
小麦隐性核不育保持系表达载体的构建及拟南芥遗传转化
2022-08-09 01:14李紫良张建朝冯小雨宋瑜龙马守才刘东涛王军卫
麦类作物学报 2022年6期
高 聪,李紫良,胡 甘,张建朝,冯小雨,牛 娜,宋瑜龙,马守才,刘东涛,王军卫
(1.西北农林科技大学农学院/小麦育种教育部工程研究中心/陕西省作物杂种优势研究与利用重点实验室,陕西杨凌 712100;2.江苏徐淮地区徐州农业科学研究所,江苏徐州 221131)
近几十年来,杂种优势利用在玉米(ZeamaysL.)、水稻(OryzasativaL.)和油菜(BrassicanapusL.)生产上已取得了举世瞩目的成就[1-3],而杂交小麦却一直未能取得重要突破。小麦是全世界主要粮食作物之一,是全球约30%人口的主要食物来源。全球人口的持续增长以及人们生活质量的提高,对小麦产量和品质的要求也随之提高。从目前来看,创制杂交小麦仍然是提高小麦产量和品质最有效的途径之一,并且对世界粮食生产和安全具有举足轻重的影响[4]。
小麦是严格的自花授粉作物,小麦杂种优势的利用严格依赖于小麦雄性不育系的选育。但是目前采用的细胞质雄性不育系、光温敏雄性不育系以及化学杂交剂诱导的生理型雄性不育系分别存在不育恢复机理复杂、对环境因素影响敏感以及化学杂交剂药效不稳定等缺点,不能满足小麦生产发展的需要[5-7]。与其他方法相比,由核不育控制的“二系”杂种优势利用技术,能够克服了细胞质雄性不育三系法和光温敏雄性不育两系法的缺陷,且兼具细胞质雄性不育三系法的稳定性和光温敏雄性不育两系法配组灵活性的优点,具有重要的市场利用价值[8]。目前已经发现的核不育基因有ms1、ms5、Ms2、Ms3和Ms4,其中ms1和ms5是隐性突变体,Ms2、Ms3和Ms4是显性突变体[9-13]。然而,利用常规的育种方法很难将植物的隐性不育基因有效地保持和利用起来。因此,培育不受环境影响且可自主保持、繁殖的单隐性核不育系已成为雄性不育系选育技术的难点。
2017年,第一个隐性核不育基因ms1被克隆,其等位恢复基因Ms1基因只在禾本科植物的小孢子和花药中表达,该基因突变导致花药中长链脂肪酸积累和花粉外壁结构被破坏,进而引起花粉败育[14]。该基因的克隆为通过基因工程创制雄性不育系和规模化制备杂交小麦种子提供了可能。近年来,美国杜邦公司开发了SPT技术,通过利用隐性核不育基因ms45,实现了玉米雄性不育的保持和杂种优势利用[15]。SPT技术是指将育性恢复基因Ms45、花药特异性表达启动子PG47、玉米α淀粉酶基因ZmAA1、红色荧光蛋白基因DsRed2及特异性启动子和终止子序列一起连接到表达载体上,转化到Ms45基因突变 (ms45/ms45)导致的核不育玉米中,T1代中基因型为Ms45-ZmAA1-DsRed2/ms45。由于α淀粉酶基因ZmAA1的表达导致携带Ms45-ZmAA1-DsRed2的花粉致死,T1代自交时只能产生基因型ms45的花粉,产生的种子基因型为带转基因的Ms45-ZmAA1-DsRed2/ms45和不带转基因ms45/ms45,2种基因型各占50%,通过观察红色荧光区分2种种子。这样生产的不育系 (ms45/ms45)能够保持且不携带转基因,保持系(Ms45-ZmAA1-DsRed2/ms45)则能够自我繁殖。Chang等[16]发现,水稻OsNP1的突变会产生雄性不育的隐性核不育OsNP1基因,并利用该基因并结合SPT技术,开展了水稻的杂种优势利用工作。
本试验以SPT技术为依托,利用表达载体pGEII能够携带较大的外源目的基因且可在转基因植物中高效表达的特性,通过无缝克隆技术和同源重组酶,将花粉致死基因ZmAA1、小麦花粉育性恢复基因Ms1、红色荧光蛋白基因DsRed2及其特异性启动子和终止子序列构建植物表达载体pGEII-Ms1-ZmAA1-DsRed2,并转化野生型拟南芥,验证表达载体构建是否成功,以期为创制可以保持和繁殖的新型不育系材料,规避育种中转基因生物安全问题提供理论依据。
1 材料与方法
1.1 试验材料
供试小麦品种中国春、植物表达载体pGEII和农杆菌菌株GV3101均由西北农林科技大学作物杂种优势研究与利用课题组提供,pGEII携带有草铵膦抗性,玉米自交系B73为西北农林科技大学农学院郭东伟副教授惠赠,载体pCAMBIA3300-Ltp2-DsRed2为吉林省农业科学院刘文国研究员惠赠。
1.2 DNA、总RNA的提取与cDNA的合成
将小麦品种中国春在人工气候室(温度10 ℃,相对湿度50%,光照/黑暗16 h/8 h,光照强度18 000 lx)培养至减数分裂期,取幼穗使用TRNzol-A+试剂盒(天根,北京)提取总RNA,取叶片使用CTAB法提取DNA[17]。将玉米自交系B73种子在人工气候室(条件同上)培养,萌发3 d时取胚乳使用TransZol Plant试剂盒(全式金,北京)提取总RNA,植株生长2周时取叶片提取DNA。用PrimeScriptTMRT reagent Kit with gDNA Eraser(Takara,北京)将RNA反转录获得cDNA。
1.3 隐性核不育恢复基因 Ms1表达盒pGEM3ZEVL-Ms1的构建
以Ms1-P-F/R为引物(表1),以中国春DNA为模板,扩增育性恢复基因Ms1的启动子,暂命名为Ms1-P(GenBank No.MG273685.1);以Ms1-F/R为引物(表1),以中国春cDNA为模板,扩增Ms1基因的编码区序列(GenBank No.MG273685.1)[14];以F8-1-F/R为引物(表1),以中国春DNA为模板,扩增小麦花粉特异表达基因F8-1的终止子(GenBank No.KP878480.1)[18]。以上序列扩增均使用KOD-Plus-Neo高保真酶(TOYOBO,日本)进行。随后,使用内切酶EcoR V酶切pGEM3ZEVL载体。使用BM无缝克隆试剂盒(博迈德,北京)将扩增得到的Ms1和F8-1终止子连接到线性化pGEM3ZEVL载体上,构建重组载体。以此载体为模板,使用引物MF-F/R(表1)扩增得到带有同源序列的Ms1-F8-1片段。使用BM无缝克隆试剂盒将Ms1-P和Ms1-F8-1连接,构建pGEM3ZEVL-Ms1载体,完成核不育恢复基因Ms1表达盒的构建。
1.4 花粉致死基因表达盒pGEII-ZmAA1-DsRed2的构建
以PG47-F/R为引物(表1),以玉米自交系B73的DNA为模板,扩增启动子PG47(GenBank No.X66692.1);以ZmAA1-F/R为引物(表1),以玉米自交系B73的 cDNA为模板,扩增花粉致死基因ZmAA1的编码区序列(GenBank No.NM_001176140.1);以IN2-1-F/R为引物(表1),以玉米自交系B73的DNA为模板,扩增终止子IN2-1(GenBank No.BT040284.1);以引物DsRed2-F/R为引物(表1),以载体pCAMBIA3300-Ltp2-DsRed2为模板,扩增DsRed2-box,包含启动子Ltp2(GenBank No.X69793.1)、DsRed2(GenBank No.KY379159.1)、NOS终止子。以上序列扩增均使用KOD-Plus-Neo高保真酶进行。用BM无缝克隆试剂盒将ZmAA1和IN2-1连接到线性化pGEM3ZEVL载体上,得到重组载体pGEM3ZEVL-ZmAA1-IN2-1。以此重组载体为模板,用引物ZmAA1-F/R-1(表1)扩增得到ZmAA1-IN2-1片段。用BM无缝克隆试剂盒将ZmAA1-IN2-1和DsRed2-box连接到pGEM3ZEVL载体,得到重组载体pGEM3ZEVL-ZmAA1-IN2-1-DsRed2-box。随后以此重组载体为模板,用引物ZID-F/R(表1)扩增得到带有同源序列的ZmAA1-IN2-1-DsRed2-box,用EcoR I和XhoI双酶切pGEII载体,将启动子PG47和ZmAA1-IN2-1-DsRed2-box连接到表达载体pGEII上,得到重组载体pGEII-ZmAA1-DsRed2,完成花粉致死基因ZmAA1表达盒的构建,其中包含红色荧光蛋白基因DsRed2。
1.5 小麦隐性核不育保持系表达载体pGEII-Ms1-ZmAA1-DsRed2的构建
以构建的Ms1表达盒pGEM3ZEVL-Ms1载体为模板,用引物Ms1-F1/R1扩增得到带有同源序列的MS1-P-Ms1-F8-1片段。用EcoR I酶切pGEII-ZmAA1-DsRed2载体,随后通过BM无缝克隆试剂盒将MS1-P-Ms1-F8-1连接到线性化pGEII-ZmAA1-DsReds载体上,得到重组表达载体pGEII-Ms1-ZmAA1-DsRed2,完成小麦隐性核不育保持系表达载体的构建。
1.6 pGEII-Ms1-ZmAA1-DsRed2表达载体的遗传转化
采用农杆菌介导的方法[19],将构建好的植物表达载体pGEII-Ms1-ZmAA1-DsRed2转入野生型拟南芥中。培养成熟后,收获T0代种子并继续种植,待发芽一周后,喷洒5 mg·L-1的除草剂草铵膦进行抗性筛选,生长7 d左右后,除去萎蔫、枯死以及生长缓慢的植株,将抗性植株继续培养并收获T1代种子。
1.7 转基因拟南芥的PCR检测及种子荧光观察
提取T1代抗性植株的叶片DNA,设计引物ZmAA1-F2/R2(表1)对抗除草剂 T1代转基因植株进行PCR检测。以pGEII-Ms1-ZmAA1-DsRed2载体作为阳性对照,野生型拟南芥叶片DNA作为阴性对照,以提取的T1代转基因拟南芥基因组 DNA为模板进行PCR检测。使用德国ZEISS生物荧光显微镜观察转基因种子的红色荧光。

表1 本研究所用的引物Table 1 Primers used in the study
2 结果与分析
2.1 隐性核不育恢复基因 Ms1表达盒pGEM3ZEVL- Ms1的构建结果
以Ms1-P-F/R、F8-1-F/R为引物,以中国春DNA为模板,扩增隐性核不育基因Ms1的启动子Ms1-P(图1A)和小麦花粉特异表达基因F8-1终止子,分别得到2 670和245 bp的特异条带(图1C);以Ms1-F/R为引物,以中国春cDNA为模板,扩增育性恢复基因Ms1,得到663 bp的特异条带(图1B)。使用BM无缝克隆试剂盒先把育性恢复基因Ms1和F8-1终止子连接起来,然后将MS1-P与Ms1-F8-1连接在一起,经测序验证与目的序列一致,得到隐性核恢复基因Ms1表达盒pGEM3ZEVL-Ms1。
2.2 花粉致死基因表达盒pGEII-ZmAA1-DsRed2的构建结果
分别以PG47-P-F/R和IN2-1-F/R为引物,以玉米B73的DNA为模板,分别扩增启动子PG47和终止子IN2-1,分别得到2 873和344 bp的特异条带(图2A);以ZmAA1-F/R为引物,以玉米B73的cDNA为模板,扩增花粉致死基因ZmAA1,得到1 346 bp的特异条带(图2A);以DsRed2-F/R为引物,以载体pCAMBIA3300-Ltp2-DsRed2为模板,扩增红色荧光蛋白基因DsRed2-box,得到2 153 bp的特异条带(图2B)。用BM无缝克隆试剂盒先把ZmAA1基因和IN2-1终止子连接起来,随后把ZmAA1-IN2-1与DsRed2-box连接在一起。用EcoR I和XhoI双酶切pGEII载体,把PG47启动子和ZmAA1-IN2-1-DsRed2连接到表达载体上,得到花粉致死基因ZmAA1表达盒重组载体pGEII-ZmAA1-DsRed2。
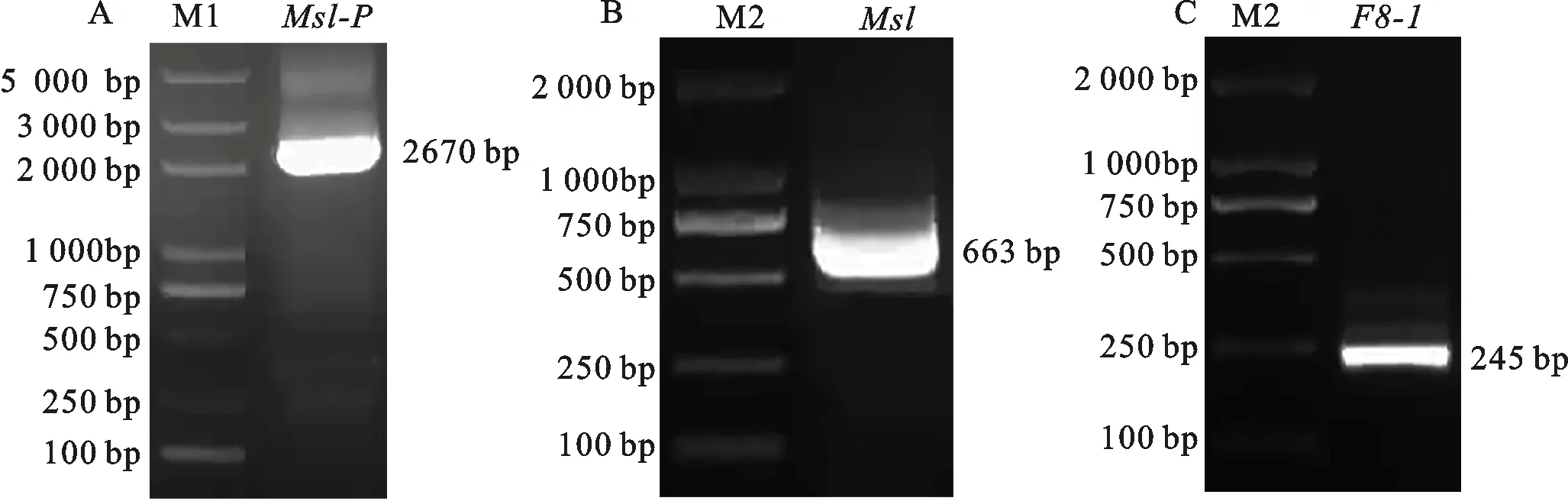
M1:DL5000。图2和图3同。M2:DL2000。图5同。
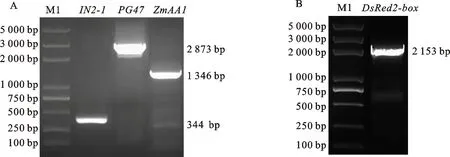
图2 启动子 PG47、玉米花粉致死基因 ZmAA1、终止子 IN2-1和红色荧光蛋白基因 DsRed2-box的克隆结果Fig.2 Cloning results of PG47 promoter,maize anther lethal gene ZmAA1,IN2-1 terminator and DsRed2-box
2.3 小麦隐性核不育保持系表达载体pGEII-Ms1-ZmAA1-DsRed2的构建结果
以构建的Ms1表达盒pGEM3ZEVL-Ms1载体为模板,用引物Ms1-F1/R1扩增得到带有同源序列的Ms1-P-Ms1-F8-1。用EcoR I酶切pGEII-ZmAA1-DsRed2载体,随后通过BM无缝克隆试剂盒将Ms1-P-Ms1-F8-1连接到线性化载体上,得到重组表达载体pGEII-Ms1-ZmAA1-DsRed2。用AscI和XhoI对重组载体进行双酶切鉴定,这两种酶的酶切位点在载体中都是唯一的,酶切后得到一条3 814 bp的条带(图3),与目标条带一致。经测序验证,重组载体pGEII-Ms1-ZmAA1-DsRed2与目的序列一致,完成了小麦隐性核不育保持系表达载体pGEII-Ms1-ZmAA1-DsRed2的构建(图4)。
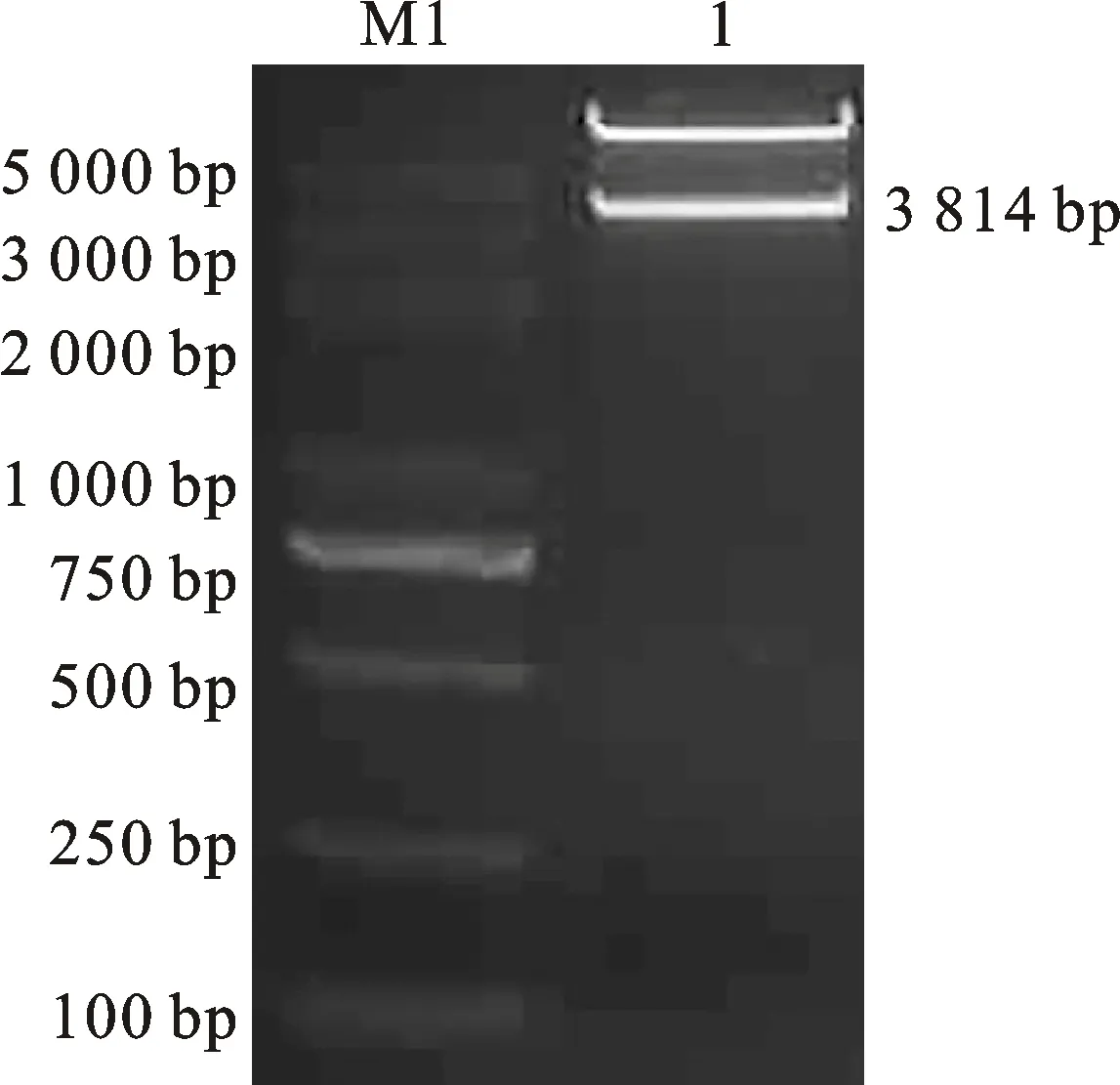
1:用Asc I和Xho I对重组载体pGEII-Ms1-ZmAA1-DsRed2的双酶切结果。
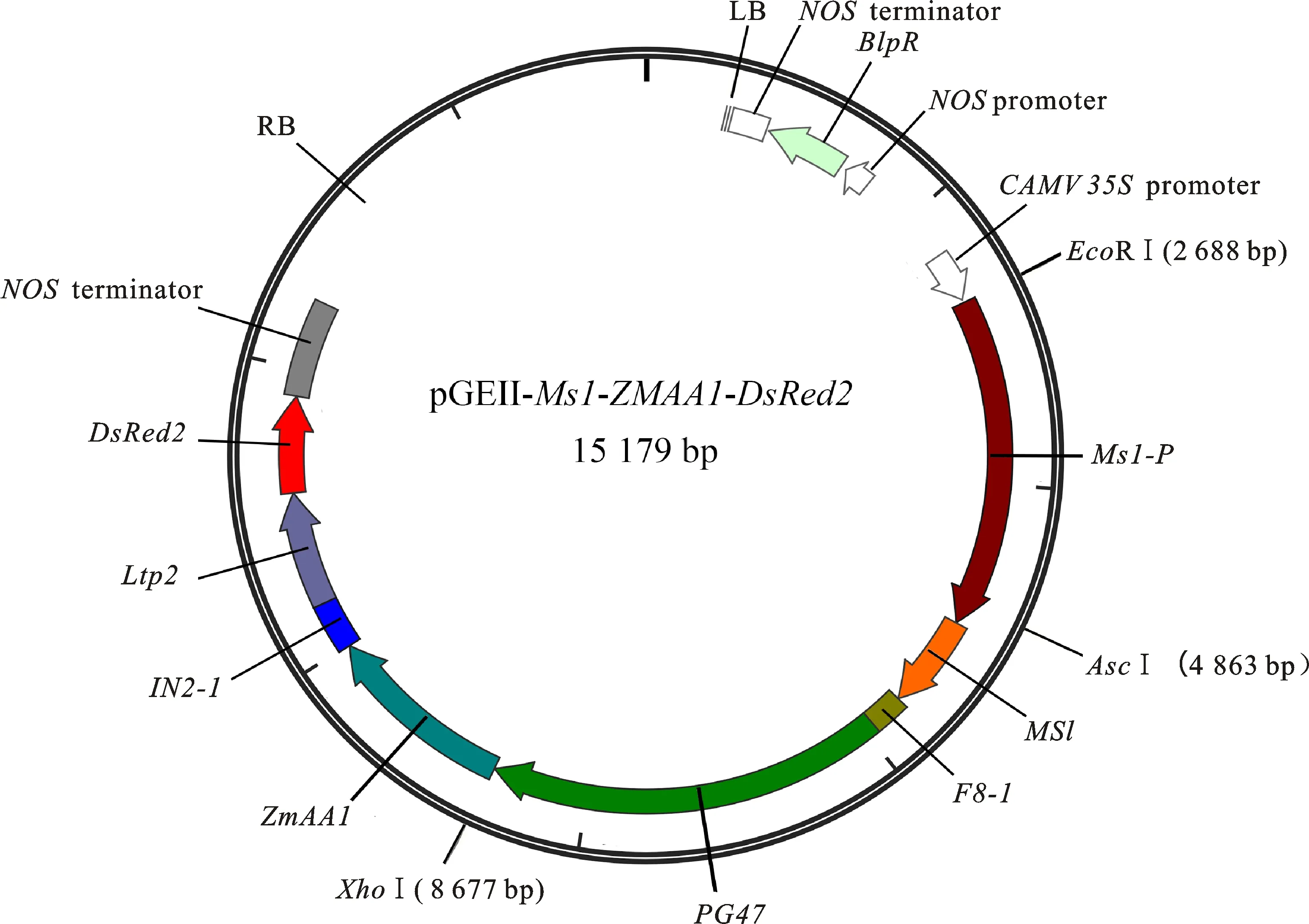
图4 pGEII-Ms1-ZmAA1-DsRed2的载体图谱Fig.4 Map of pGEII-Ms1-ZmAA1-DsRed2 vector
2.4 pGEII-Ms1-ZmAA1-DsRed2表达载体的遗传转化结果
将重组质粒pGEII-Ms1-ZmAA1-DsRed2转化到农杆菌GV3101中,使用农杆菌转化法转化野生型拟南芥。经过除草剂草铵膦筛选,发现有9株T1代抗性拟南芥成活。提取T1代抗性拟南芥叶片DNA,用目的基因ZmAA1的特异引物ZmAA1-F2/R2进行PCR检测,以野生型拟南芥DNA为阴性对照,以重组质粒pGEII-Ms1-ZmAA1-DsRed2为阳性对照。经PCR检测,发现有6株抗性拟南芥的DNA能够扩增出目的条带(图5),说明T1代共获得6株转基因阳性拟 南芥。
2.5 转基因拟南芥种子的鉴定结果
使用荧光显微镜在红色荧光滤光片下观察野生型拟南芥种子和T1代转基因阳性拟南芥种子,发现野生型拟南芥的种子表面平滑,不能发出红色荧光信号;而转基因种子表面呈颗粒状,会发出红色荧光信号(图6),说明红色荧光蛋白基因DsRed2在转基因拟南芥种子中得到表达。
3 讨 论
杂种优势利用是提高小麦产量和品质最有效的途径之一,对世界的粮食安全保障具有重要的意义[20]。尽管目前利用三系、光温敏和化学杂交剂诱导的杂交种已在生产上得到应用,但是由于制种复杂、不育系不稳定以及药剂残留等问题,导致制种成本高,质量难以保障[15]。因此,需要采用新的途径,来提高杂交小麦种子质量以及降低制种成本,并且能够快速选育杂交种,这是目前小麦生产上急需解决的问题。现有的蓝标型隐性核不育杂交小麦生产技术体系虽然可以通过颜色分辨不育系种子(白色)和保持系种子(浅蓝色),但是同时也产生了纯合育性正常的深蓝粒种子,而目前的色选机仅能够做到区分蓝、白粒,不能区分浅蓝、深蓝粒,这就造成了随着繁殖年代的增加,不育系的繁殖效率显著下降[21]。SPT技术的发明,建立了一种新型杂交种子生产技术体系,通过基因工程获得的隐性核不育保持系自交产生的种子中,50%是不带荧光的不育系种子,50%是带荧光的保持系种子,采用色选技术很容易区分保持系与不育系种子,并且不育系种子中不含转基因成分。因此,利用该体系不仅可以提高制种纯度、降低制种成本,而且生产的不育系种子中不含外源基因,确保了生物安全[22]。这就避免了蓝标型隐性核不育杂交小麦生产技术体系深蓝粒筛选产生的问题。Chang等[16]利用SPT技术,将 1 200份水稻恢复系和不育系杂交,发现约85%的F1植株单株产量优于亲本,表明该技术在水稻的杂交育种和生产中具有良好的应用前景。
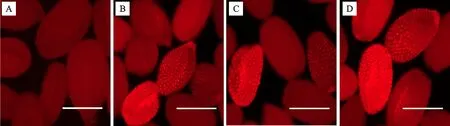
A:野生型拟南芥种子;B~D:转基因拟南芥种子。比例尺=0.2 mm。
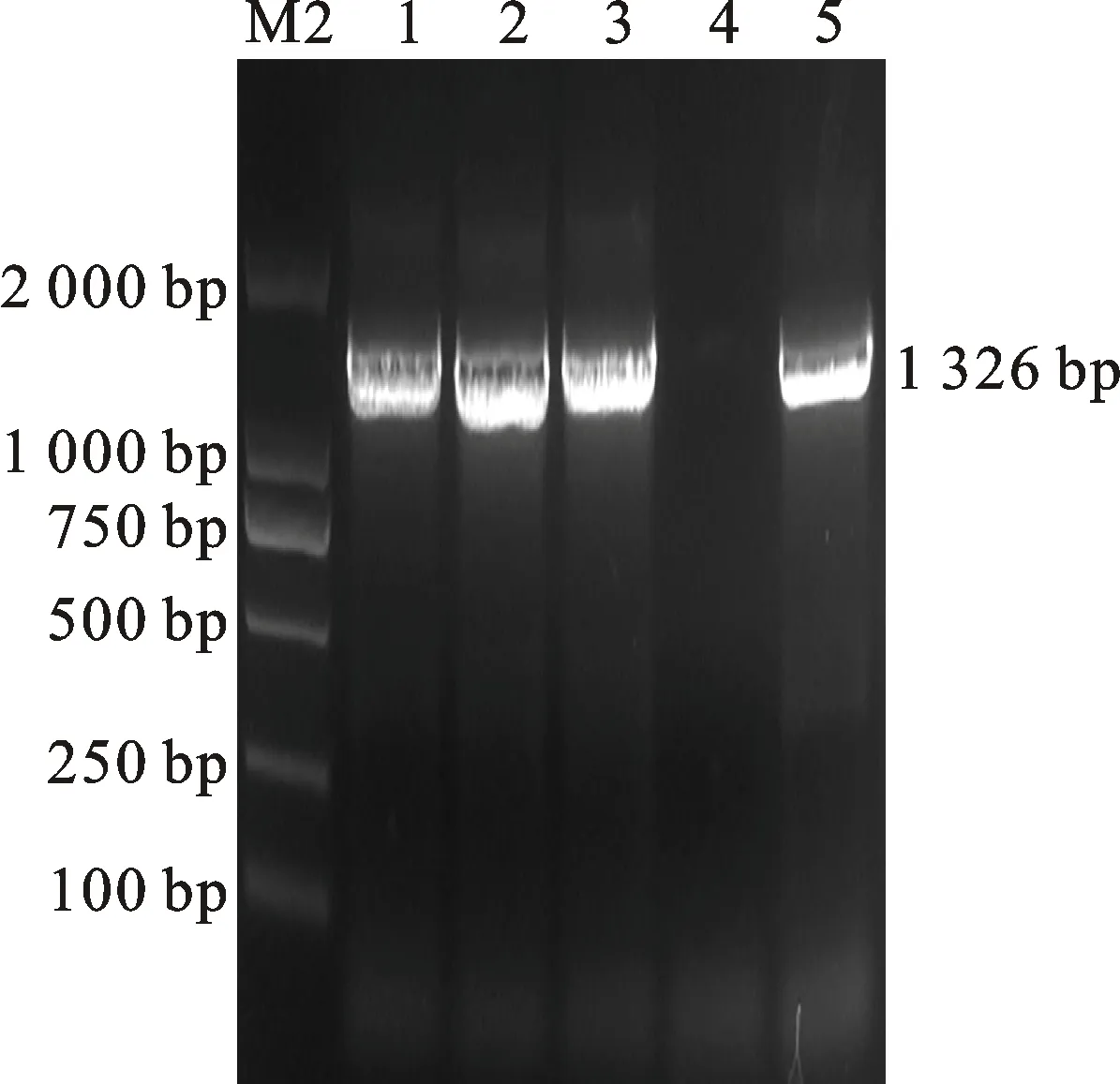
1~3:转基因拟南芥;4:阴性对照;5:阳性对照。
本研究借鉴SPT技术经验,将小麦花粉育性恢复基因Ms1、花粉致死基因ZmAA1、荧光筛选基因DsRed2及其特异性花药启动子和终止子序列构建了小麦隐性恢复基因表达载体pGEII-Ms1-ZmAA1-DsRed2,通过农杆菌介导转化拟南芥,并利用红色荧光对转基因拟南芥种子进行验证,说明小麦隐性恢复基因表达载体构建成功。鉴于小麦转化难度较大,尤其是冬小麦基因型的复杂性,所以本研究暂时未能获得转基因小麦材料。下一步的工作重点是建立现有隐性核不育小麦材料(冬小麦)的再生体系,然后将构建好的小麦隐性恢复基因表达载体转化到隐性核不育小麦材料中,从而获得新型的隐性核不育系,为采用该技术体系开展杂交小麦的育种提供理论和材料 依据。
猜你喜欢
新疆农业科学(2022年6期)2022-07-13
中国农学通报(2022年12期)2022-06-01
教育家(2022年17期)2022-04-23
作物学报(2022年4期)2022-02-10
中学生物学(2019年7期)2019-10-17
中学生理科应试(2016年12期)2017-01-07
新高考·高一数学(2016年3期)2016-05-19
飞碟探索(2015年9期)2015-11-05
红领巾·探索(2015年9期)2015-09-10
恋爱婚姻家庭·养生版(2011年8期)2011-05-14
