
中国草原红牛PDK4 基因克隆及其组织表达差异性研究
2022-10-12 13:17马彦茹于永生秦立红赵玉民
中国畜牧杂志 2022年10期
马彦茹 ,于永生,肖 成,曹 阳,秦立红,赵玉民,3,4,5,吴 健,3,4*
(1.吉林农业大学动物科学技术学院,吉林长春 130000;2.吉林省农业科学院,吉林公主岭 136100;3.农业农村部肉牛遗传育种重点实验室,吉林公主岭 136100;4.农业农村部东北种牛性能测定中心,吉林公主岭 136100;5.吉林省肉牛繁育及养殖技术科技创新中心,吉林公主岭 136100)
中国草原红牛是采用英国短角牛(Shorthorn)与蒙古牛杂交培育出的具有耐粗饲、抗病力强、适应性强等优良特性的肉乳兼用型品种。丙酮酸脱氢酶激酶4(Pyruvate Dehydrogenase Kinase 4,)是丙酮酸脱氢酶激酶的重要亚型之一,位于4 号染色体上,全长13 474 nt,具有11 个外显子,是一种线粒体酶。通过抑制丙酮酸脱氢酶活性来抑制丙酮酸进入三羧酸循环(Tricarboxylic Acid Cycle,TCA cycle),研究表明其在葡萄糖的消耗和脂肪酸的代谢转换中起重要作用。目前,基因在牛的脂肪代谢方面研究较少。赵伟明等研究发现基因在2 种牛背膘脂肪中的表达量不同,表明基因可能与脂肪沉积相关。潘鹏丞等发现基因在陆川猪各组织中均有表达,其中皮下脂肪中的表达量最高。张亚楠研究发现山羊基因在肺脏与臂三头肌2 种组织中表达最高,并且极显著高于其他组织。近年,虽然有一些对牛基因的研究,但未见在中国草原红牛方面的报道。因此,本研究通过克隆草原红牛基因完整的CDS 序列并进行生物信息学分析,明确该基因在草原红牛不同组织间及成脂分化过程中的表达差异,为进一步开展基因功能及其在草原红牛上的作用机制研究提供参考资料。
1 材料与方法
1.1 材料
1.1.1 样品采集 从吉林省农业科学院草原红牛育种实验场选取6 头健康的24 月龄中国草原红牛为实验动物,屠宰后采集心脏、肝脏、脾脏、肺脏、肾脏、胃、十二指肠、背最长肌和脂肪等组织样品,样品采集后迅速放入液氮灌中保存。
1.1.2 主要试剂、仪器 胶回收纯化试剂盒(Omega)、pMD18-T 载体和5×PrimeScript RT Master Mix(TaKaRa)、LB 液体及固体培养基(北京金桥科技有限公司)、Trizol试剂(Invitrogen)、质粒小量提取试剂盒(Axygeno)、2×ES Taq Master Mix(CWBIO)、LightCycler 480 SYBR Green ⅠMaste(Roche 中 国);uawell-Q5000超微量分光光度计(北京鼎盛生物公司)。
1.2 方法
1.2.1 引物设计与合成 根据GenBank 数据库中公布的牛mRNA(登录号:NM_001101883.1)序列利用Primer Premier 5.0 软件设计引物,再进行RT-PCR和实时荧光定量PCR 引物扩增,其中-基因作为实时荧光定量PCR 的内参基因使用。引物详情见表1。所有引物均由苏州金唯智生物科技有限公司合成。
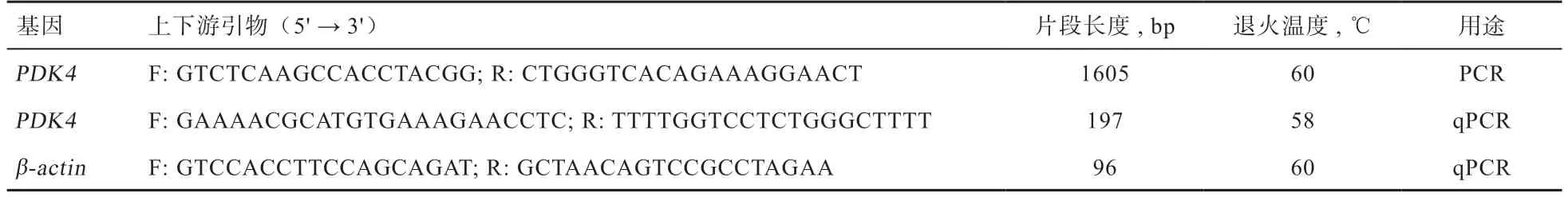
表1 引物序列
1.2.2 提取RNA 及制备cDNA 将采集到的草原红牛心、脾、胃、背最长肌等9 种组织利用Trizol 法提取总RNA,将RNA 逆转录为cDNA,逆转录体系为500 ng的RNA,1 μL 的5×PrimeScript RT Master Mix,然后将RNase-Free HO 补至10 μL。反应程序:37℃15 min;85℃5 s。cDNA 于-20℃保存备用。
1.2.3 PCR 扩增 以1.2.2 提取的cDNA 为模板,通过特异性引物进行PCR 扩增。反应体系为20 μL,其中2×Taq MasterMix 10 μL,上下游引物(1 μmol/L)各0.5 μL,cDNA 1 μL,ddHO 8 μL。反应程序为:94℃5 min;94℃30 s、60℃30 s、72℃49 s,34 个循环;最后72℃8 min。PCR 产物进行1.5%琼脂糖凝胶电泳检测,然后利用胶回收试剂盒回收目的条带。
1.2.4 载体的构建与克隆测序 利用pMD18-T 载体与胶回收产物连接,连接体系10 μL:胶回收产物4 μL、pMD18-T 载体1 μL、solution I 5 μL,混匀4℃过夜。次日在大肠杆菌DH5感受态细胞中转入连接产物,转化体系共55 μL,其中连接产物5 μL、DH5感受态细胞50 μL。轻弹混匀后冰浴30 min,42℃水浴90 s,再冰浴2~3 min;在800 μL 的无抗生素LB 培养液中加入连接产物,置于摇床室温活化45 min,将活化好的菌液取200 μL 涂布于含氨苄青霉素的LB 固体培养基中,倒置放入37℃恒温培养箱中12~16 h。在含氨苄青霉素的LB 培养液中加入从菌板上挑取的单菌落,37℃摇床过夜。利用质粒小量提取试剂盒提取质粒后进行菌落PCR 验证,将阳性菌液送至苏州金唯智生物科技有限公司测序。
1.2.5基因序列分析 利用DNAMAN 软件将测序拼接出的草原红牛基因的CDS 区与NCBI 中牛基因序列进行比对,并利用EditSeq 将测序序列翻译为氨基酸序列。利用NCBI-BLAST 分析序列同源性,将获得的不同物种序列利用Mega 6.0 构建基因系统进化树。利用ProtParam 软件分析基因蛋白的理化性质,Protean 和SWISS-MODEL 软件预测PDK4 蛋白的二级与三级结构;利用TargetP 与PSORT II、ProtScale、NetPhos3.1、NetNGlyc、TMPred、Signal IP 软件分别预测PDK4 蛋白的亚细胞定位、亲疏水性、潜在磷酸化位点、蛋白糖基化位点、跨膜区结构及信号肽。通过String 软件分析PDK4 蛋白分子与其他分子的互作网络。
1.2.6 实时荧光定量PCR 以1.2.2 中的方法制备的各组织cDNA 为模板,-actin(上下游均1 μmol/L)为内参基因,进行实时荧光定量PCR 反应。反应体系为20 μL,其 中LightCycler 480 SYBR Green I Maste 10 μL,上、下游引物(1 μmol/L)各0.5 μL,模板1 μL,ddHO 8 μL。反应程序:95℃5 min,95℃10 s,58℃15 s,72℃20 s,共43 个循环。
1.2.7基因在成脂分化过程中的mRNA 表达 复苏第三代的草原红牛前体脂肪细胞,待汇合度达70%~80% 进行诱导分化;再利用Trizol 法提取分化过程中的0、2、4、6 d 的总RNA,反转录为cDNA,进行实时荧光定量PCR,检测基因在成脂分化过程中的相对表达量。
1.2.8 统计分析 实时荧光定量PCR 结果采用2方法进行统计分析,显著性分析利用GraphPad Prism 软件进行检验,<0.05 判定为差异显著。
2 结果与分析
2.1 草原红牛基因PCR 扩增及克隆 如图1 所示,草原红牛基因的PCR 扩增产物条带明亮清晰,条带大小约为1 605 bp,与预期目的条带大小一致。然后利用TA 克隆的方法,成功获得基因的阳性重组菌落,测序检验后,证明成功克隆出草原红牛基因的CDS 区。

图1 中国草原红牛PDK4 基因编码区PCR 扩增电泳图
2.2基因同源性分析和系统进化树构建 构建系统进化树,草原红牛与其他11 种动物同源性关系如图2、3 所示。由图2 可知,野牦牛的同源性与草原红牛的同源性最高,达99.8%,而与埃及果蝠的同源性最低(89.7%),与水牛(XM_006065918.2)、山羊(XM_005678949.3)、羚羊(XM_040260478.1)、绵羊(XM_004007738.4)、瘤牛(XM_019959714.1)、虎鲸(XM_004265573.2)、宽吻海豚(XM_00432293 2.3)、野猪(NM_001159306.1)和狐蝠(XM_03987804 7.1)的同源性分别为98.6%、97.6%、97.5%、97.5%、96.7%、94.6%、94.3%、93.5%、90.4%。由图3 可知,草原红牛和野牦牛进化关系密切,属于一个分支,与埃及果蝠的亲缘关系最远。

图2 不同物种间PDK4 基因核苷酸序列同源性对比
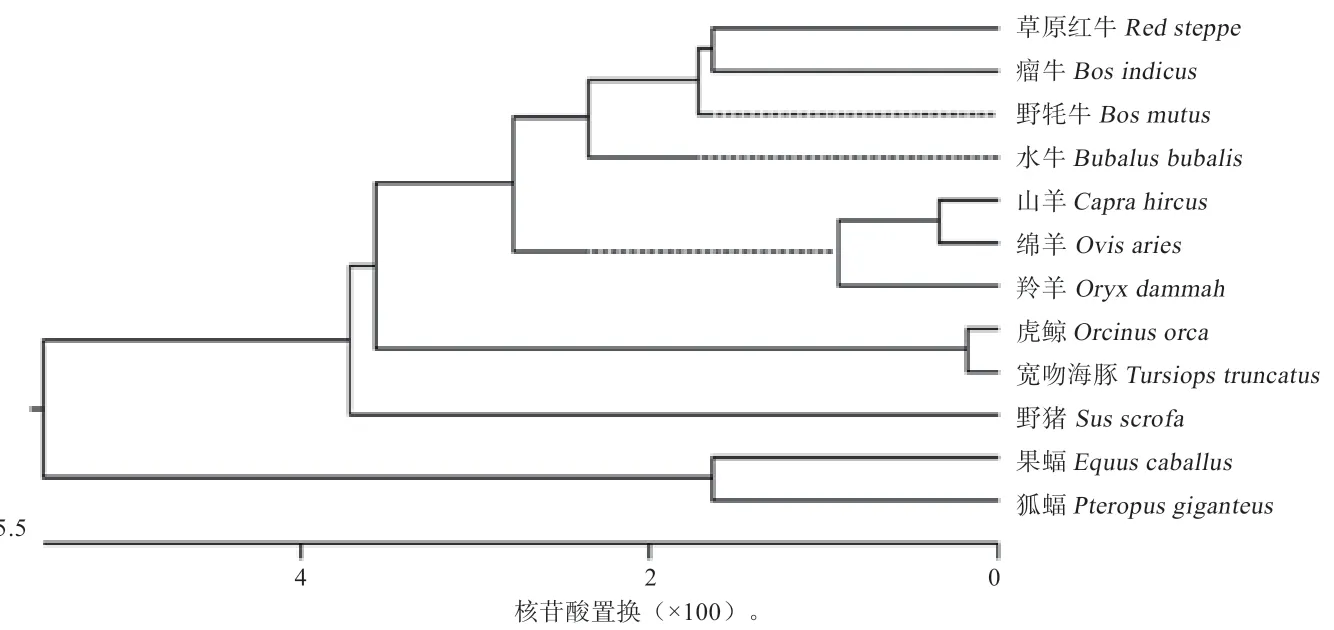
图3 不同物种间PDK4 基因系统进化树
2.3 PDK4 蛋白理化性质分析 利用ExPASy 在线软件分析PDK4 蛋白理化特性,如表2 所示,草原红牛基因编码407 个氨基酸,其中带负电荷和正电荷的氨基酸残基总数分别为42 和41。PDK4 蛋白分子式为CHNOS,分子质量为46.158 ku,理论等电点为6.83,属于酸性氨基酸。脂肪系数为91.25,总平均亲水性为-0.169,属于可溶性蛋白。消光系数为43 695,不稳定指数为45.91,属于不稳定蛋白质。
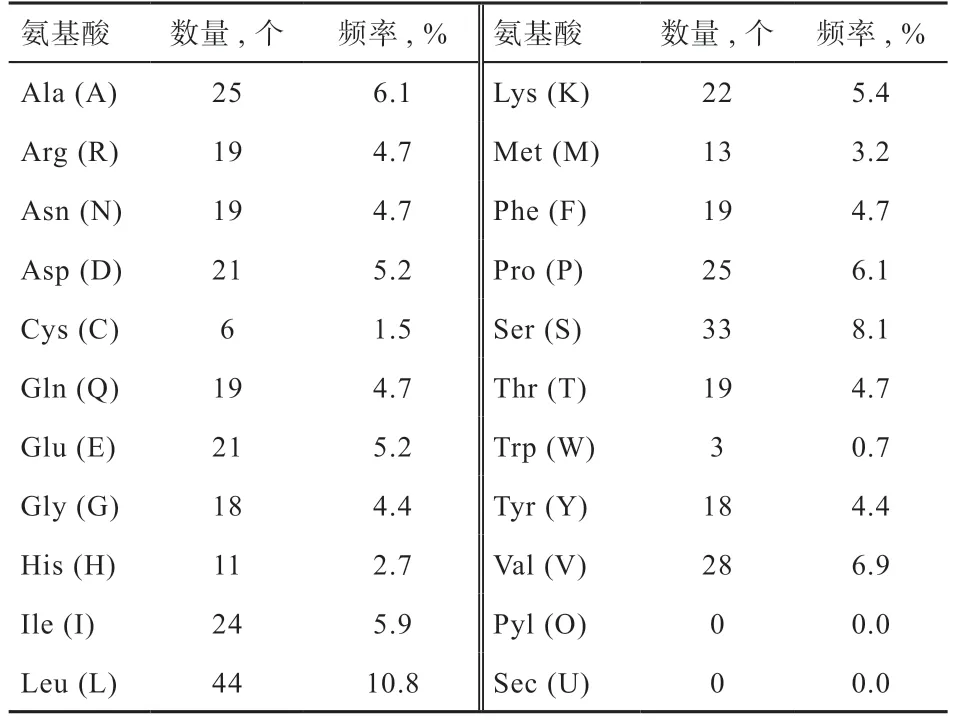
表2 草原红牛PDK4 蛋白的氨基酸组成
2.4 PDK4 蛋白亚细胞定位及蛋白的二级与三级结构预测分析 通过TargetP 在线软件预测结果如表3 所示,PDK4 蛋白属于线粒体靶向肽;同时,PSORT II 在线软件预测结果显示PDK4 蛋白主要位于细胞质(30.4%)、细胞核(21.7%)、线粒体(39.1%)、质膜(4.3%)、液泡(4.3%)中,证明PDK4 蛋白主要在线粒体中存在;PDK4 蛋白二级结构预测结果如图4 所示,可见-折叠占比最大(32.9%),其次是-转角(26%)、无规则卷曲(23.2%),-螺旋占比最少(17.8%)。三级结构预测结果如图5 所示,可见蛋白质结构主要通过-折叠和-转角构成,再经由无规则卷曲连接。
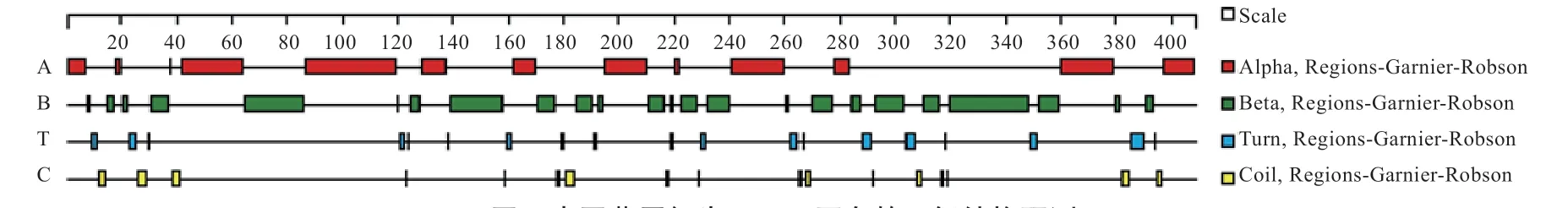
图4 中国草原红牛PDK4 蛋白的二级结构预测

图5 中国草原红牛PDK4 蛋白的三级结构预测

表3 TargetP 分析结果
2.5 PDK4 蛋白亲水性、疏水性和潜在的磷酸化位点分析 ProtScale 在线分析结果如图6 所示,PDK4 蛋白氨基酸残基大多集中于亲水性区,且亲水性最大值为-3.044,疏水性最大值为2.211,属于亲水性蛋白。在线NetPhos3.1 分析结果如图7 所示,可见PDK4 氨基酸序列存在33 个潜在的磷酸化位点,其中丝氨酸21处、苏氨酸9 处、酪氨酸3 处。
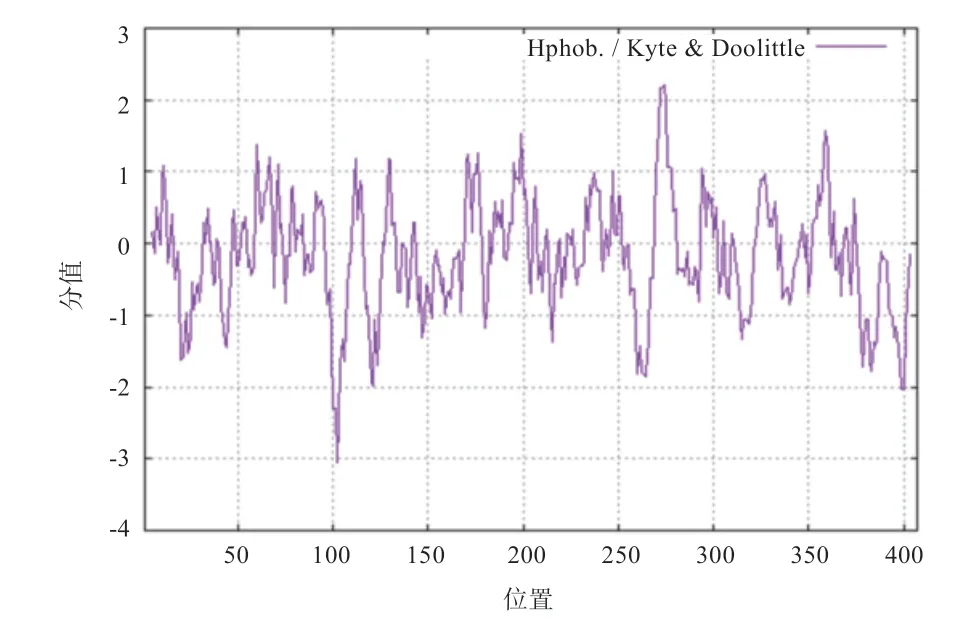
图6 中国草原红牛PDK4 蛋白亲疏水性分析

图7 中国草原红牛PDK4 蛋白的磷酸化位点预测
2.6跨膜区、蛋白糖基化位点及信号肽预测 利用TMPred 在线分析发现PDK4 氨基酸序列在(168,187)位氨基酸处有跨膜区。NetNGlyc1.0 分析表明,PDK4蛋白在76、182、221 位氨基酸处存在3 个N-糖基化潜在位点(图8)。在线NetNGlyc4.0 分析表明,位于第10、23、303、305、307、308、317、376、383、384、385、394 位氨基酸处存在12 个O-糖基化潜在位点。Signal IP 在线预测发现,PDK4 蛋白不存在信号肽。

图8 中国草原红牛PDK4 蛋白的N-糖基化位点预测
2.7 PDK4 蛋白分子互作网络分析 利用String 在线软件分析PDK4 蛋白分子与其他分子的互作网络,结果显示,PDK4 蛋白可以与肉碱棕榈酰转移酶I(CPT1)的亚型(CPT1A)、核糖体蛋白S6 激酶beta-1(RPS6KB1)、过氧化物酶体增殖物激活受体(PPARA)、丙酮酸脱氢酶(PDH)复合物的亚基(PDHA1)、CPT1 的亚型CPT1B、丝氨酸/苏氨酸蛋白激酶密切相关激酶1(AKT1)、线粒体转运蛋白3(UCP3)、过氧化物酶体增殖物激活受体共激活物1-(PPARGC1A)、血管生成素相关蛋白4(ANGPTL4)、磷酸烯醇丙酮酸羧激酶1(PCK1)等分子形成互作网络,在体内发挥作用(图9)。如在MDA-MB-231 细胞中,脂肪酸氧化在PPARA 过表达时增加了近3 倍,在TTA 处理时增加了4 倍,在这些条件下,表达高度增加;在各种营养条件下,通过PDHA1 的磷酸化来调节多种组织和细胞类型的能量稳态;UCP3 和共同参与胰岛素抵抗的葡萄糖和脂质代谢过程。
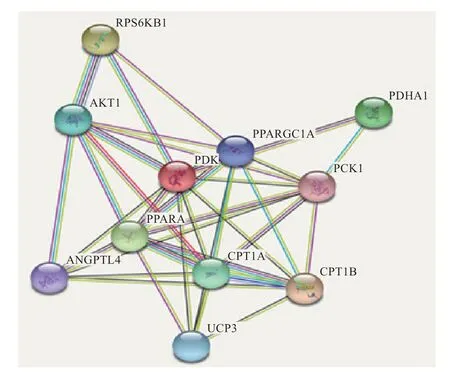
图9 中国草原红牛PDK4 蛋白分子互作网络
2.8基因在不同组织中的表达差异分析 如图10所示,在中国草原红牛的心、肝、脾、肺、肾、胃、十二指肠、背最长肌和脂肪组织中,基因在各组织中均显著表达,表达量最高为背最长肌(<0.05),在肺、肾、肠、脂肪中表达量较低。
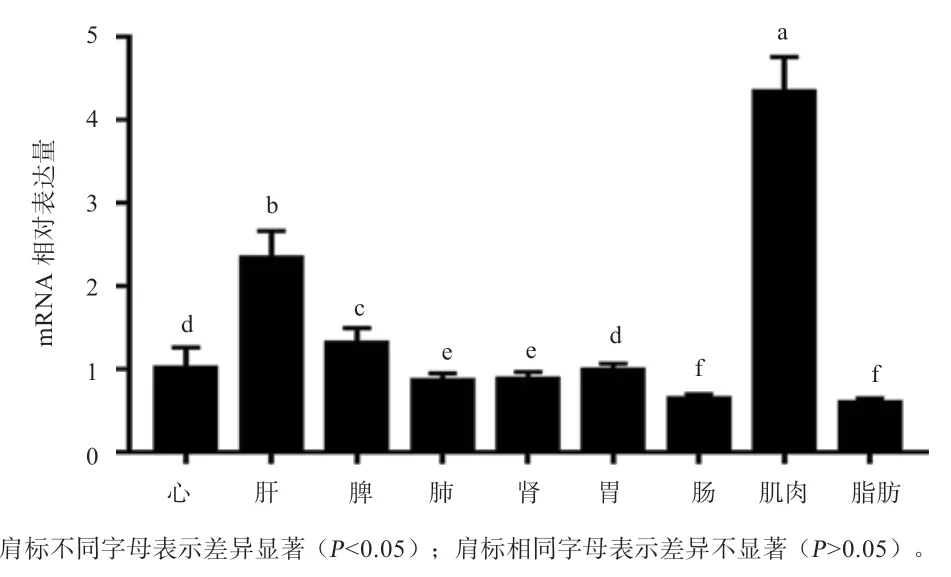
图10 PDK4 在中国草原红牛各组织间的表达差异
2.9基因在成脂分化过程中的表达分析 利用实时荧光定量PCR 对基因在草原红牛前体脂肪细胞分化的第0、2、4、6 天的mRNA 表达量进行检测,如图11 所示,前体脂肪细胞向脂肪细胞分化过程中,在第0 天表达量最低,与第0 天相比第2 天表达量显著增高,且第4 天表达量显著高于第2 天,第4 天与第6 天之间无显著差异。
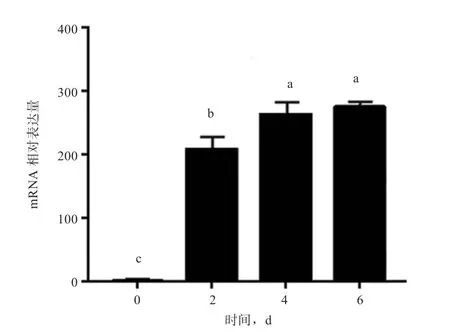
图11 PDK4 在前体脂肪细胞分化过程中的表达
3 讨 论
目前,对基因的研究主要集中在癌症、糖尿病及代谢综合征等疾病上,Guda 等研究表明基因在乳腺癌中相对高表达,其表达与患者预后不良相关。Pin 等研究发现癌症恶病质患者PDK4 蛋白表达升高与代谢异常和肌肉萎缩之间存在关联。Park 等研究发现肝的上调促进了胰高血糖素介导的糖异生基因的表达,而肝的敲除或抑制对糖异生基因的表达产生相反的影响并降低肝葡萄糖的产生。Pettersen 等通过TTA 治疗的大鼠与经PPARA 过表达MDA-MB-231 细胞中,发现表达高度增加,表明mRNA 表达上调与代谢适应细胞和大鼠组织中线粒体脂肪酸氧化活性的增加密切相关。
基因目前在畜禽动物中研究相对较多。通常在具有高能量需求的组织中高度表达,包括心脏、骨骼肌、肝脏、肾脏、胰岛和哺乳期乳腺。Cadoudal 等研究表明的抑制或敲低减弱了噻唑烷二酮(TZDs)对甘油生成的影响,从而证明了这种酶在脂肪酸循环中的重要作用。杨洋等发现,基因在从江香猪和大白猪不同组织中表达量不同,从江香猪在肾脏中表达量最高,大白猪中脂肪表达量最高,表明基因在不同物种间存在差异性。奚子英等研究发现,基因在生长速度差异大的杏花鸡与隐性白洛克鸡的组织表达显著不同,表明基因对家禽生长有一定的调控作用。张榕婧等研究发现基因在肌肉组织(包括腿肌、胸肌、心脏)表达最高,与本实验研究结果一致,基因在草原红牛背最长肌中表达量最高。
本研究成功获得草原红牛基因编码区,全长1 223 bp,预测其编码407 个氨基酸,蛋白分子式为CHNOS,分子质量为46.158 ku,总平均亲水性为-0.169,蛋白为亲水性蛋白,草原红牛基因测序结果与Zimin 等对家牛的基因组使用分层和全基因组鸟枪法混合测序法的结果一致。草原红牛克隆序列与野牦牛和水牛的同源性较高(99.8%和98.6%),说明该基因保守性较强;PDK4 作为一种重要的线粒体蛋白,其亚细胞定位预测显示其主要存在于线粒体中;预测基因存在3 个N-糖基化潜在位点和12 个O-糖基化潜在位点,33 个潜在的磷酸化位点,蛋白糖基化与磷酸化对于调节细胞功能具有重要作用。实时荧光定量PCR 结果表明,基因在草原红牛不同组织间均显著表达,在背最长肌中表达最高,其次是肝脏,在脾、心脏、胃中中度表达;此外基因在草原红牛前体脂肪细胞分化过程中表达差异显著。本研究进一步介绍了基因的功能,为今后研究基因对草原红牛脂代谢的影响提供资料。
4 结 论
本研究成功克隆出中国草原红牛基因编码区,全长1 223 bp,共编码407 个氨基酸,PDK4 蛋白为亲水性蛋白,其与多种蛋白形成互作网络,共同进行调控作用;基因在中国草原红牛各组织间表达差异显著,其中背最长肌表达最高,十二指肠和脂肪表达最低。草原红牛前体脂肪细胞分化过程中,基因mRNA 表达量在0~4 d 呈上升趋势,4~6 d 之间无显著差异。
猜你喜欢
中国农业科学(2022年16期)2022-09-19
广西植物(2022年8期)2022-09-07
中国农业科学(2022年15期)2022-08-09
电脑报(2020年40期)2020-11-06
故事作文·低年级(2020年5期)2020-05-14
中学生物学(2019年7期)2019-10-17
电脑知识与技术(2018年19期)2018-11-01
中学生物学(2017年7期)2017-08-23
科技资讯(2016年32期)2017-03-31
小学阅读指南·高年级版(2015年5期)2015-09-10
