
以甘油为碳源生产低分子质量β-1,3-葡聚糖的发酵工艺
2022-09-17 12:57李敏朱莉詹晓北
食品与发酵工业 2022年17期
李敏,朱莉,詹晓北*
1(教育部糖化学与生物技术重点实验室,江南大学 生物工程学院,江苏 无锡,214122) 2(无锡格莱克斯生物科技有限公司,江苏 无锡,214125)
低分子质量β-1,3-葡聚糖是一种葡萄糖以β-1,3-糖苷键连接而成的同型寡糖[1],相比于高聚合度的β-1,3-葡聚糖,低分子质量β-1,3-葡聚糖水溶性较好,具有抗菌、抗肿瘤、免疫调节以及调节血糖血脂等生物活性和医药活性[2-4],且其生物活性随着分子质量的降低而增高[5]。低分子质量β-1,3-葡聚糖多样的生物活性使其市场需求逐渐增加,如何低成本、绿色环保地生产低分子质量β-1,3-葡聚糖已经成为研究热点。
目前,制备低分子质量β-1,3-葡聚糖主要通过酶法降解[6-7]、物理降解[8]以及化学降解[9-10]等方法,但这些方法存在工艺复杂、化学废液排放以及成本较高等问题[11-13],限制了其大规模工业化生产。近些年来,已有利用微生物共培养发酵获得低分子质量β-1,3-葡聚糖的方法[14-15],但共培养技术工艺较为困难,菌体培养流程较为复杂。王冰等[16]对产热凝胶菌种Agrobacteriumsp.ATCC 31749进行甘油适应性驯化,得到1株可产低分子质量β-1,3-葡聚糖的菌株Agrobacteriumsp.WS-8E3T,但产量仅有1.648 g/L。
发酵过程中的碳源、氮源、pH等因素都会对产量造成影响,本文利用该菌株,在摇瓶水平进行发酵条件的优化,并进行7 L发酵罐的验证实验,为进一步大规模生产低分子质量β-1,3-葡聚糖提供理论基础。
1 实验材料与方法
1.1 实验材料
1.1.1 实验菌种
Agrobacteriumsp.WS-8E3T,保藏于江南大学糖生物制造与生物反应器研究室。
1.1.2 培养基
平板培养基(g/L):甘油100.0,酵母粉10.0,鱼粉蛋白胨2.0,胰蛋白胨2.0,琼脂粉20.0,pH 7.0,121 ℃灭菌20 min。
种子培养基(g/L):甘油40.0,酵母粉6.0,KH2PO41.5,MgSO40.5,pH 7.0,121 ℃灭菌20 min。
基础摇瓶发酵培养基(g/L):甘油100.0,酵母粉6.0,KH2PO43.0,MgSO41.5,(NH4)2SO42.0,pH 7.0,121 ℃灭菌20 min。
所有优化实验均在此基础培养基的基础上进行,7 L发酵罐培养基为优化后的摇瓶发酵培养基。
1.2 主要试剂
甘油、鱼粉蛋白胨、胰蛋白胨、酵母粉、无水乙醇、三氯甲烷、正丁醇、三氟乙酸、甲醇,国药集团化学试剂有限公司;岩藻糖、木糖、阿拉伯糖、鼠李糖、半乳糖、甘露糖,Sigma公司。
1.3 仪器与设备
Y15型Enology全自动葡萄酒分析仪,西班牙Biosystems公司;TU-1810型紫外可见分光光度计,北京普析通用仪器有限责任公司;NEXUS型傅里叶变换红外光谱仪,美国Nicolet公司;ICS5000型离子色谱仪,美国Thermo科技公司;Ultrafle Xtreme型基质辅助激光解析电离串联飞行时间质谱仪,美国Bruck公司;BF115型7 L发酵罐,美国NBS公司。
1.4 实验方法
1.4.1 不同起始甘油浓度对发酵的影响
保持基础摇瓶发酵培养基其他成分不变,起始甘油质量浓度分别设置为20、40、60、100 g/L,30 ℃,200 r/min,发酵96 h,考察碳源浓度对发酵的影响。
1.4.2 甘油补料对发酵的影响
起始甘油质量浓度为20 g/L,分别在36 h补加10或20 g/L甘油,在48 h补加10或20 g/L甘油,考察甘油补料对发酵的影响。
1.4.3 单一氮源对发酵的影响
分别使用酵母粉或硫酸铵作为单一氮源,将起始质量浓度分别设置为1、2、3、4、5、6 g/L,30 ℃,200 r/min,发酵96 h,考察单一氮源对发酵的影响。
1.4.4 复合氮源对发酵的影响
设置酵母粉质量浓度为4 g/L,分别在发酵培养基中添加1、2、3、4、5、6 g/L硫酸铵,30 ℃,200 r/min,发酵96 h,考察复合氮源对发酵的影响。
1.4.5 装液量对发酵的影响
在500 mL三角瓶中将装液量分别设置为25、50、75、100 mL,30 ℃,200 r/min,发酵96 h,考察转速对发酵的影响。
1.4.6 转速对发酵的影响
将摇床转速分别设置为160、180、200、220 r/min,30 ℃,发酵96 h,考察装液量对发酵的影响。
1.4.7 pH对发酵过程的影响
在培养基中分别添加0、1、2、3、4、5 g/L CaCO3,30 ℃,发酵96 h,考察pH对发酵的影响。
1.4.8 低聚糖的提取
将发酵液于4 ℃、8 000 r/min离心20 min,取上清液加3倍体积乙醇,4 ℃过夜后,4 ℃、8 000 r/min离心20 min,取上清液补加3倍体积无水乙醇,4 ℃过夜后,4 ℃、8 000 r/min离心20 min,得到的白色沉淀为低分子质量β-1,3-葡聚糖(以下简称低聚糖)。
1.4.9 糖的纯化
配制Sevag试剂[V(氯仿)∶V(正丁醇)=4∶1],按照V(糖溶液)∶V(Sevag试剂)=3∶1比例,将二者混合振荡30 min,4 ℃,8 000 r/min离心10 min,取上清液重复除蛋白后,采用Sep-Pak C18固相萃取柱(1 g,Waters)再次除蛋白。用紫外分光光度计在200~500 nm进行波长扫描,确保多糖中没有蛋白质残留。
1.4.10 分析方法
1.4.10.1 发酵液中甘油残余量的测定
将2 mL发酵液于4 ℃,8 000 r/min离心10 min后,取上清液用全自动葡萄酒分析仪测定发酵液中的甘油残余量。
1.4.10.2 发酵液中无机氮、伯胺氮残余量的测定
将2 mL发酵液于4 ℃,8 000 r/min离心10 min后,取上清液用全自动葡萄酒分析仪测定发酵液中的无机氮及伯胺氮残余量。
1.4.10.3 生物量的测定
取3 mL发酵液充分混匀后,稀释一定倍数,以未接种的培养基为空白,用紫外分光光度计在600 nm处测定吸光度。另取3 mL发酵液于4 ℃,8 000 r/min离心10 min后,沉淀中加入3 mL 0.5 mol/L的NaOH,充分混匀30 min,4 ℃,8 000 r/min离心10 min后,取沉淀于60 ℃烘干,恒重3次后测干重。测得菌体干重与OD600换算得回归方程为y=0.335 7x-0.251 7。
1.4.10.4 红外光谱分析
产品的结构采用全反射傅里叶变换红外光谱仪(Fourier transform infrared spectrometer, FT-IR),在波数范围400~4 000 cm-1进行扫描,分辨率为0.5 cm-1。
1.4.10.5 单糖组成分析
单糖组分的测定参考LIU等[17]的方法并略作改动:用2 mol/L三氟乙酸溶解5 mg产品,120 ℃金属浴酸解12 h,利用高效离子交换层析色谱柱(CarboPacPA20柱,3 mm×150 mm)结合脉冲电流检测器进行产品单糖组分分析。
1.4.10.6 聚合度分析
利用基质辅助激光解析电离飞行时间质谱(matrix-assisted laser desorption ionization time of flight-mass spectrometer, MALDI-TOF MS)在正离子模式下对产品的质荷比进行分析[18]。
1.4.11 数据处理
采用Origin 8.5软件绘制图表并进行数据分析。
2 结果与讨论
2.1 培养基组分优化
2.1.1 甘油浓度对发酵的影响
碳源浓度对发酵至关重要,过低或过高都不利于菌体生长及产物累积。甘油是具有高渗作用的物质,作为碳源使用时,浓度过高带来的高渗环境可能会抑制菌体生长及代谢产物的累积[19]。
由图1和表1可知,当起始甘油质量浓度分别为60、100 g/L时,0~24 h内OD600平均上升速率及甘油平均消耗速率较低,这直接导致了最终低聚糖的产量较低(分别为2.010和1.648 g/L)。而当起始甘油质量浓度分别为20及40 g/L时,0~24 h内OD600平均上升速率及甘油平均消耗速率相对较高,最终低聚糖的产量较高(分别为2.210、2.421 g/L)。这可能是因为甘油浓度过高时,菌体产生的能量要用于抵御外界高渗环境对自身的迫害[20],同时用于合成葡聚糖前体物质,导致能量因子多段消耗,菌株的生长和产糖能力因此降低[16]。从图1及表1可见,20 g/L甘油起始质量浓度,无论是OD值的上升速率还是甘油的利用速率都高于40 g/L甘油起始质量浓度。但是甘油起始质量浓度为20 g/L时,发酵液中的甘油残余量在36 h降低到8 g/L,48 h已低至6 g/L。综上,20 g/L甘油前期对菌体生长更有利。

a-生物量(OD600);b-甘油消耗情况图1 不同甘油浓度对生物量及甘油消耗的影响Fig.1 Biomassand consumption of glycerol under different glycerol concentration
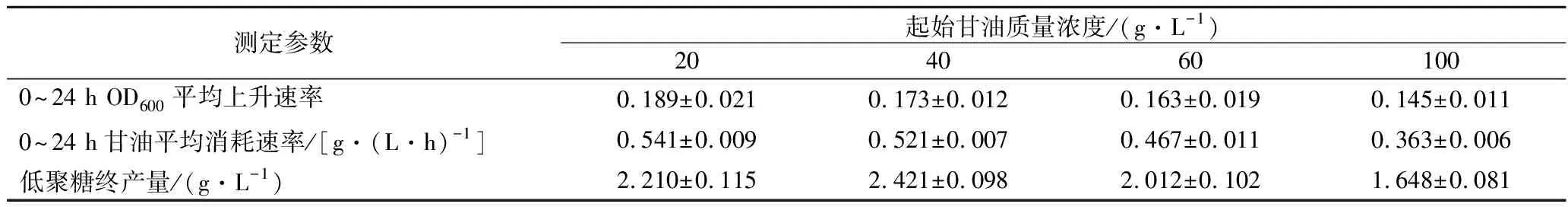
表1 不同甘油浓度下的生物量(OD600)增长速率、甘油消耗速率及低聚糖产量Table 1 Biomass(OD600)growth rate, glycerol consumption rate and yield at different concentration of glycerol
2.1.2 甘油补料对发酵的影响
由甘油浓度实验可知,将初始甘油质量浓度设置为20 g/L时,菌体前期生长速率最快,因此将初始甘油质量浓度设置为20 g/L,分别采用在发酵36或48 h补加甘油的策略,结果如表2所示。
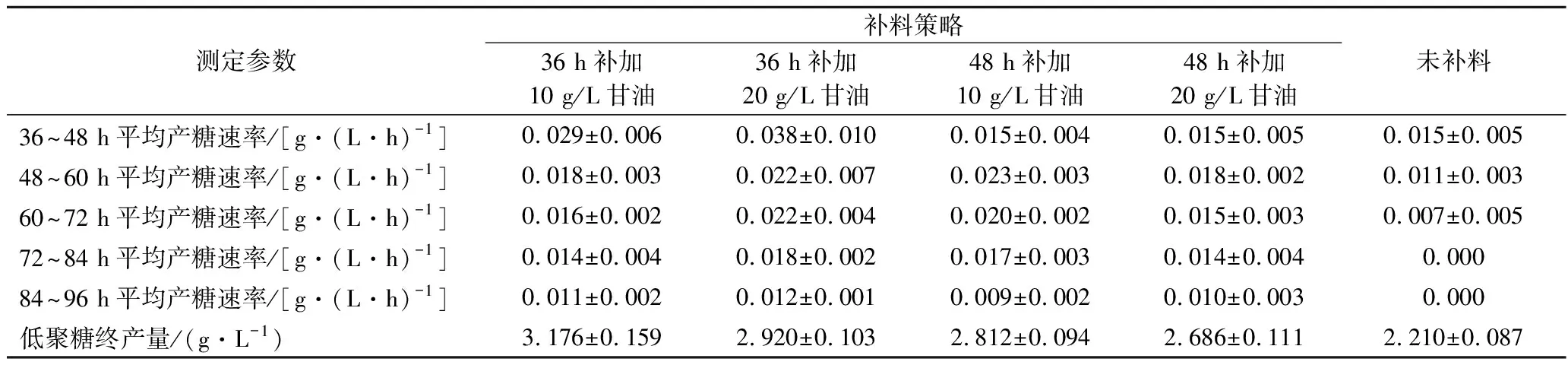
表2 不同甘油补料策略下的产糖速率及最终低聚糖产量Table 2 Rate of oligosaccharide production and yield at different supplementation strategies of glycerol
发酵36 h分别向发酵液中补加10或20 g/L甘油,36~48 h产糖速率与未补料相比均有不同程度的提升,分别为0.029以及0.038 g/(L·h)。发酵结束低聚糖产量分别为3.176及2.92 g/L。发酵48 h分别向发酵液中补加10或20 g/L甘油,48~60 h产糖速率分别为0.018及0.023 g/(L·h),最终产量分别为2.812及2.686 g/L,皆低于在36 h补加甘油。分析原因可能是48 h时体系中甘油残余量极低,对菌体的正向推动力减弱,产糖能力降低,补加甘油导致菌体不耐受环境中突然增高的渗透压,因此在48 h时补加甘油不利于菌体生长及低聚糖的累积。
2.1.3 单一氮源对发酵的影响
通常情况下,氮源主要在发酵前期用于菌体生长,构成细胞物质及含氮代谢产物,发酵中后期菌体需要在限制氮源或维持较高C/N的条件下产胞外多糖[21]。但菌体在反复、长时间驯化过程中,微生物许多生理机制也会发生相应的变化以帮助其适应外界环境的变化[22-23]。研究不同氮源及浓度对驯化菌的发酵影响是十分必要的。
分别利用酵母粉或硫酸铵作为单一氮源,分别设置初始质量浓度为1、2、3、4、5、6 g/L,研究单一氮源对发酵的影响。结果如图2及表3所示,当酵母粉作为单一氮源时,随着酵母粉质量浓度的增高,生物量与产量皆提高,伯胺氮消耗速率越快,氮源耗尽后,菌体会产水不溶性胶体,酵母粉为4 g/L时,低聚糖产量为1.603 g/L,当酵母粉质量浓度继续增高时,产量的提高不再明显,因此,以4 g/L酵母粉添加量较好。当以硫酸铵作为单一氮源时,随着硫酸铵质量浓度提高,生物量与产量皆提高,但相较于使用酵母粉作为单一氮源时,无论是菌体的生物量还是低聚糖的产量都整体偏低。

a, b-酵母粉作为单一氮源时的生物量(OD600)及伯胺氮消耗情况;c, d-硫酸铵作为单一氮源时的生物量(OD600)及无机氮消耗情况图2 单一氮源时生物量及氮源消耗情况Fig.2 Biomass and consumption of nitrogen at different single nitrogen source
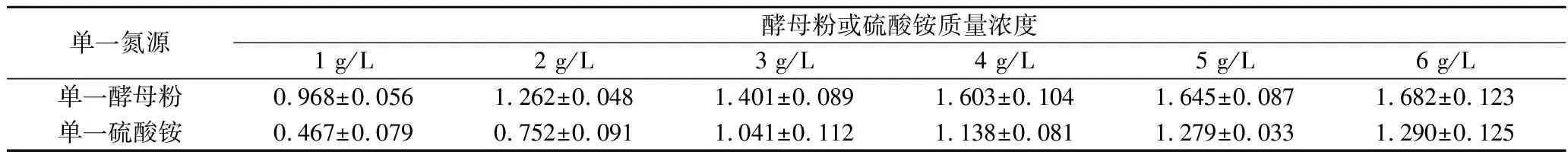
表3 单一氮源时低聚糖产量 单位:g/L
2.1.4 复合氮源对发酵的影响
根据前述实验结果,选择添加4 g/L酵母粉作为有机氮源,分别添加1、2、3、4、5、6 g/L硫酸铵,考察复合氮源对发酵的影响,结果如图3及表4所示。当硫酸铵为1 g/L时,菌体产生了少量的低聚糖并在氮源耗尽后产水不溶性胶体。当硫酸铵质量浓度为3 g/L时,菌体缓慢消耗无机氮并产低聚糖,产量达到2.612 g/L且不产水不溶性胶体。但当发酵体系中硫酸铵浓度进一步增加时,菌体的生长和产低聚糖能力都降低。综上,选择4 g/L酵母粉,3 g/L硫酸铵作为初始复合氮源较益。

表4 复合氮源时低聚糖产量 单位:g/L
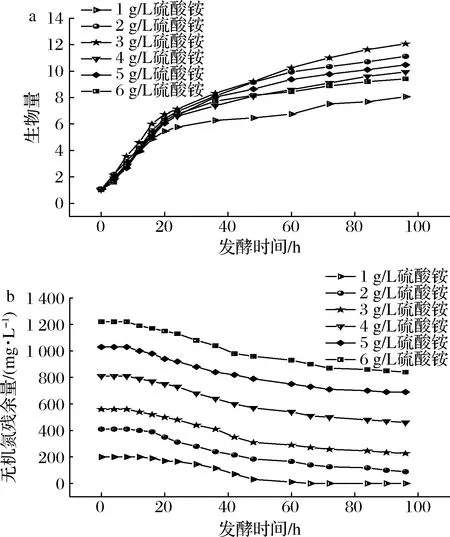
a-生物量(OD600);b-无机氮消耗情况图3 复合氮源时的生物量及无机氮消耗情况Fig.3 Biomass and consumption of nitrogen at compound nitrogen source
2.2 装液量对发酵的影响
以甘油为碳源合成低分子质量β-1,3-葡聚糖属于高耗能过程[24],改善发酵体系中的溶氧水平可有效促进三羧酸循环合成ATP,为菌体生长及产糖提供能量[25]。摇瓶实验中,装液量是溶氧的直接影响因素。为考察溶氧对发酵的影响,在500 mL摇瓶中将装液量分别设置为25、50、75、100 mL,考察装液量对发酵的影响,结果如图4所示。装液量从25 mL增加到50 mL时,菌体浓度及低聚糖产量几乎相当,当装液量为75 mL时,低聚糖产量显著提高,为2.636 g/L。当装液量继续增加至100 mL时,低聚糖产量随之降低。这是因为高溶氧时菌体对底物的利用速率较快,后期底物浓度偏低,不利于产物的累积;而高装液量会限制摇瓶中的氧气扩散,使发酵体系中溶氧水平降低,从而限制了菌体的生长与产物的累积。因此,装液量为75 mL/500 mL时的溶氧可能是该菌株的较佳溶氧水平。
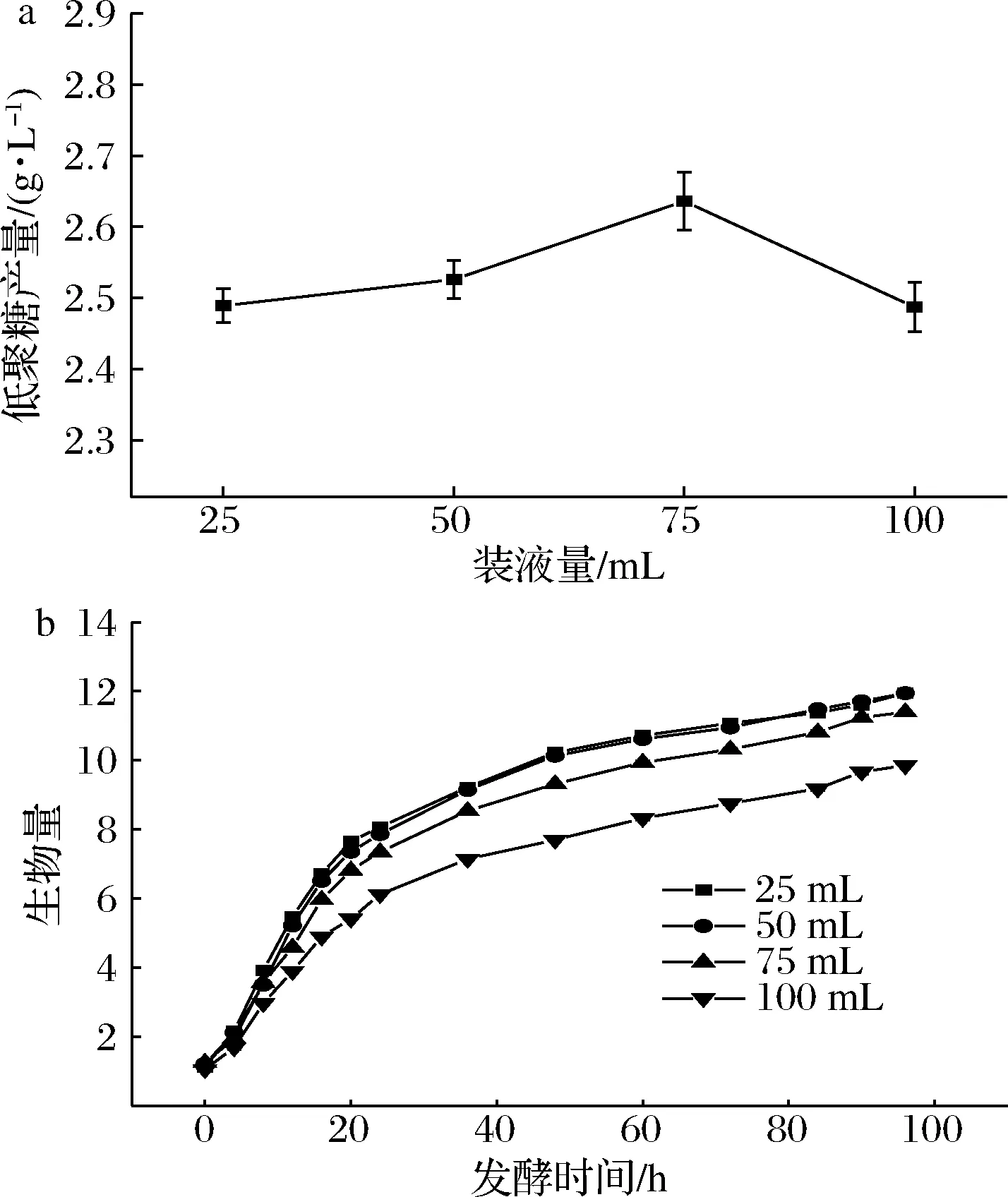
a-低聚糖产量;b-生物量(OD600)图4 不同装液量的产量及生物量Fig.4 Yield and biomass at different medium volume
2.3 pH对发酵的影响
发酵是一种多酶复合反应体系,体系中的pH值影响菌体细胞内各种酶的活性进而影响菌体的生长和产物的合成。多数微生物生长繁殖所需pH与产物累积所需pH不同,两阶段pH控制策略在发酵生产微生物多糖中被广泛利用[26-27]。在发酵过程中分别设置CaCO3质量浓度为0、1、2、3、4、5 g/L,以维持不同pH值,考察发酵过程中pH的变化与菌体生长、产物累积之间的关系。发酵前24 h各组菌体生长OD600值与pH值几乎相同,24 h以后各组OD600值、pH值及低聚糖产量出现了差异(数据未显示),其中添加2 g/L CaCO3时的低聚糖产量最高,为2.510 g/L(图5-a)。添加2 g/L CaCO3时,菌体在发酵过程中产糖与pH的关系如图5-b所示。在24~36 h ,pH为6.5~6.3时,产糖速率最快,为0.060 g/(L·h)。综上所述,菌体的最佳产糖pH可能为6.3左右。
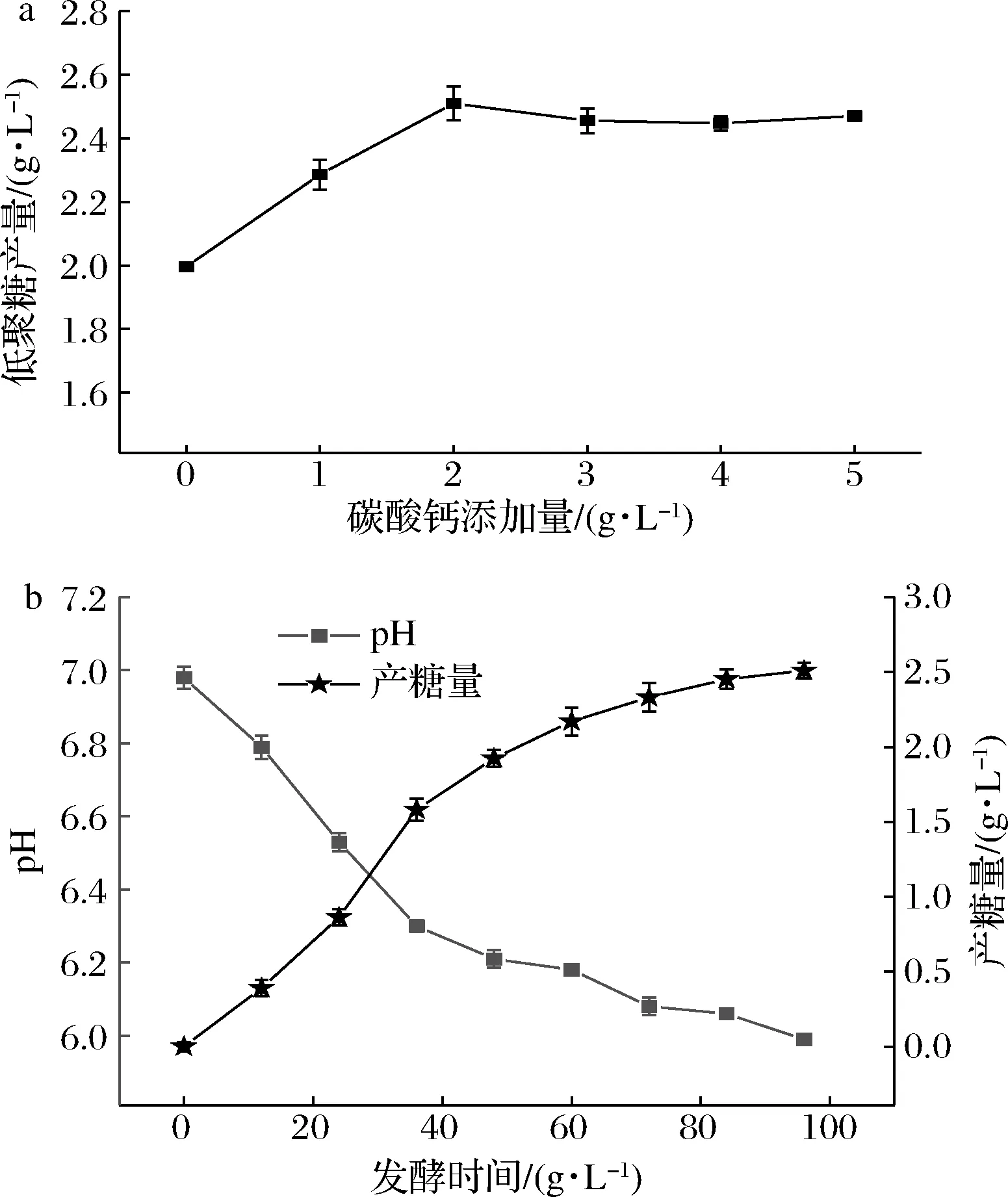
a-低聚糖终产量; b-添加2 g/L CaCO3时发酵过程中产糖量与pH变化图5 不同pH下产糖情况Fig.5 Yield at different pH
2.4 7 L发酵罐验证实验
在7 L发酵罐上进行摇瓶实验结果的初步验证。初始甘油质量浓度为20 g/L,发酵液甘油残量为10 g/L时补加甘油;以4 g/L酵母粉以及3 g/L硫酸铵作为复合氮源;菌体生长阶段0~24 h控制pH为7,菌体产糖阶段24~96 h控制pH为6.3;维持通气比2 vvm-1;搅拌速率150 r/min;发酵96 h。
如图6所示,发酵0~24 h为菌体生长阶段,此阶段菌体快速生长,OD600平均上升速率为0.395;发酵24~36 h已有少量低聚糖产生,产糖速率为0.066 g/(L·h);36~48 h产糖速率显著提升,为0.117 g/(L·h);发酵48~60 h,产糖速率为0.119 g/(L·h);60 h后,产糖速率随发酵时间的延长逐渐减缓直到96 h发酵结束,低聚糖的产量达到8.03 g/L,结果表明,生长期甘油质量浓度为20 g/L,产糖期甘油浓度为30 g/L并控制产糖期pH为6.3,可大幅度提高低聚糖产量。
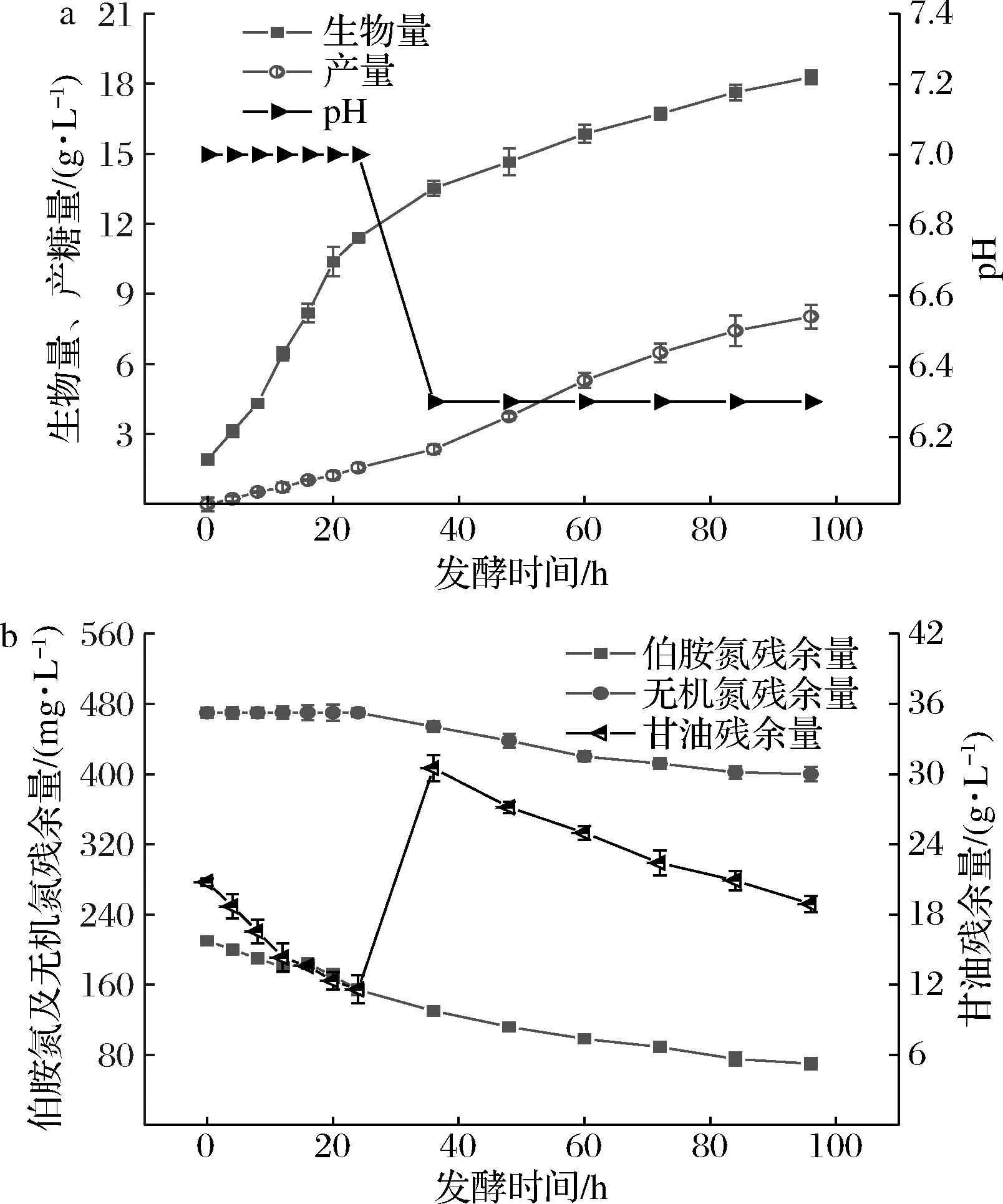
a-生物量(OD600)、产糖量及pH变化; b-甘油、无机氮以及伯胺氮残余量图6 7 L发酵罐发酵过程中参数变化Fig.6 Changes of parameters during the fermentation in a 7 L fermenter
2.5 产品结构解析
对7 L发酵罐所得产品进行结构分析。产品的红外光谱特征吸收峰与热凝胶标准品一致(如图7-a所示),样品在波数891.2 cm-1处有典型的β-糖苷键特征吸收峰,而在840和、790 cm-1处无β-糖苷键的特征吸收峰,另外在波数为3 340.4、1 640.5、2 900.1 cm-1处有多糖特征吸收峰,1 030.7、1 160.4 cm-1处有吡喃型糖环构型的特征吸收峰。离子色谱单糖组成分析结果如图7-b所示,样品中只含有一组峰,其保留时间与标准单糖混合液中葡萄糖的保留时间一致,说明产品仅由葡萄糖组成。MALDI-TOF MS聚合度分析结果如图7-c所示,出现了m/z为2 956.8、3 118.8、3 280.9、3 442.9、3 605.1的峰,分别对应聚合度为18,19,20,21和22。优化后的产品为低分子质量β-1,3-葡聚糖,聚合度在18~22。其结构特征与优化前摇瓶发酵产物结构一致(详见参考文献[16])。

a-FT-IR;b-单糖组成分析(1~7分别是岩藻糖、鼠李糖、阿拉伯糖、半乳糖、葡萄糖、木糖、果糖);c-MALDI-TOF MS图7 低聚糖的结构及聚合度分析Fig.7 Analysis of structure and degree of polymerization of oligosaccharides
3 结论
以甘油为碳源,发酵驯化菌Agrobacteriumsp.WS-8E3T产β-1,3-低聚糖,分别进行了碳源、氮源初始浓度的优化,并在摇瓶水平进行补料发酵的初步探索,优化得到起始甘油质量浓度为20 g/L,并在低于10 g/L时补加20 g/L,确定最佳酵母粉与硫酸铵的初始质量浓度分别为4和3 g/L,最佳装液比为75 mL/500 mL,产糖期最适pH为6.3左右。通过7 L发酵罐的验证实验,最终低聚糖产量达到8.03 g/L,相比于未优化前提高386.7%,通过结构鉴定,判定该低聚糖为β-1,3-葡聚糖,聚合度范围在18~22。
猜你喜欢
食品科学(2022年13期)2022-07-29
中国调味品(2022年6期)2022-06-05
阅读(快乐英语中年级)(2022年5期)2022-06-03
食用菌(2022年1期)2022-02-14
乡村科技(2021年12期)2021-09-06
农产品加工(2020年20期)2020-11-14
食品科学(2020年17期)2020-09-21
江西水产科技(2019年3期)2019-09-10
天津农业科学(2019年4期)2019-05-31
中国糖料(2019年1期)2019-02-13
