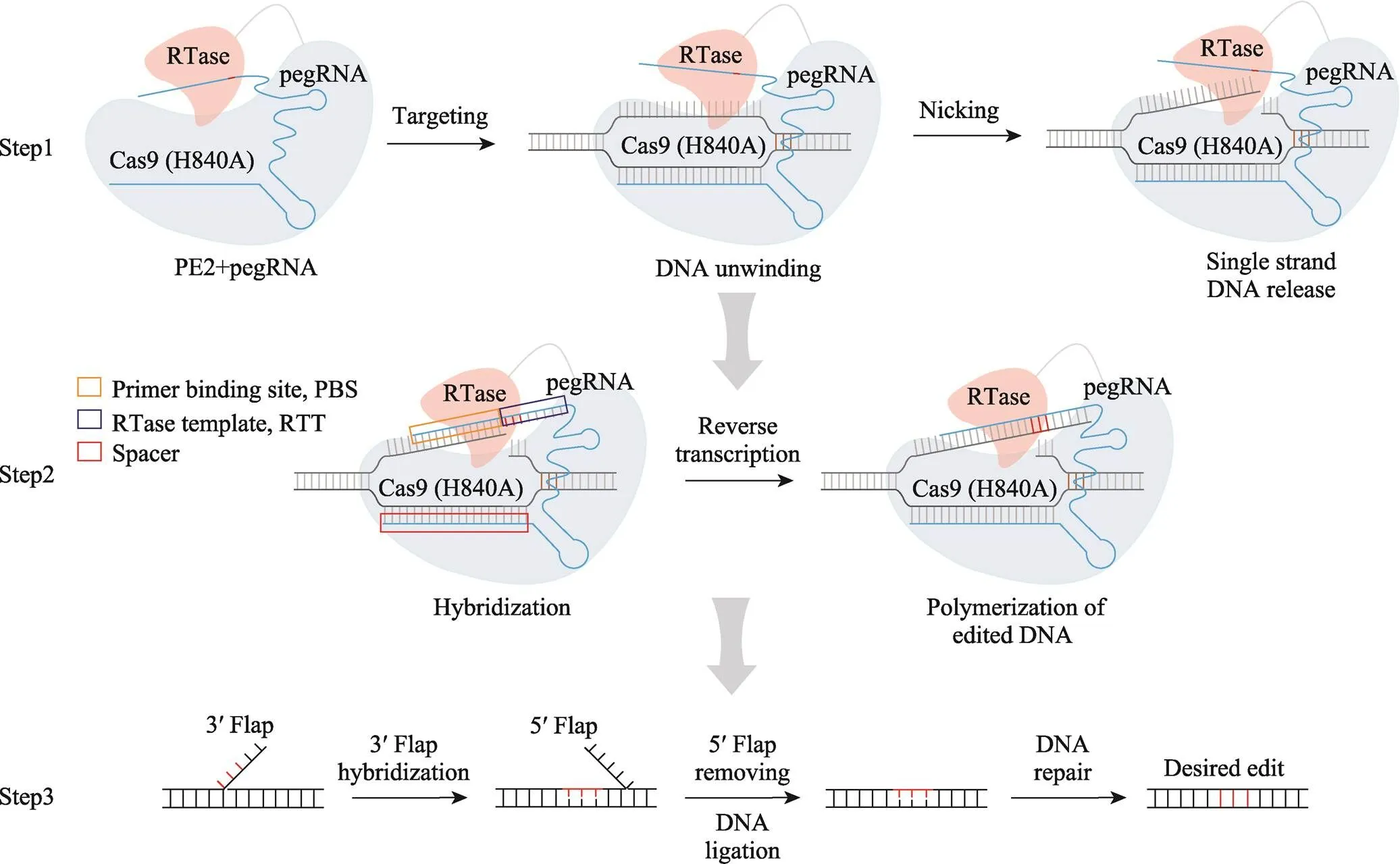
图1 PE工作原理图
PE系统主要通过3步在基因组靶位点引入基因修饰:PE2蛋白的靶向及切割、逆转录反应和DNA的修复。
1.2 PE系统的特点
PE系统相比原有的基于CRISPR/Cas9的基因编辑工具优势十分明显,其主要特点可归结如下:
(1)编辑类型的多样化。PE系统可以实现所有12种类型的碱基突变,还可以实现小片段插入删除编辑,及多种编辑类型的组合,研究人员可以根据目标突变设计相应的pegRNA来进行靶位点的编辑[6]。
(2)编辑的高精准性。相较于CRISPR/Cas9系统通过HDR途径引入点突变和插入删除,PE系统不依赖DSB的产生,避免了过多indels产物。另外碱基编辑系统在作用过程中,常常会引起邻近非靶标碱基的编辑,而PE系统通过逆转录生成目标突变,避免了这种副产物的产生。对于PE3来说,虽然另一条sgRNA的引入造成了indels产物的增加,但indels的水平大多在10%以下,仍然保持了较高的精准性[6]。
(3)编辑的高特异性。以往的CRISPR/Cas9系统和碱基编辑系统均存在sgRNA依赖性脱靶,此外碱基编辑系统还存在sgRNA非依赖性脱靶和RNA脱靶,造成了很大的安全风险。PE系统在基因组靶位点上引入目标编辑过程中,需要发生3种核酸杂交:靶向区域DNA与pegRNA的spacer的杂交、靶向区域DNA与pegRNA的PBS序列的杂交、靶向区域DNA与逆转录产物的杂交,以上3种杂交的存在决定了PE系统在理论上很难脱靶[6]。
2 PE系统的优化及其衍生技术
PE系统虽然优势明显,但其早期版本的编辑效率均较低,极大地限制了其应用。目前,已经有许多研究从pegRNA[12~17]、PE2蛋白[18~22]、靶向范围[23]、DNA修复通路[20,24,25]等方面对其进行优化,使其效率得到了极大提升(表1)。此外,还有研究在PE系统基础上加以改造,建立了新的具有特殊编辑能力的新系统[26~29]。
2.1 pegRNA的优化
pegRNA是PE系统的核心部分之一,其结构及稳定性对引导编辑效率有重要影响(图2)[6]。David R. Liu课题组发现pegRNA的PBS和RTT长度会影响引导编辑效率,他们建议对PBS及RTT长度进行测试以便于找到最优pegRNA[6]。本课题组发现pegRNA的PBS与spacer发生互补会导致pegRNA环化,进而影响Cas9的靶向效率,于是在pegRNA的3′末端添加RNA茎环结构来抑制此互补,显著提高了PE系统的编辑效率;随后结合Csy4蛋白RNA加工系统,本课题组开发了增强型PE系统ePE,最高可平均提高4.9倍的编辑效率[12]。Nelson等[15]发现pegRNA 3′末端的降解会降低PE系统的活性,他们将结构化RNA基序融合到pegRNA的3′末端,以防止3′末端的降解增强其稳定性,由此产生了工程化pegRNA (epegRNA);在多种细胞系中,epegRNA可将引导编辑效率提高3~4倍。类似地,本课题组[13]和Li等[14]分别将具有稳定二级结构的xrRNA和G-四链体融合到pegRNA的3′末端构建了xrRNA-pegRNA和G-四链体修饰pegRNA,基于这两种pegRNA的PE系统分别被命名为xrPE和G-PE;xrPE和G-PE均有效提高了引导编辑效率,且效果与epegRNA相当。Li等[30]分别通过在pegRNA的RTT上引入同义突变和改变pegRNA二级结构构建了spegRNA和apegRNA,二者将PE效率分别平均提高353倍和2.77倍。在植物上,Jiang等[16]通过增强pegRNA的表达在玉米上实现了比在水稻中更高的编辑效率。此外,有研究指出在合成的pegRNA上增加化学修饰可能会提高pegRNA的稳定性,从而提高引导编辑效率[31]。
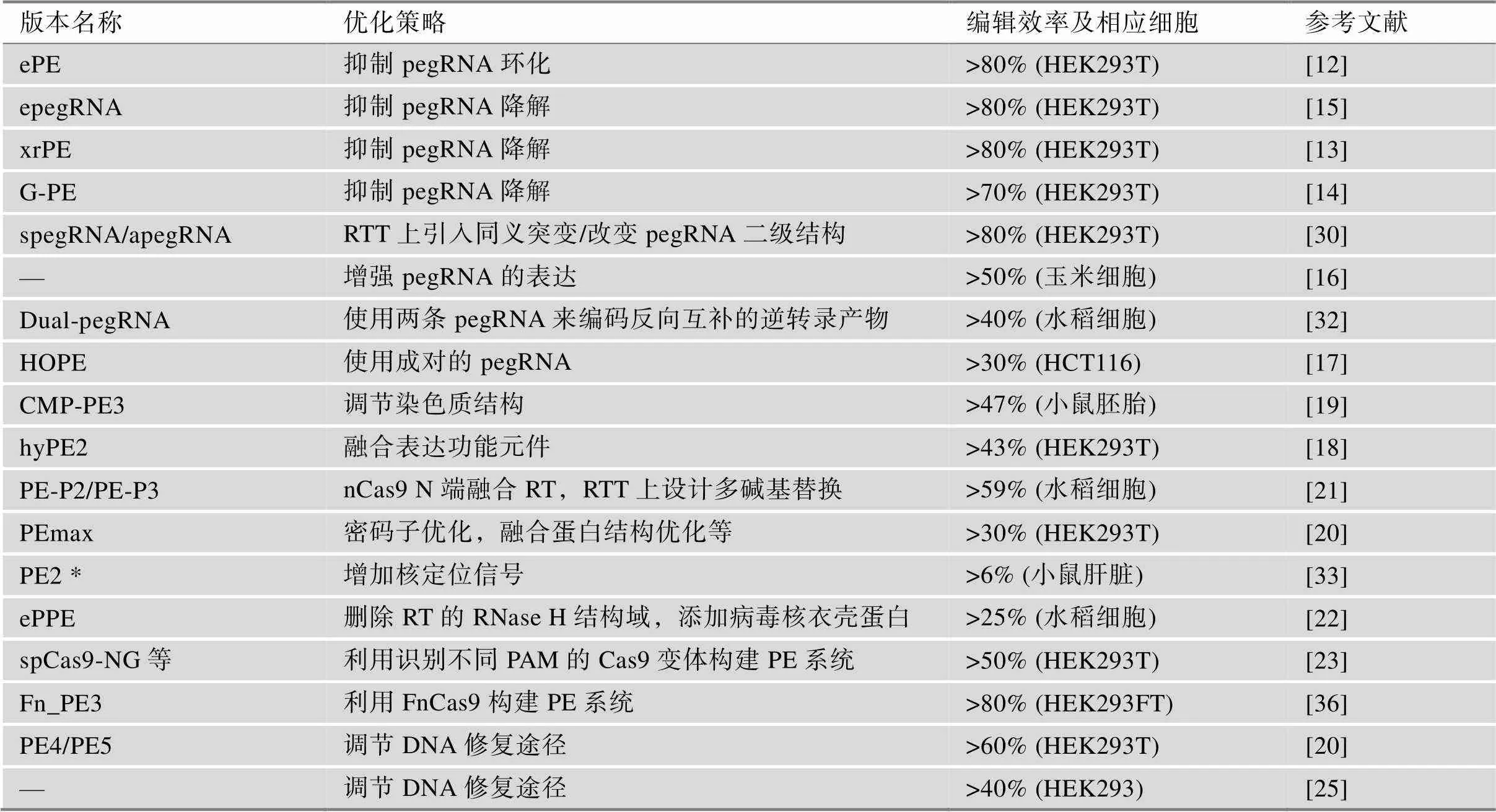
表1 各种优化版本的引导编辑器

图2 不同pegRNA修饰示意图
图中展示6种对pegRNA的优化方式,spacer、scaffold、RTT和PBS表示pegRNA的组成部分,Csy4 recongnition site、xrRNA、G-quadruplexes和pseudoknot表示具体特殊二级结构的RNA序列,SSMs表示RTT编码的DNA为同义突变。
PE3系统需要另一条sgRNA来在非编辑链引入切口,研究人员通过使用另一条pegRNA来替代这条sgRNA,即双pegRNA策略。其中,Lin等[32]通过优化pegRNA的PBS的解链温度及使用两条pegRNA来编码反向互补的逆转录产物在水稻中引入目标突变,将编辑效率最高提升了17.4倍。Zhuang等[17]使用配对的pegRNA开发了HOPE系统,HOPE显示出比PE2和PE3系统更高的编辑效率,并且具有比PE3更高的编辑产物纯度。
2.2 效应蛋白的优化
效应蛋白是PE系统的另一个核心组成部分,对其进行精心设计改良是优化PE系统的另一种途径,且优化的方向主要有3个方面。一是融合表达相关功能元件;将PE2蛋白分别与染色质调节肽(CMPs)、Rad51蛋白的DNA结合结构域、c-Myc NLS融合表达构建的PE2蛋白变体均可有效提高引导编辑效率[18,19,33];此外,Zong等[22]通过去除逆转录酶的RNase H结构域并添加具有核酸伴侣活性的病毒核衣壳蛋白来优化逆转录酶,将引导编辑效率平均提高了5.8倍,且不会增加副产物和脱靶编辑。二是对效应蛋白序列和结构的优化;David R. Liu课题组通过对逆转录酶进行人类密码子优化、在SpCas9上引入两个可以提高其活性的R221K和N394K突变、在C端添加c-Myc NLS序列和将nCas9和RT之间连接肽中的XTEN换成SV40 NLS得到了具有高度普适性的PEmax蛋白,经过测试,PEmax显著优于已报道的包括利用c-Myc NLS构建的PE2*在内的其他变体,后续对效应蛋白的进一步改进均可在PEmax的基础上进行[20];Xu等[21]发现在nCas9 (H840A)的N端融合逆转录酶比现有的C端融合具有更高的编辑效率,于是他们使用N端融合逆转录酶的PE2蛋白结合含有多个碱基替换的逆转录模板在水稻和玉米上分别实现了最高达24.3%和6.2%的编辑[21]。三是对效应蛋白尺寸的优化;PE2蛋白较大的尺寸会降低其递送的效率,在两项研究中,Liu等[34]和Zheng等[35]分别采取将nCas9(H840A)和逆转录酶分开表达和去除逆转录酶的RNase H结构域的策略来减小蛋白尺寸,且未造成明显的编辑效率损失。
2.3 拓展引导编辑的范围
PE系统最初是基于识别NGG PAM的SpCas9进行开发的,虽然PE系统的编辑窗口决定了其对PAM的依赖性较低,但仍有部分位点缺乏设计pegRNA所需的PAM[6]。利用可以识别不同PAM的Cas9的变体或者同系物如SpCas9-VQR、SpCas9- VRQR、SpCas9-NG、SpCas9-SpRY、SaCas9等构建PE,是一种很好的解决策略[23,33]。此外,Oh等[36]利用SpCas9的同系物FnCas9构建了新的PE;SpCas9是在PAM上游3~4 bp之间对非靶向链进行切割,而FnCas9的切割位置在PAM上游6~8 bp之间,且FnCas9和SpCas9识别同样的NGG PAM,因此基于FnCas9的PE系统具有更大的编辑窗口,扩大了PE的编辑范围。
2.4 调控DNA修复相关通路
PE系统在基因组上引入目标突变需要细胞内源DNA损伤修复机制的参与,通过对影响编辑效率及编辑产物纯度的关键DNA修复通路进行调控有助于改善PE系统的性能[20]。David R. Liu课题组利用CRISPRi筛选确定了错配修复通路(mismatch repair,MMR)对引导编辑的效率和产物纯度的影响;他们发现,敲低、、、这4种MutSα–MutLα错配修复复合体组成蛋白的编码基因,PE2的编辑效率可提高5.8倍,PE3的编辑效率可提高2.5倍;此外,敲低(MMR通路中的核酸外切酶)也会将PE2编辑效率提高2.3倍;随后他们利用失去功能活性的MLH1蛋白变体(MLH1dn)来抑制MMR,并将共表达MLH1dn的PE2和PE3分别命名为PE4和PE5;与PE2相比,PE4的编辑效率提高了2.0倍,indels频率从0.40%降低到0.31%;与PE3相比,PE5的编辑平均提高了1.2倍,编辑产物纯度提高了2.8倍[20]。无独有偶,Silva等[25]通过对涵盖了所有已知的修复途径的32种DNA修复因子进行遗传筛选,发现在、、、和这些MMR通路的关键基因缺失的单倍体HAP1细胞系中,PE效率提高了2~6.8倍,表明MMR对PE的抑制作用;通过使用siRNA特异性降解细胞中MLH1 mRNA,可以将PE效率提高约2倍。
2.5 引导编辑的衍生技术
引导编辑技术以其高度的灵活性得到了广泛的应用,并且基于其衍生出了多种针对特定编辑类型的编辑系统(表2)。其中,Anzalone等[37]使用两条相向的pegRNA来逆转录出互补的DNA链进而替换两个DNA切口之间的内源性DNA序列,此系统被命名为TwinPE;TwinPE可以实现基因组中大片段的替换,插入或删除,极大地提高了PE系统的精准基因编辑能力[37]。类似地,Choi等[38]同样使用一对相向的pegRNA开发了PRIME-Del系统,PRIME-Del可以精准删除长达10 kb的DNA片段,编辑效率介于1%~30%;Wang等[28]使用一对pegRNA编码与靶位点非同源但彼此互补的DNA单链,开发出了GRAND系统,GRAND诱导150 bp插入的效率高达63.0%,诱导250 bp插入的效率为28.4%,最高可以实现约1 kb的插入。
PE系统中的nCas9(H840A)具有单链切割活性,最近有多项研究表明,具有双链切割活性的Cas9同样可以介导引导编辑的进行,并且具有一些独特的优势。Peterka等[24]将逆转录酶与和野生型SpCas9进行融合构建了PEn蛋白;利用常规的pegRNA,PEn可以通过同源依赖性DSB修复机制实现片段插入;利用在3′末端仅包含PBS和目标插入的pegRNA,PEn可以通过精确的NHEJ实现小片段插入编辑,效率高达50%。Jiang等[29]将双链切割活性的Cas9与逆转录酶融合,并结合两条靶向互补DNA链pegRNA,开发出了PEDAR系统,它可以高效地诱导20~700 bp的片段删除和长达30 bp的片段插入。Adikusuma等[26]同样利用野生型SpCas9构建了新的PE系统PEA1,并且将蛋白、pegRNA和nicking sgRNA整合在同一个质粒中进行表达,该系统在HEK293T细胞中平均编辑效率可达到67%。此外,将此系统注射小鼠胚胎,在新生小鼠中最高可以实现100%的编辑效率。Tao等[27]将逆转录酶与野生型spCas9融合构建了WT-PE系统,通过使用分别靶向基因组上的不同区域两条pegRNA来逆转录出互补的DNA单链,WT-PE可以实现高效、多功能的大规模基因组编辑,包括长达16.8 Mb长的大片段删除和染色体易位,为模拟或治疗与大片段异常相关的疾病的提供了工具。
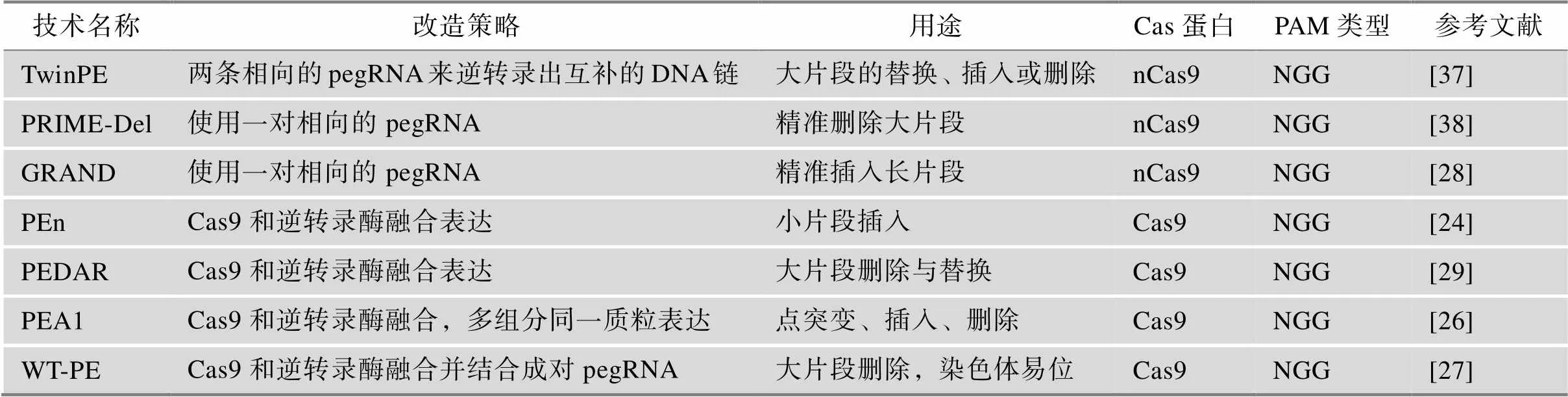
表2 引导编辑的衍生技术
3 PE系统的应用
PE系统突破了碱基编辑系统编辑类型和CRISPR/ Cas9系统副产物的限制,极具应用潜力。自其诞生以来,被迅速应用到动物如小鼠()、果蝇()、斑马鱼()、兔子(),植物如小麦(L.)、水稻(L.)、玉米(L.)、番茄(Mill.)、烟草()、小立碗藓()、马铃薯(L.)、拟南芥(),还有大肠杆菌()及各种细胞模型中[9,10,16,31,39~47];此外,其在基因治疗、遗传突变筛选等方面也多有应用[35,48~50](图3)。
3.1 在动物中的应用
建立动物疾病模型是研究人类疾病的发病机制及开发相关药物的重要手段,基因编辑技术的发展为建立人类遗传疾病的动物模型提供了强大工具。利用CRISPR/Cas9系统和碱基编辑系统,研究人员已经建立一大批动物疾病模型,极大的促进了相关疾病的研究[51,52]。然而,通过CRISPR/Cas9系统介导的HDR建立疾病模型效率低下,碱基编辑系统也存在bystander编辑和编辑范围受限的问题;PE系统的出现,很大程度上弥补了这些弊端。小鼠是最常见的模型动物,基于引导编辑技术的多种小鼠疾病模型已经被成功创制。其中,本课题组通过显微注射PE2 mRNA、pegRNA和nicking sgRNA编辑小鼠胚胎基因,首次成功利用PE系统在小鼠上诱导人类同源致病突变[10]。Gao等[53]通过显微注射PE2 mRNA和pegRNA成功地在小鼠基因的启动子区域引入C>G碱基颠换,26%的子代小鼠携带目标突变同时表现出相应的疾病特征;通过检测发现,PE系统在被编辑小鼠中保持高保真性和高特异性。Park等[19]以RNA的形式注射优化后PE系统到小鼠胚胎,同时将G>C的单碱基突变和TA两碱基插入引入小鼠基因,阳性子代小鼠体内的编辑效率最高可达47%;获得的基因突变的后代小鼠成功地表现出矮小的表型。Lin等[9]通过向小鼠胚胎注射质粒利用PE系统成功编辑小鼠基因获得了白内障小鼠模型,子代小鼠中最高编辑效率为41%,并且表现出明显的白内障症状。Petri等[31]以RNP的形式将PE系统注射到斑马鱼胚胎,成功将人类眼部皮肤白化病致病突变和癌基因突变引入斑马鱼,成功诱导了可遗传突变,这也证明了利用RNP递送PE系统的可行性。Qian等[40]同样使用显微注射的方法利用PE系统编辑兔的基因,在其第11外显子插入4个碱基TATC,成功构建了人泰萨克斯病(Tay-Sachs disease,TSD)兔模型。除了小鼠外,Bosch等[39]通过构建体内表达PE2蛋白和pegRNA的转基因果蝇,在果蝇上也成功进行了引导编辑,证明了PE系统其他动物物种中的可行性。

图3 PE系统的应用
A:在动物中的应用;B:在基因治疗中的应用;C:在植物中的应用;D:在突变筛选中的应用。
基因编辑技术在动物上的另一项重要应用是进行基因编辑育种。利用基因编辑技术来修饰影响经济性状的关键功能基因,从而获得具备良好生产性能的优质种质资源,突破了传统家畜育种周期长、遗传改良效率低的弊端。目前,研究人员已经利用CRISPP/ Cas9系统和碱基编辑系统创制了包括山羊、绵羊、猪、兔、奶牛等大批具有优异经济性状的种质资源[54~60]。PE系统的出现,为动物基因编辑育种提供了更加强大的工具,可以在动物基因组上有效引入更加多样的修饰类型。目前,PE系统还未有在大动物上应用的报道。本课题组前期在绵羊上对PE系统在大动物上的有效性进行了相关探索,利用经典版本的PE系统PE3编辑绵羊和两个基因,虽然在子代羔羊中检测到了目标突变,但个体中最高编辑频率仅有14.7%,这说明PE系统的效率还需要进一步优化。
3.2 在基因治疗中的应用
基因治疗是基因编辑工具的一项重要应用,使用基因编辑工具直接修复致病突变有望治愈人类遗传疾病[61]。CRISPR/Cas9系统和碱基编辑系统出现以后,研究人员已经进行了许多基因治疗方面的探索,其中进展最快的转甲状腺素蛋白淀粉样变性的CRISPR疗法已经进入临床试验阶段并取得了优异的治疗效果[62]。
引导编辑技术高精准性及高特异性的特点使其在基因治疗方面更有潜力。目前研究人员已经在细胞及动物个体水平进行了大量的研究。Schene等[63]利用PE3系统成功纠正了来源于患者的肠道类器官基因的3 bp缺失突变(c.629_631delCCT,p.S210del)及肝脏类器官中基因的1 bp重复突变。Chemello等[64]利用PE3系统成功在人诱导性多能干细胞的基因第52外显子插入AC两个碱基,恢复细胞中的抗肌萎缩蛋白表达。Kim等[65]将来自遗传性酪氨酸血症1型(HT1)小鼠模型中的肝细胞重编程肝脏祖细胞(CdHs),并利用PE系统成功纠正基因上的致病突变,编辑效率达到2.3%。Geurts等[43]利用PE3系统成功修复了肠道类器官基因的F508del和R785*突变,但编辑效率较低。在动物成体治疗方面,Jang等[33]采用流体动力学注射的方式递送质粒编码的PE3系统到1型遗传性酪氨酸血症的小鼠模型体内,通过修复基因上G>A致病突变,改善了小鼠的疾病表型[11]。Liu等[33]采用流体动力学注射的方式以质粒的形式递送改进后的PE系统到成体小鼠的肝细胞来修复基因上G>A的致病突变,平均编辑效率达到6.7%。Böckd等[50]使用人腺病毒5(AdV5)载体递送PE系统到苯丙酮尿症(PKU)小鼠模型的肝脏细胞,在新生小鼠体内修复致病突变的效率可达5.6%,在成年小鼠体内可达2%。Jiang等[29]基于双pegRNA策略开发了PEDAR,并采用流体动力学注射的方法递送表达PEDAR系统的质粒到1型遗传性酪氨酸血症小鼠模型体内来修复其基因的1.38 kb插入突变,修复率为0.76±0.25%。Liu等[34]通过分割PE蛋白开发了sPE系统,通过流体动力学注射腺相关病毒(AAV)载体递送sPE系统到I型酪氨酸血症小鼠模型体内,纠正其基因上的G>A致病突变。
尽管引导编辑技术在基因治疗上展现出了明显的优势,如精准性高、PAM限制小和可以实现片段的插入删除等,但其编辑效率仍然需要进一步提高。此外,其体积过大的特点也影响了它的递送效率,限制了其在基因治疗中的应用,亟待解决。
3.3 在植物中的应用
对于作物育种,传统的诱变育种耗时耗力且突变率低,而基因编辑技术的出现大大改善了这一状况。CRISPR/Cas9系统及碱基编辑系统已经在作物育种方面得到了广泛应用,一系列的具有优良性状(如抗病性和除草剂抗性)的种质材料被创制[66~70]。PE系统的出现为快速精准地创制具有优良性状的突变体提供了更有力的工具[71]。
自2019年引导编辑技术出现以来,国内外许多研究团队将引导编辑技术应用于作物性状的改良。其中,Lin等[8]首先将引导编辑技术应用到植物领域,他们建立了植物引导编辑系统PPE,在水稻和小麦中成功进行了点突变、插入和删除编辑,最高效率可达21.8%;在水稻中,Tang等[72]开发了3个版本的植物PE系统,获得最高1.55%的编辑效率;而Xu等[73]使用PE系统编辑水稻原生质体获得突变体频率最高达到31.3%。Xu等[74]利用PE系统编辑水稻除草剂抗性相关的基因,获得被编辑植株的效率最高达到26%。Li等[44]利用PE系统编辑水稻和基因,编辑效率最高达到9.38%,并获得的纯合突变体植株。Butt等[42]使用PE系统编辑水稻、、基因,最高编辑效率达到2%。Xu等[21]使用优化后的PE系统在水稻的13个内源性靶点处实现了平均达到24.3%的编辑效率。
除水稻和小麦外,Lu等[45]将PE系统应用到番茄基因组的编辑,最多在6.7%的植株上检测到了目标编辑,最高编辑效率为1.66%;Jiang等[16]通过增强pegRNA的表达在玉米上实现了更高效率的引导编辑,最高达到了53.2%;Xu等[21]采用在Cas9的N-末端融合逆转录酶及在逆转录酶模板中引入多核苷酸突变的方式来优化PE系统,在玉米原生质体四个靶位点的平均编辑效率达到了6.2%。Perroud等[47]在小立碗藓中测试了PE系统,但最高0.06%的编辑效率明显低于CRISPR/Cas9介导的编辑效率;此外,他们还利用PE系统编辑马铃薯基因,但编辑效率同样较低。Wang等[46]开发了更易于使用的载体来将引导编辑应用于双子叶植物烟草和拟南芥及单子叶植物水稻,验证了其有效性。
PE系统在多种植物中得到广泛应用,彰显了PE系统在植物上巨大的应用价值,但还应注意到的是其在植物中的编辑效率仍明显低于在动物细胞中的效率。因此,仍然需要结合植物的特点对其编辑效率进行有针对性的优化。
3.4 在突变筛选中的应用
基于CRISPR/Cas的基因编辑技术的发展为高通量的遗传变异筛选提供了可能。从最初的全基因组敲除筛选,到HDR介导的饱和突变筛选,再到碱基编辑系统介导的突变筛选,大规模的基因功能研究拥有了更多工具选择[75~78]。引导编辑技术出现后,同样也被应用于遗传筛选研究。Xu等[48]首先在植物上建立基于pegRNA文库的饱和诱变方法PLSM,对于水稻基因中的6个保守残基,分别针对每个残基建立pegRNA文库,最终鉴定到16种类型的赋予水稻除草剂抗性的突变,其中大部分突变尚未在水稻育种中报告或应用,表明PLSM方法在作物改良与农艺重要性状相关基因的直接进化中具有巨大潜力。Derwood等[49]将PE系统与基因位点单倍体化策略相结合,在人类细胞中建立了基于引导编辑的饱和突变系统,他们成功地将此系统应用于和基因变异的功能注释。
4 PE系统的递送方式
将基因编辑工具高效地递送到作用位置是其发挥功能的关键。PE系统出现以来,研究人员已经通过多种方式将其递送到细胞、胚胎及组织中,实现靶位点的高效编辑。其中,最常见的递送方式是质粒DNA的递送。通过转染可以将质粒DNA高效地递送到动物或植物细胞中,还可以通过流体动力学注射质粒到小鼠体内,从而在细胞中大量表达PE系统组分来编辑目标位点[6,11,29,33]。此外,也有研究通过显微注射将质粒DNA导入小鼠胚胎中,实现小鼠胚胎基因组的靶向编辑[9]。mRNA与gRNA的组合也常用来递送基因编辑系统。本课题组通过显微注射PE2 mRNA、pegRNA和nicking sgRNA到小鼠胚胎中,实现了小鼠位点的编辑[10];Nelson等[15]通过核转染将PE2 mRNA、pegRNA和nicking sgRNA递送到HEK293T细胞中来编辑细胞基因组。RNP(ribonucleoprotein)是另一种常见的基因编辑系统递送手段,其优势在于较短的活性持续时间从而诱导较少的脱靶编辑。Petri等[31]以RNA的形式递送PE系统到斑马鱼胚胎和人类细胞中,成功诱导了目标突变;Liu等[34]也以RNP的形式将PE系统递送人类细胞中来编辑靶位点,两项研究都证明了利用RNP来递送PE系统的有效性。AAV是一种的常见病毒载体,一般用于基因编辑系统的体内递送。对于PE系统来说,因为AAV包装容量的限制,所以需要将PE2蛋白拆分成两部分置于两个AAV上。目前,在多项研究中,研究人员通过双AAV策略成功地将PE系统递送到小鼠体内,实现了靶位点的引导编辑[33,35,50,79]。慢病毒(LVs)是另一种用于基因编辑工具递送的病毒载体,与AAV载体相比,具有更大的包装容量,可以包装完整PE2蛋白编码基因,已经在多项研究中得到应用[13,15]。
除了上述递送方法外,还有脂质纳米颗粒、金纳米颗粒、病毒样颗粒等递送手段还未应用在PE系统上[80~82]。这些方法都有各自的优缺点,使用时应该考虑每种基因编辑应用的需要来选择合适的递送方法。
5 引导编辑的生物信息学工具研究
生物信息学工具可以给研究人员提供很大的便利。自PE系统出现以来,已经有多种相关的生物信息学工具被开发出来,主要集中在pegRNA的设计和编辑效率的预测与评估(表3)。
界面友好、方便快捷的pegRNA设计工具有助于研究人员快速得到高效的pegRNA序列用于后续研究。Hsu等[83]开发了PrimeDesign用于pegRNA的设计,并且他们基于PrimeDesign构建了PrimeVar数据库,其中包括用于诱导或纠正来自ClinVar数据库的68500个致病性人类遗传变异的pegRNA和nicking sgRNA的组合。Hwang等[84]开发了PE- Designer,其提供了识别不同PAM的Cas9变体的pegRNA的设计,并且支持包括脊椎动物,植物,昆虫和细菌在内的543种生物。Li等[85]开发了基于深度学习的Easy-Prime pegRNA 设计工具,Easy- Prime可以自动搜索和优化pegRNA 的设计。Lin等[32]开发了应用于植物的PlantPegDesigner,为植物中的pegRNA设计尤其是双pegRNA策略的使用提供了便利。此外,还有pegFinder、PnB Designer、pegIT和PINE-CONE等pegRNA设计工具可供选择[86~89]。
PE系统在基因组不同位点上的编辑效率具有较大的差异,Kim等[90]通过建立pegRNA文库对PE2效率进行高通量分析,系统地评估了PBS和RTT长度及其他因素对PE2效率的影响,基于机器学习建立了预测PE2效率的计算模型,并基于此开发了PE2编辑效率预测工具DeepPE,可辅助pegRNA的设计。
高通量测序在基因编辑领域得到广泛应用,常常被用来对基因编辑效率进行评估。为了便于对PE系统相关的高通量测序数据进行分析,Hwang等[84]开发了PE-Analyzer,其可以快速分析测序数据来评估引导编辑结果。此外,Clement等[91]在原有的CRISPResso2软件基础上增加了其用于分析引导编辑相关的高通量测序数据的模块,极大地促进了PE系统的应用。
6 PE系统的安全风险
基因编辑工具的安全性是其广泛应用尤其是疾病治疗方面应用的前提,且其安全性主要体现在其脱靶风险的大小,但PE系统相比其他基因编辑系统还需要对其逆转录酶的安全性进行评估。对于逆转录酶的安全性,多项研究表明,逆转录酶不会带来安全风险,更多的研究主要是针对其脱靶风险[6,92]。
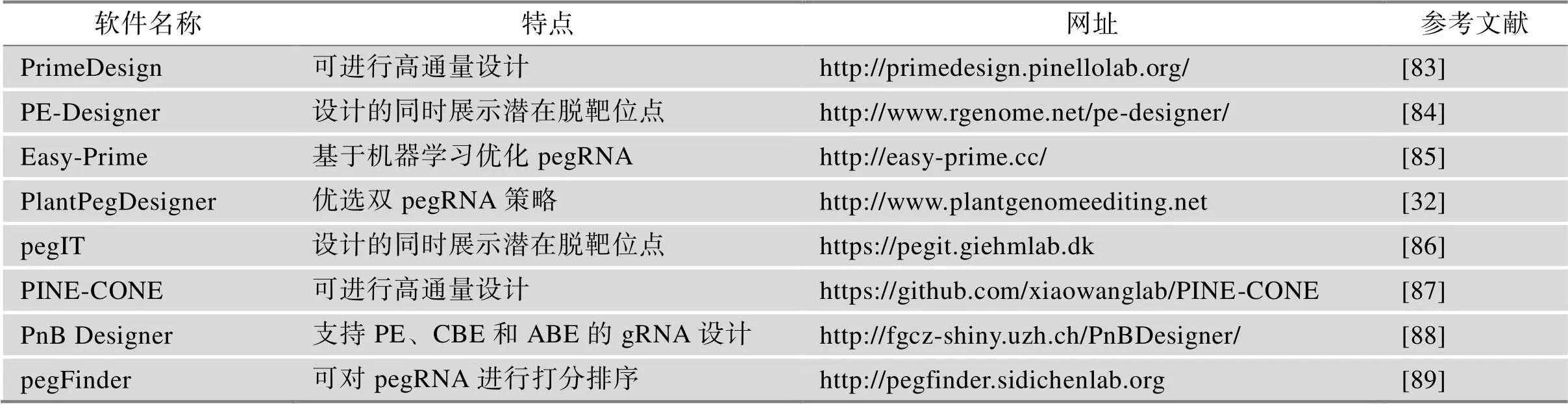
表3 pegRNA设计软件
PE系统在作用过程中的3种核酸杂交决定了PE系统在理论上很难脱靶,这也解释了PE系统的精准性[6]。David R. Liu课题组在开发PE系统过程中对其脱靶效应进行了表征。他们选择已经被报道的具有明确Cas9脱靶的4个基因位点(、、、)及每个位点对应的4个脱靶位点进行测试;结果显示,在16个脱靶位点中,PE系统在其中3个位点发生脱靶编辑,并且只有一个位点脱靶编辑效率大于1%,这说明PE系统具有很高的特异性,其安全性远高于CRISPR/Cas9系统[6]。为了评估PE系统在全基因组水平的特异性,Kim等[93]开发了基于nCas9(H840A) 的nDigenome-seq;nDigenome-seq可以有效地在全基因组范围内分析nCas9(H840A)诱导的DNA单链断裂(SSB)位点,利用靶向深度测序对SSB位点进行分析可以确定引导编辑在这些位点上的脱靶编辑情况;借助nDigenome-seq,Kim等[93]在部分脱靶位点检测到了0.1%~1.9%的PE脱靶编辑,这表明PE在全基因组水平上具有高度特异性;另外,他们还发现利用具有更高特异性的Cas9变体如eSpCas9和Sniper Cas9来构建PE系统,可以进一步提高PE系统在人类细胞中的特异性,这说明在以往研究中降低Cas9脱靶编辑的方法可能同样适用于PE系统特异性的优化,从而为PE系统特异性的优化提供了一个方向。在植物上,Jin等[92]通过靶向深度测序和全基因组测序对水稻原生质体和再生水稻植物中的pegRNA依赖性和pegRNA非依赖性脱靶效应进行了全面分析,他们发现PE系统会产生了极低水平的pegRNA依赖性脱靶(0.00%~0.23%),但不会产生pegRNA非依赖性脱靶。Gao等[94]通过全基因组测序和全转录组测序证明了PE不会产生gRNA非依赖性DNA脱靶和RNA脱靶,进一步证明了PE系统的特异性。
总之,PE系统相比CRIPSR/Cas9系统和碱基编辑系统具有更高的特异性,也更加安全,但仍需进一步降低其脱靶风险,为其后续在基因治疗中的应用奠定基础。
7 结语与展望
引导编辑技术的出现为基因组的精准编辑提供了更加强大的工具,碱基颠换和小片段的插入删除得以高效地实现,在遗传疾病的模拟与治疗和动植物新品种的培育等方面具有极高的应用价值,然而PE系统仍然存在一定的不足之处,最为明显的是引导编辑的效率问题。相比CRISPR/Cas9系统和碱基编辑系统,引导编辑的编辑效率仍然较低;利用PE系统编辑小鼠胚胎获得的基因编辑小鼠多为嵌合小鼠且嵌合比例较低[10,19,53]。虽然已经有大量的研究从不同途径对引导编辑的效率进行了优化,但其仍有一定的提升空间。此外,PE系统的indels也是一个不容忽视的问题。PE2系统由于只在基因组上引入一个切口,其indels频率保持在很低的水平,但PE2的低效性限制了其应用。而对于PE3来说,另一条sgRNA虽然提升了编辑效率,但同时也造成了更多的indels[6];最近有研究表明,在小鼠胚胎中,PE3系统会造成高频率的片段删除等副产物[95],这说明了降低PE系统的indels频率是亟需解决的问题。有研究发现,DNA错配修复机制在indels形成过程中起到了关键作用,通过抑制错配修复机制,可以减少引导编辑介导的indels的产生[20]。相信随着研究的深入,PE系统的作用机制会将被更加清晰地阐释,通过对相关通路的调控,可以将indels降低到更加安全的水平。基因治疗有望根治人类遗传病,基于CRISPP/Cas9的基因编辑疗法已经在临床上取得优异效果,同时基于碱基编辑系统的基因治疗在鼠和猴疾病模型上也成效显著[62,96,97]。PE系统在基因治疗上也表现出了极大的优势,但目前已有的在小鼠成体治疗上效果仍不尽如人意,还停留在初期阶段。未来随着更加高效的PE系统和递送方式的开发,相信PE系统也将会有更加优异的治疗效果。
综上所述,引导编辑技术高效、精准、安全,随着更深入的优化和改造,其必将推动生物医学、动植物遗传育种等领域研究和应用的快速发展。
[1] Cong L, Ran FA, Cox D, Lin SL, Barretto R, Habib N, Hsu PD, Wu XB, Jiang WY, Marraffini LA, Zhang F. Multiplex genome engineering using CRISPR/Cas systems., 2013, 339(6121): 819–823.
[2] Komor AC, Kim YB, Packer MS, Zuris JA, Liu DR. Programmable editing of a target base in genomic DNA without double-stranded DNA cleavage., 2016, 533(7603): 420–424.
[3] Gaudelli NM, Komor AC, Rees HA, Packer MS, Badran AH, Bryson DI, Liu DR. Programmable base editing of A*T to G*C in genomic DNA without DNA cleavage., 2017, 551(7681): 464–471.
[4] Ceccaldi R, Rondinelli B, D'Andrea AD. Repair pathway choices and consequences at the double-strand break., 2016, 26(1): 52–64.
[5] Zong Y, Gao CX. Progress on base editing systems., 2019, 41(9): 777–800.
宗媛, 高彩霞. 碱基编辑系统研究进展. 遗传, 2019, 41(9): 777–800.
[6] Anzalone AV, Randolph PB, Davis JR, Sousa AA, Koblan LW, Levy JM, Chen PJ, Wilson C, Newby GA, Raguram A, Liu DR. Search-and-replace genome editing without double-strand breaks or donor DNA., 2019, 576(7785): 149–157.
[7] Wang XL, Huang XX, Liu Y. Search-and-replace editing of genetic information., 2020, 7(2): 231–232.
[8] Lin QP, Zong Y, Xue CX, Wang SX, Jin S, Zhu ZX, Wang YP, Anzalone AV, Raguram A, Doman JL, Liu DR, Gao CX. Prime genome editing in rice and wheat., 2020, 38(5): 582–585.
[9] Lin JX, Liu XC, Lu ZY, Huang SS, Wu SS, Yu WX, Liu Y, Zheng XG, Huang XX, Sun Q, Qiao YB, Liu Z. Modeling a cataract disorder in mice with prime editing., 2021, 25 494–501.
[10] Liu Y, Li XY, He ST, Huang SH, Li C, Chen YL, Liu Z, Huang XX, Wang XL. Efficient generation of mouse models with the prime editing system., 2020, 6(1): 27.
[11] Jang H, Jo DH, Cho CS, Shin JH, Seo JH, Yu G, Gopalappa R, Kim D, Cho SR, Kim JH, Kim HH. Application of prime editing to the correction of mutations and phenotypes in adult mice with liver and eye diseases., 2022, 6(2): 181–194.
[12] Liu Y, Yang G, Huang SH, Li XY, Wang X, Li GL, Chi T, Chen YL, Huang XX, Wang XL. Enhancing prime editing by Csy4-mediated processing of pegRNA., 2021, 31(10): 1134–1136.
[13] Zhang GQ, Liu Y, Huang SS, Qu SY, Cheng DL, Yao Y, Ji QJ, Wang XL, Huang XX, Liu JH. Enhancement of prime editing via xrRNA motif-joined pegRNA., 2022, 13(1): 1856.
[14] Li XY, Wang X, Sun WJ, Huang SS, Zhong MT, Yao Y, Ji QJ, Huang XX. Enhancing prime editing efficiency by modified pegRNA with RNA G-quadruplexes., 2022, 14(4): mjac022.
[15] Nelson JW, Randolph PB, Shen SP, Everette KA, Chen PJ, Anzalone AV, An M, Newby GA, Chen JC, Hsu A, Liu DR. Engineered pegRNAs improve prime editing efficiency., 2022, 40(3): 402–410.
[16] Jiang YY, Chai YP, Lu MH, Han XL, Lin QP, Zhang Y, Zhang Q, Zhou Y, Wang XC, Gao CX, Chen QJ. Prime editing efficiently generates W542L and S621I double mutations in two ALS genes in maize., 2020, 21(1): 257.
[17] Zhuang Y, Liu JL, Wu H, Zhu QG, Yan YC, Meng HW, Chen PR, Yi CQ. Increasing the efficiency and precision of prime editing with guide RNA pairs., 2022, 18(1): 29–37.
[18] Song M, Lim JM, Min S, Oh JS, Kim DY, Woo JS, Nishimasu H, Cho SR, Yoon S, Kim HH. Generation of a more efficient prime editor 2 by addition of the Rad51 DNA-binding domain., 2021, 12(1): 5617.
[19] Park SJ, Jeong TY, Shin SK, Yoon DE, Lim SY, Kim SP, Choi J, Lee H, Hong JI, Ahn J, Seong JK, Kim K. Targeted mutagenesis in mouse cells and embryos using an enhanced prime editor., 2021, 22(1): 170.
[20] Chen PJ, Hussmann JA, Yan J, Knipping F, Ravisankar P, Chen PF, Chen CD, Nelson JW, Newby GA, Sahin M, Osborn MJ, Weissman JS, Adamson B, Liu DR. Enhanced prime editing systems by manipulating cellular determinants of editing outcomes., 2021, 184(22):5635–5652.e29.
[21] Xu W, Yang YX, Yang BY, Krueger CJ, Xiao QL, Zhao S, Zhang L, Kang GT, Wang FP, Yi HM, Ren W, Li L, He XQ, Zhang CM, Zhang B, Zhao JR, Yang JX. A design optimized prime editor with expanded scope and capability in plants., 2022, 8(1): 455–52.
[22] Zong Y, Liu YJ, Xue CX, Li BS, Li XY, Wang YP, Li J, Liu GW, Huang XX, Cao XF, Gao CX. An engineered prime editor with enhanced editing efficiency in plants., 2022.
[23] Kweon J, Yoon JK, Jang AH, Shin HR, See JE, Jang GY, Kim JI, Kim Y. Engineered prime editors with PAM flexibility., 2021, 29(6): 20015–2007.
[24] Peterka M, Akrap N, Li SY, Wimberger S, Hsieh PP, Degtev D, Bestas B, Barr J, van de Plassche S, Mendoza- Garcia P, Šviković S, Sienski G, Firth M, Maresca M. Harnessing DSB repair to promote efficient homology- dependent and -independent prime editing., 2022, 13(1): 1240.
[25] Ferreira da Silva J, Oliveira GP, Arasa-Verge EA, Kagiou C, Moretton A, Timelthaler G, Jiricny J, Loizou JI. Prime editing efficiency and fidelity are enhanced in the absence of mismatch repair., 2022, 13(1): 760.
[26] Adikusuma F, Lushington C, Arudkumar J, Godahewa GI, Chey YCJ, Gierus L, Piltz S, Geiger A, Jain Y, Reti D, Wilson LOW, Bauer DC, Thomas PQ. Optimized nickase- and nuclease-based prime editing in human and mouse cells., 2021, 49(18): 107855–10795.
[27] Tao R, Wang YH, Hu Y, Jiao YG, Zhou LF, Jiang LR, Li L, He XY, Li M, Yu YM, Chen Q, Yao SH. WT-PE: prime editing with nuclease wild-type Cas9 enables versatile large-scale genome editing., 2022, 7(1): 108.
[28] Wang JL, He Z, Wang GQ, Zhang RW, Duan JY, Gao P, Lei XL, Qiu HY, Zhang CP, Zhang Y, Yin H. Efficient targeted insertion of large DNA fragments without DNA donors., 2022, 19(3): 3315–340.
[29] Jiang TT, Zhang XO, Weng ZP, Xue W. Deletion and replacement of long genomic sequences using prime editing., 2022, 40(2): 2275–234.
[30] Li XS, Zhou LN, Gao BQ, Li GY, Wang X, Wang Y, Wei J, Han WY, Wang ZX, Li JF, Gao RZ, Zhu JJ, Xu WC, Wu J, Yang B, Sun XD, Yang L, Chen J. Highly efficient prime editing by introducing same-sense mutations in pegRNA or stabilizing its structure., 2022, 13(1): 1669.
[31] Petri K, Zhang WT, Ma JY, Schmidts A, Lee H, Horng JE, Kim DY, Kurt IC, Clement K, Hsu JY, Pinello L, Maus MV, Joung JK, Yeh JJ. CRISPR prime editing with ribonucleoprotein complexes in zebrafish and primary human cells., 2022, 40(2): 1895–193.
[32] Lin QP, Jin S, Zong Y, Yu H, Zhu ZX, Liu GW, Kou LQ, Wang YP, Qiu JL, Li JY, Gao CX. High-efficiency prime editing with optimized, paired pegRNAs in plants., 2021, 39(8): 9235–927.
[33] Liu PP, Liang SQ, Zheng CW, Mintzer E, Zhao YG, Ponnienselvan K, Mir A, Sontheimer EJ, Gao GP, Flotte TR, Wolfe SA, Xue W. Improved prime editors enable pathogenic allele correction and cancer modelling in adult mice., 2021, 12(1): 2121.
[34] Liu B, Dong XL, Cheng HY, Zheng CW, Chen ZX, Rodríguez TC, Liang SQ, Xue W, Sontheimer EJ. A split prime editor with untethered reverse transcriptase and circular RNA template., 2022.
[35] Zheng CW, Liang SQ, Liu B, Liu PP, Kwan SY, Wolfe SA, Xue W. A flexible split prime editor using truncated reverse transcriptase improves dual-AAV delivery in mouse liver., 2022, 30(3): 13435–1351.
[36] Oh Y, Lee WJ, Hur JK, Song WJ, Lee Y, Kim H, Gwon LW, Kim YH, Park YH, Kim CH, Lim KS, Song BS, Huh JW, Kim SU, Jun BH, Jung C, Lee SH. Expansion of the prime editing modality with Cas9 from Francisella novicida., 2022, 23(1): 92.
[37] Anzalone AV, Gao XD, Podracky CJ, Nelson AT, Koblan LW, Raguram A, Levy JM, Mercer JAM, Liu DR. Programmable deletion, replacement, integration and inversion of large DNA sequences with twin prime editing., 2022, 40(5): 7315–740.
[38] Choi J, Chen W, Suiter CC, Lee C, Chardon FM, Yang W, Leith A, Daza RM, Martin B, Shendure J. Precise genomic deletions using paired prime editing., 2022, 40(2): 2185–226.
[39] Bosch JA, Birchak G, Perrimon N. Precise genome engineering inusing prime editing., 2021, 118(1): e2021996118.
[40] Qian YQ, Zhao D, Sui TT, Chen M, Liu ZQ, Liu HM, Zhang T, Chen SY, Lai LX, Li ZJ. Efficient and precise generation of Tay-Sachs disease model in rabbit by prime editing system., 2021, 7(1): 50.
[41] Tong YJ, Jørgensen TS, Whitford CM, Weber T, Lee SY. A versatile genetic engineering toolkit forbased on CRISPR-prime editing., 2021, 12(1): 5206.
[42] Butt H, Rao GS, Sedeek K, Aman R, Kamel R, Mahfouz M. Engineering herbicide resistance via prime editing in rice., 2020, 18(12): 23705–2372.
[43] Geurts MH, de Poel E, Pleguezuelos-Manzano C, Oka R, Carrillo L, Andersson-Rolf A, Boretto M, Brunsveld JE, van Boxtel R, Beekman JM, Clevers H. Evaluating CRISPR-based prime editing for cancer modeling and CFTR repair in organoids., 2021, 4(10): e202000940.
[44] Li HY, Li JY, Chen JL, Yan L, Xia LQ. Precise modifications of both exogenous and endogenous genes in rice by prime editing., 2020, 13(5): 6715–674.
[45] Lu YM, Tian YF, Shen RD, Yao Q, Zhong DT, Zhang XN, Zhu JK. Precise genome modification in tomato using an improved prime editing system., 2021, 19(3): 4155–417.
[46] Wang L, Kaya HB, Zhang N, Rai RT, Willmann MR, Carpenter SCD, Read AC, Martin F, Fei ZJ, Leach JE, Martin GB, Bogdanove AJ. Spelling changes and fluorescent tagging with prime editing vectors for plants., 2021, 3: 617553.
[47] Perroud PF, Guyon-Debast A, Veillet F, Kermarrec MP, Chauvin L, Chauvin JE, Gallois JL, Nogué F. Prime editing in the model plant physcomitrium patens and its potential in the tetraploid potato., 2022, 316: 111162.
[48] Xu RF, Liu XS, Li J, Qin RY, Wei PC. Identification of herbicide resistance OsACC1 mutations via in planta prime-editing-library screening in rice., 2021, 7(7): 8885–892.
[49] Erwood S, Bily TMI, Lequyer J, Yan J, Gulati N, Brewer RA, Zhou LC, Pelletier L, Ivakine EA, Cohn RD. Saturation variant interpretation using CRISPR prime editing., 2022, 40(6): 8855–895.
[50] Böck D, Rothgangl T, Villiger L, Schmidheini L, Matsushita M, Mathis N, Ioannidi E, Rimann N, Grisch-Chan HM, Kreutzer S, Kontarakis Z, Kopf M, Thöny B, Schwank G.prime editing of a metabolic liver disease in mice., 2022, 14(636): eabl9238.
[51] Li JN, Liu Z, Huang SS, Wang X, Li GL, Xu YT, Yu WX, Chen SS, Zhang Y, Ma HH, Ke ZF, Chen J, Sun Q, Huang XX. Efficient base editing in G/C-rich regions to model androgen insensitivity syndrome., 2019, 29(2): 1745–176.
[52] Liu Z, Lu ZY, Yang G, Huang SS, Li GL, Feng SJ, Liu YJ, Li JN, Yu WX, Zhang Y, Chen J, Sun Q, Huang XX. Efficient generation of mouse models of human diseases via ABE- and BE-mediated base editing., 2018, 9(1): 2338.
[53] Gao P, Lyu Q, Ghanam AR, Lazzarotto CR, Newby GA, Zhang W, Choi M, Slivano OJ, Holden K, Walker JA, 2nd, Kadina AP, Munroe RJ, Abratte CM, Schimenti JC, Liu DR, Tsai SQ, Long XC, Miano JM. Prime editing in mice reveals the essentiality of a single base in driving tissue-specific gene expression., 2021, 22(1): 83.
[54] Xiang GH, Ren JL, Hai T, Fu R, Yu DW, Wang J, Li W, Wang HY, Zhou Q. Editing porcine IGF2 regulatory element improved meat production in Chinese Bama pigs., 2018, 75(24): 46195–4628.
[55] Crispo M, Mulet AP, Tesson L, Barrera N, Cuadro F, dos Santos-Neto PC, Nguyen TH, Crénéguy A, Brusselle L, Anegón I, Menchaca A. Efficient generation of myostatin knock-out sheep using CRISPR/Cas9 technology and microinjection into zygotes., 2015, 10(8).
[56] Li GW, Zhou SW, Li C, Cai B, Yu HH, Ma BH, Huang Y, Ding YG, Liu Y, Ding Q, He C, Zhou JK, Wang Y, Zhou GX, Li Y, Yan Y, Hua JL, Petersen B, Jiang Y, Sonstegard T, Huang XX, Chen YL, Wang XL. Base pair editing in goat: nonsense codon introgression into FGF5 results in longer hair., 2019, 286(23): 46755–4692.
[57] Lv QY, Yuan L, Deng JC, Chen M, Wang Y, Zeng J, Li ZJ, Lai LX. Efficient generation of myostatin gene mutated rabbit by CRISPR/Cas9., 2016, 6: 25029.
[58] Wang XL, Yu HH, Lei AM, Zhou JK, Zeng WX, Zhu HJ, Dong ZM, Niu YY, Shi BB, Cai B, Liu JW, Huang S, Yan HL, Zhao X, Zhou GX, He XL, Chen XX, Yang YX, Jiang Y, Shi L, Tian X, Wang YJ, Ma BH, Huang XX, Qu L, Chen YL. Generation of gene-modified goats targeting MSTN and FGF5 via zygote injection of CRISPR/Cas9 system., 2015, 5: 13878.
[59] Whitworth KM, Rowland RRR, Ewen CL, Trible BR, Kerrigan MA, Cino-Ozuna AG, Samuel MS, Lightner JE, McLaren DG, Mileham AJ, Wells KD, Prather RS. Gene-edited pigs are protected from porcine reproductive and respiratory syndrome virus., 2016, 34(1): 205–22.
[60] Sun JY, Sun KX, Ding YG, Zhou SW, Gao YW, Chen YL, Wang XL. Editing efficiency of four single base editors in sheep () and goat () fibroblasts., 2021, 29(1): 1695–177.
孙嘉媛, 孙珂欣, 丁一格, 周世卫, 高亚伟, 陈玉林, 王小龙. 四种单碱基编辑器在羊成纤维细胞上的编辑效率. 农业生物技术学报, 2021, 29(1): 1695–177.
[61] Niu XR, Yin SM, Chen X, Shao TT, Li DL. Gene editing technology and its recent progress in disease therapy., 2019, 41(7): 5825–598.
牛煦然, 尹树明, 陈曦, 邵婷婷, 李大力. 基因编辑技术及其在疾病治疗中的研究进展. 遗传, 2019, 41(7): 5825–598.
[62] Gillmore JD, Gane E, Taubel J, Kao J, Fontana M, Maitland ML, Seitzer J, O'Connell D, Walsh KR, Wood K, Phillips J, Xu YX, Amaral A, Boyd AP, Cehelsky JE, McKee MD, Schiermeier A, Harari O, Murphy A, Kyratsous CA, Zambrowicz B, Soltys R, Gutstein DE, Leonard J, Sepp-Lorenzino L, Lebwohl D. CRISPR-Cas9gene editing for transthyretin amyloidosis., 2021, 385(6): 4935–502.
[63] Schene IF, Joore IP, Oka R, Mokry M, van Vugt AHM, van Boxtel R, van der Doef HPJ, van der Laan LJW, Verstegen MMA, van Hasselt PM, Nieuwenhuis EES, Fuchs SA. Prime editing for functional repair in patient- derived disease models., 2020, 11(1): 5352.
[64] Chemello F, Chai AC, Li H, Rodriguez-Caycedo C, Sanchez-Ortiz E, Atmanli A, Mireault AA, Liu N, Bassel- Duby R, Olson EN. Precise correction of Duchenne muscular dystrophy exon deletion mutations by base and prime editing., 2021, 7(18).
[65] Kim Y, Hong SA, Yu J, Eom J, Jang K, Yoon S, Hong DH, Seo D, Lee SN, Woo JS, Jeong J, Bae S, Choi D. Adenine base editing and prime editing of chemically derived hepatic progenitors rescue genetic liver disease., 2021, 28(9): 16145–1624.e5.
[66] Lu YM, Zhu JK. Precise editing of a target base in the rice genome using a modified CRISPR/Cas9 system., 2017, 10(3): 5235–525.
[67] Zong Y, Wang YP, Li C, Zhang R, Chen KL, Ran YD, Qiu JL, Wang DW, Gao CX. Precise base editing in rice, wheat and maize with a Cas9-cytidine deaminase fusion., 2017, 35(5): 4385–440.
[68] Hua K, Tao XP, Han PJ, Wang R, Zhu JK. Genome engineering in rice using Cas9 variants that recognize NG PAM sequences., 2019, 12(7): 10035–1014.
[69] Li JY, Sun YW, Du JL, Zhao YD, Xia LQ. Generation of targeted point mutations in rice by a modified CRISPR/ Cas9 system., 2017, 10(3): 5265–529.
[70] Zhang R, Liu JX, Chai ZZ, Chen S, Bai Y, Zong Y, Chen KL, Li JY, Jiang LJ. Gao CX. Generation of herbicide tolerance traits and a new selectable marker in wheat using base editing., 2019, 5(5): 4805–485.
[71] Qin RY, Wei PC. Prime editing creates a novel dimension of plant precise genome editing., 2020, 42(6): 5195–523.
秦瑞英, 魏鹏程. Prime editing引导植物基因组精确编辑新局面. 遗传, 2020, 42(6): 519–523.
[72] Tang X, Sretenovic S, Ren QR, Jia XY, Li MK, Fan TT, Yin DS, Xiang SY, Guo YC, Liu L, Zheng XL, Qi YP, Zhang Y. Plant prime editors enable precise gene editing in rice cells., 2020, 13(5): 6675–670.
[73] Xu RF, Li J, Liu XS, Shan TF, Qin RY, Wei PC. Development of plant prime-editing systems for precise genome editing., 2020, 1(3): 100043.
[74] Xu W, Zhang CW, Yang YX, Zhao S, Kang GT, He XQ, Song JL, Yang JX. Versatile nucleotides substitution in plant using an improved prime editing system., 2020, 13(5): 6755–678.
[75] Wang T, Wei JJ, Sabatini DM, Lander ES. Genetic screens in human cells using the CRISPR-Cas9 system., 2014, 343(6166): 805–84.
[76] Hanna RE, Hegde M, Fagre CR, DeWeirdt PC, Sangree AK, Szegletes Z, Griffith A, Feeley MN, Sanson KR, Baidi Y, Koblan LW, Liu DR, Neal JT, Doench JG. Massively parallel assessment of human variants with base editor screens., 2021, 184(4): 10645–1080.e20.
[77] Cuella-Martin R, Hayward SB, Fan X, Chen X, Huang JW, Taglialatela A, Leuzzi G, Zhao JF, Rabadan R, Lu C, Shen Y, Ciccia A. Functional interrogation of DNA damage response variants with base editing screens., 2021, 184(4): 10815–1097.e19.
[78] Findlay GM, Boyle EA, Hause RJ, Klein JC, Shendure J. Saturation editing of genomic regions by multiplex homology-directed repair., 2014, 513(7516): 1205–123.
[79] Zhi SY, Chen YX, Wu GL, Wen JK, Wu JN, Liu QY, Li Y, Kang R, Hu SH, Wang JH, Liang PP, Huang JJ. Dual- AAV delivering split prime editor system forgenome editing., 2022, 30(1): 2835–294.
[80] Banskota S, Raguram A, Suh S, Du SW, Davis JR, Choi EH, Wang X, Nielsen SC, Newby GA, Randolph PB, Osborn MJ, Musunuru K, Palczewski K, Liu DR. Engineered virus-like particles for efficientdelivery of therapeutic proteins., 2022, 185(2): 2505–265.e16.
[81] Pensado A, Seijo B, Sanchez A. Current strategies for DNA therapy based on lipid nanocarriers., 2014, 11(11): 17215–1731.
[82] Lee K, Conboy M, Park HM, Jiang FG, Kim HJ, Dewitt MA, Mackley VA, Chang K, Rao A, Skinner C, Shobha T, Mehdipour M, Liu H, Huang WC, Lan F, Bray NL, Li S, Corn JE, Kataoka K, Doudna JA, Conboy I, Murthy N. Nanoparticle delivery of Cas9 ribonucleoprotein and donor DNAinduces homology-directed DNA repair., 2017, 1: 8895–901.
[83] Hsu JY, Grünewald J, Szalay R, Shih J, Anzalone AV, Lam KC, Shen MW, Petri K, Liu DR, Joung JK, Pinello L. PrimeDesign software for rapid and simplified design of prime editing guide RNAs., 2021, 12(1): 1034.
[84] Hwang GH, Jeong YK, Habib O, Hong SA, Lim K, Kim JS, Bae S. PE-Designer and PE-Analyzer: web-based design and analysis tools for CRISPR prime editing., 2021, 49(W1): W4995–W504.
[85] Li YC, Chen JJ, Tsai SQ, Cheng Y. Easy-Prime: a machine learning-based prime editor design tool., 2021, 22(1): 235.
[86] Anderson MV, Haldrup J, Thomsen EA, Wolff JH, Mikkelsen JG. PegIT—a web-based design tool for primeediting., 2021, 49(W1): W5055–W509.
[87] Standage-Beier K, Tekel SJ, Brafman DA, Wang X. Prime editing guide RNA eesign automation using PINE- CONE., 2021, 10(2): 4225–427.
[88] Siegner SM, Karasu ME, Schroder MS, Kontarakis Z, Corn JE. PnB Designer: a web application to design prime and base editor guide RNAs for animals and plants., 2021, 22(1): 101.
[89] Chow RD, Chen JS, Shen J, Chen S. A web tool for the design of prime-editing guide RNAs., 2021, 5(2): 1905–194.
[90] Kim HK, Yu G, Park J, Min S, Lee S, Yoon S, Kim HH. Predicting the efficiency of prime editing guide RNAs in human cells., 2021, 39(2): 1985–206.
[91] Clement K, Rees H, Canver MC, Gehrke JM, Farouni R, Hsu JY, Cole MA, Liu DR, Joung JK, Bauer DE, Pinello L. CRISPResso2 provides accurate and rapid genome editing sequence analysis., 2019, 37(3): 2245–226.
[92] Jin S, Lin QP, Luo YF, Zhu ZX, Liu GW, Li YJ, Chen KL, Qiu JL, Gao CX. Genome-wide specificity of prime editors in plants., 2021, 39(10): 12925–1299.
[93] Kim DY, Moon SB, Ko JH, Kim YS, Kim D. Unbiased investigation of specificities of prime editing systems in human cells., 2020, 48(18): 105765– 10589.
[94] Gao RZ, Fu ZC, Li XY, Wang Y, Wei J, Li GY, Wang LJ, Wu J, Huang XX, Yang L, Chen J. Genomic and transcriptomic analyses of prime editing guide RNA-independent off-target effects by prime editors., 2022, 5(2): 2765–293.
[95] Aida T, Wilde JJ, Yang L, Hou Y, Li M, Xu D, Lin J, Qi P, Lu Z, Feng G. Prime editing primarily induces undesired outcomes in mice., 2020.
[96] Musunuru K, Chadwick AC, Mizoguchi T, Garcia SP, DeNizio JE, Reiss CW, Wang K, Iyer S, Dutta C, Clendaniel V, Amaonye M, Beach A, Berth K, Biswas S, Braun MC, Chen HM, Colace TV, Ganey JD, Gangopadhyay SA, Garrity R, Kasiewicz LN, Lavoie J, Madsen JA, Matsumoto Y, Mazzola AM, Nasrullah YS, Nneji J, Ren H, Sanjeev A, Shay M, Stahley MR, Fan SHY, Tam YK, Gaudelli NM, Ciaramella G, Stolz LE, Malyala P, Cheng CJ, Rajeev KG, Rohde E, Bellinger AM, Kathiresan S.CRISPR base editing of PCSK9 durably lowers cholesterol in primates., 2021, 593(7859): 4295–434.
[97] Koblan LW, Erdos MR, Wilson C, Cabral WA, Levy JM, Xiong ZM, Tavarez UL, Davison LM, Gete YG, Mao XJ, Newby GA, Doherty SP, Narisu N, Sheng QH, Krilow C, Lin CY, Gordon LB, Cao K, Collins FS, Brown JD, Liu DR.base editing rescues Hutchinson-Gilford progeria syndrome in mice., 2021, 589(7843): 6085–614.
Prime editing: a search and replace tool with versatile base changes
Yao Liu, Xianhui Zhou, Shuhong Huang, Xiaolong Wang
Prime editing is a newly developed CRISPR/Cas system-based genome editing technique. The effector of prime editor (PE) is termed PE2, which is generated by fusing a reverse transcriptase (RT) with a Cas9 H840A nickase. The guide RNA of PE is termed prime editing guide RNA (pegRNA), which consists of a single guide RNA (sgRNA) with a 3′ extension containing the RT template (RTT) and primer binding site (PBS). PE can install all 12 types of point mutations, small insertions and deletions and combinations thereof. Since its emergence in 2019, with the high versatility and specificity, PE has been applied to many living organisms, including animals, plants and bacteria. This led to many explorations of PE on gene therapy and genetic improvement in agriculture. In this review, we systematically describe the development, characteristics, optimizations, applications and security of PE. In addition, we discuss the future applications of PE. We expect that this review will help researchers to grasp and better use PE.
prime editor; CRISPR/Cas9; optimization; application
2022-07-23;
2022-09-01;
2022-09-16
国家自然科学基金项目(编号:31972526, 32161143010)资助[Supported by the National Natural Science Foundation of China (Nos. 31972526, 32161143010)]
刘尧,在读博士研究生,专业方向:动物遗传育种。E-mail: yaoliu@nwafu.edu.cn
王小龙,博士,教授,研究方向:动物生物育种。E-mail: xiaolongwang@nwafu.edu.cn
10.16288/j.yczz.22-156
(责任编委: 李大力)
猜你喜欢
分子催化(2022年1期)2022-11-02
军事文摘(2022年16期)2022-08-24
今日农业(2021年11期)2021-08-13
昆明医科大学学报(2021年3期)2021-07-22
烟草科技(2021年6期)2021-06-24
科学之谜(2021年2期)2021-04-25
中国生殖健康(2020年4期)2020-12-09
教学考试(高考生物)(2020年6期)2020-11-23
中西医结合肝病杂志(2020年2期)2020-10-27
学苑创造·B版(2019年5期)2019-06-14
