
小麦面粉色泽相关基因在河南地方品种中的分布
2022-08-09 01:14李春鑫赵明忠韩留鹏李正玲昝香存
麦类作物学报 2022年6期
李春鑫,赵明忠,韩留鹏,高 崇,李正玲,王 艳,昝香存,胡 琳
(1.河南省农业科学院经济作物研究所,河南郑州 450002;2.河南省农业科学院作物分子育种研究院,河南郑州 450002)
小麦农家品种遗传差异大,性状表型变异丰富,是小麦种质资源重要组成部分,在人类社会发展过程中,对全球小麦生产发挥着基础性作用。随着现代育种技术的兴起,地方品种逐渐被现代品种代替,但随着现代品种的增多,生产上种植的品种逐渐出现了遗传基础狭窄、抗性降低、品质类型单一等问题。河南省是我国小麦传统核心种植区,处于南北气候类型过渡地带,在漫长的传统农业生产中,保留了丰富的地方品种,是今后小麦育种和遗传改良的重要基因来源。
随着社会生活水平的提高,人们对小麦食品的要求更加全面,传统小麦制品如馒头、面条的色泽等外观特性作为制成品品质的直观体现,已成为小麦品质育种的一个重要改良目标。小麦制品的色泽相关基因中,PPO活性是影响在存放期间面粉颜色褐化的主要原因[1],而LOX可以提高小麦面粉的白度,对小麦的加工价值有较大影响[2-3]。
在小麦育种中,通过分子标记选择低PPO活性和高LOX活性的育种材料是品质改良的普遍做法[3-6]。小麦的LOX活性是受多基因控制的数量性状,在染色体第四和第五同源群的4A、4B、4D、5A、5B和5D染色体上均发现了LOX QTL[4,7]。Geng等[8]发现,来自染色体4BS的TaLox-B1基因对LOX活性起重要的调控作用,其互补性检测标记LOX16和LOX18,是目前LOX分子辅助育种的主要检测标记,可以准确地完成对LOX基因的检测[4,6,9]。
PPO可以解释面制品褐化变异的50%~70%[10],是一个高遗传力性状,其主效控制基因位于2AL和2DL上,两个位点对表型变异的解释均在25%以上[11]。Sun等[12]开发了共显性STS标记PPO18,能够有效区分2AL上面的PPO-A1a和PPO-A1b;He等[13]开发了两个互补型的STS标记PPO16和PPO29,用于准确检测另一个主要位点基因PPO-D1a和PPO-D1b;He等[13]开发出了另一个用于检测PPO 2AL位点的功能标记PPO33。同时利用以上四个标记,可以实现对低PPO活性亲本和分离单株的筛选。
本研究以河南省不同地市的308份小麦地方品种为材料,利用LOX16、LOX18、PPO16、PPO18、PPO29和PPO33 共6个功能标记对河南省特色地方品种进行LOX及PPO基因检测,以明确其在河南省地方品种中的分布,为小麦品质育种材料选择提供参考。
1 材料与方法
1.1 试验材料
所用河南省小麦地方品种均来自于河南省农业科学院小麦研究所资源研究室种质库,所有品种在试验前均经过连续两年的田间严格去杂和性状一致性鉴定,本研究选取了308份背景纯合、多样性好的地方品种作为供试材料,品种名称和来源详见表1。
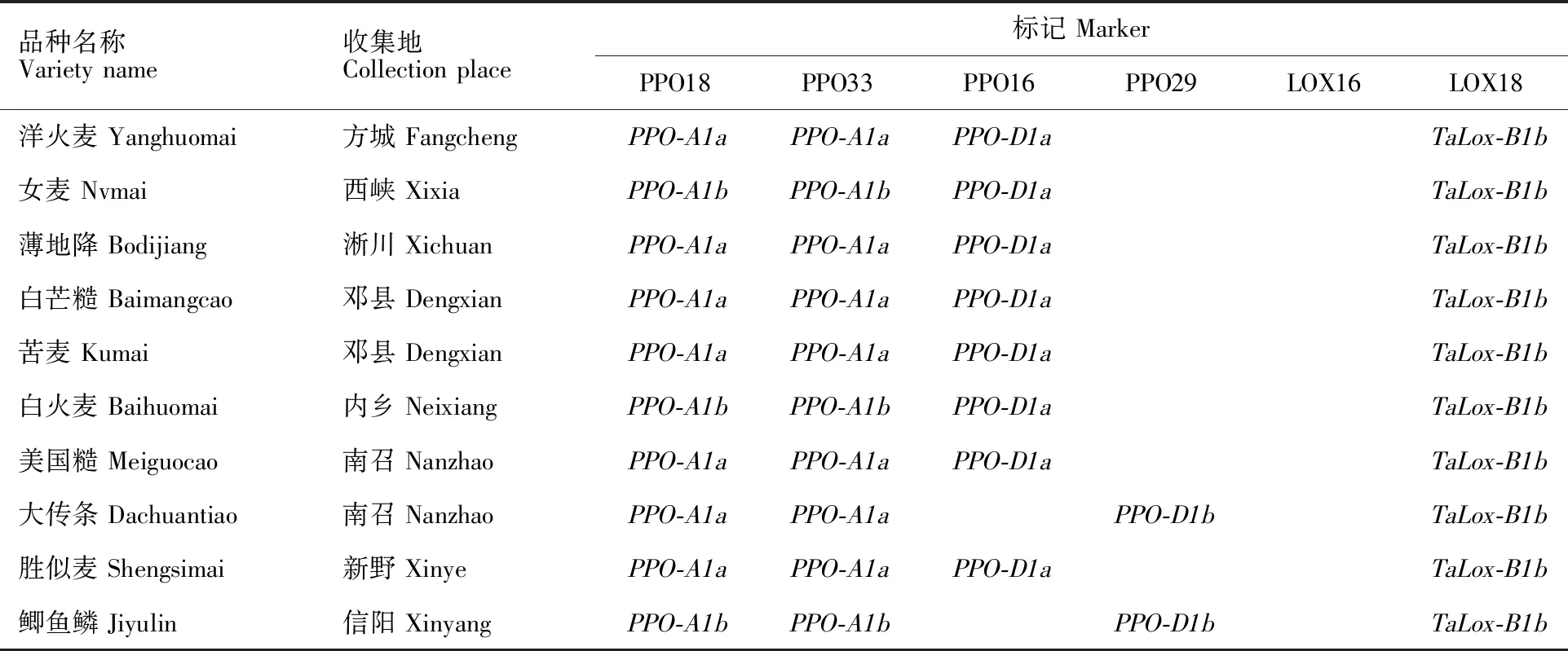
表1 供试小麦品种及其检测基因型Table 1 Wheat varieties and genotypes in the experiment
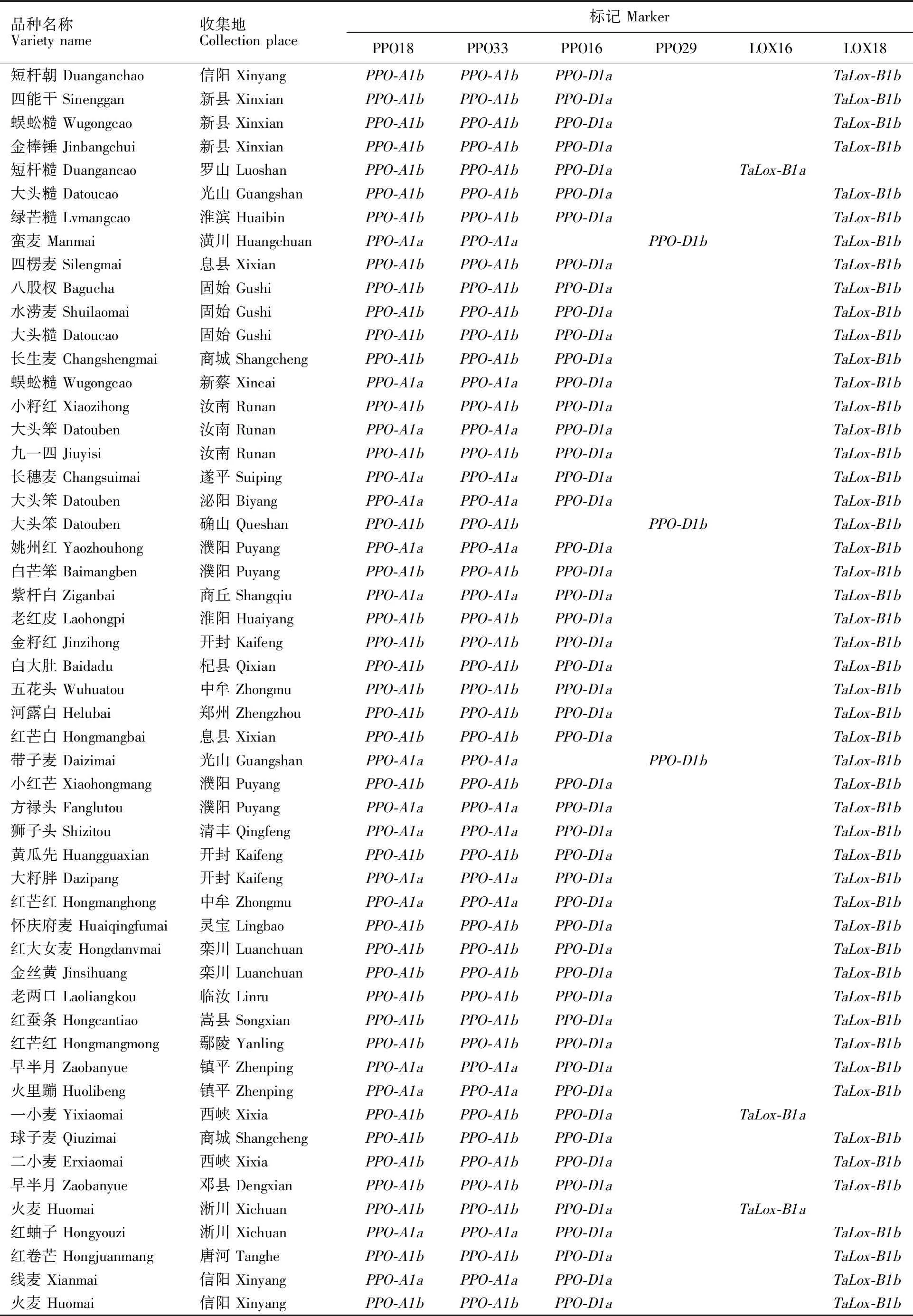
(续表1 Continued table 1)
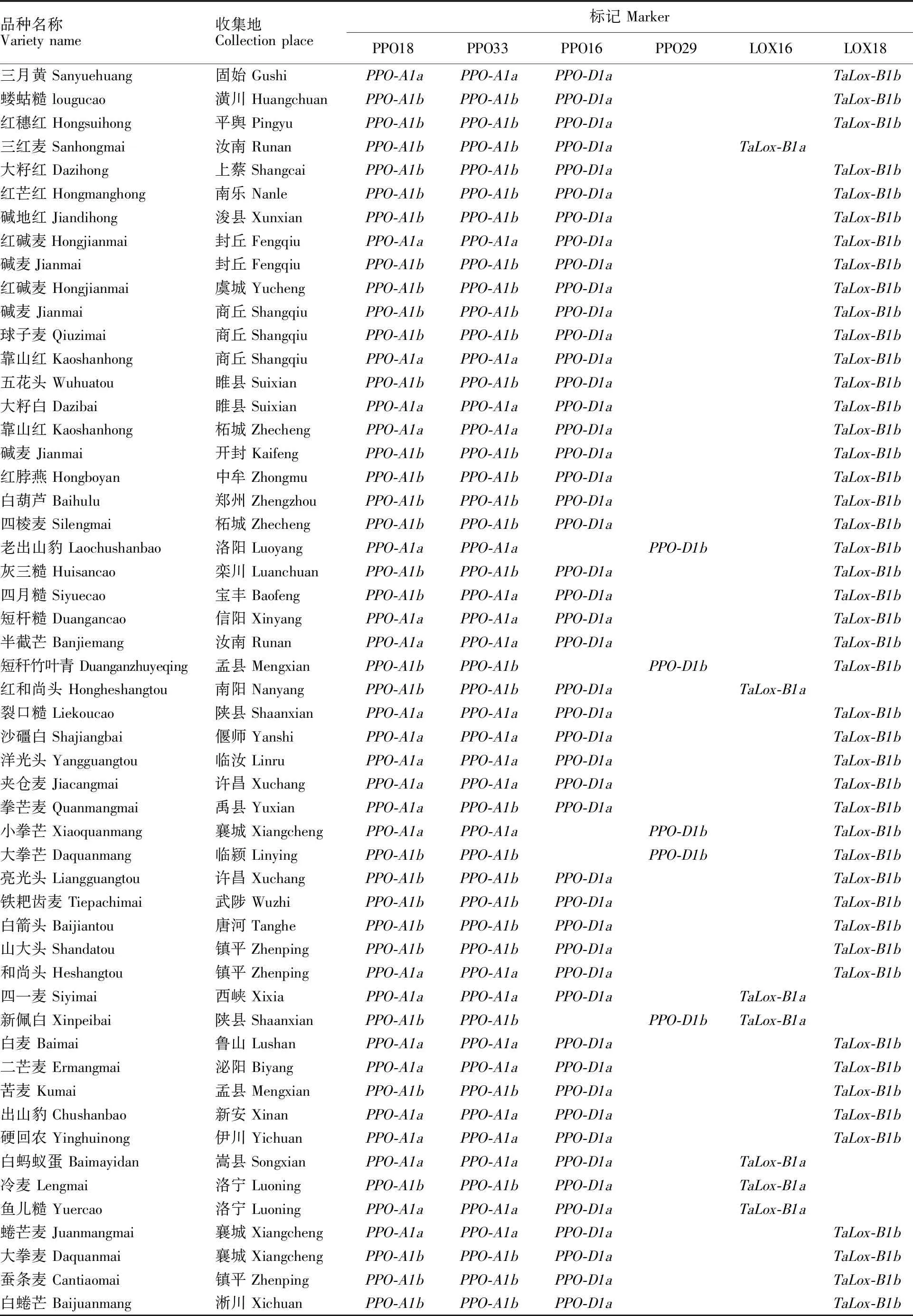
(续表1 Continued table 1)
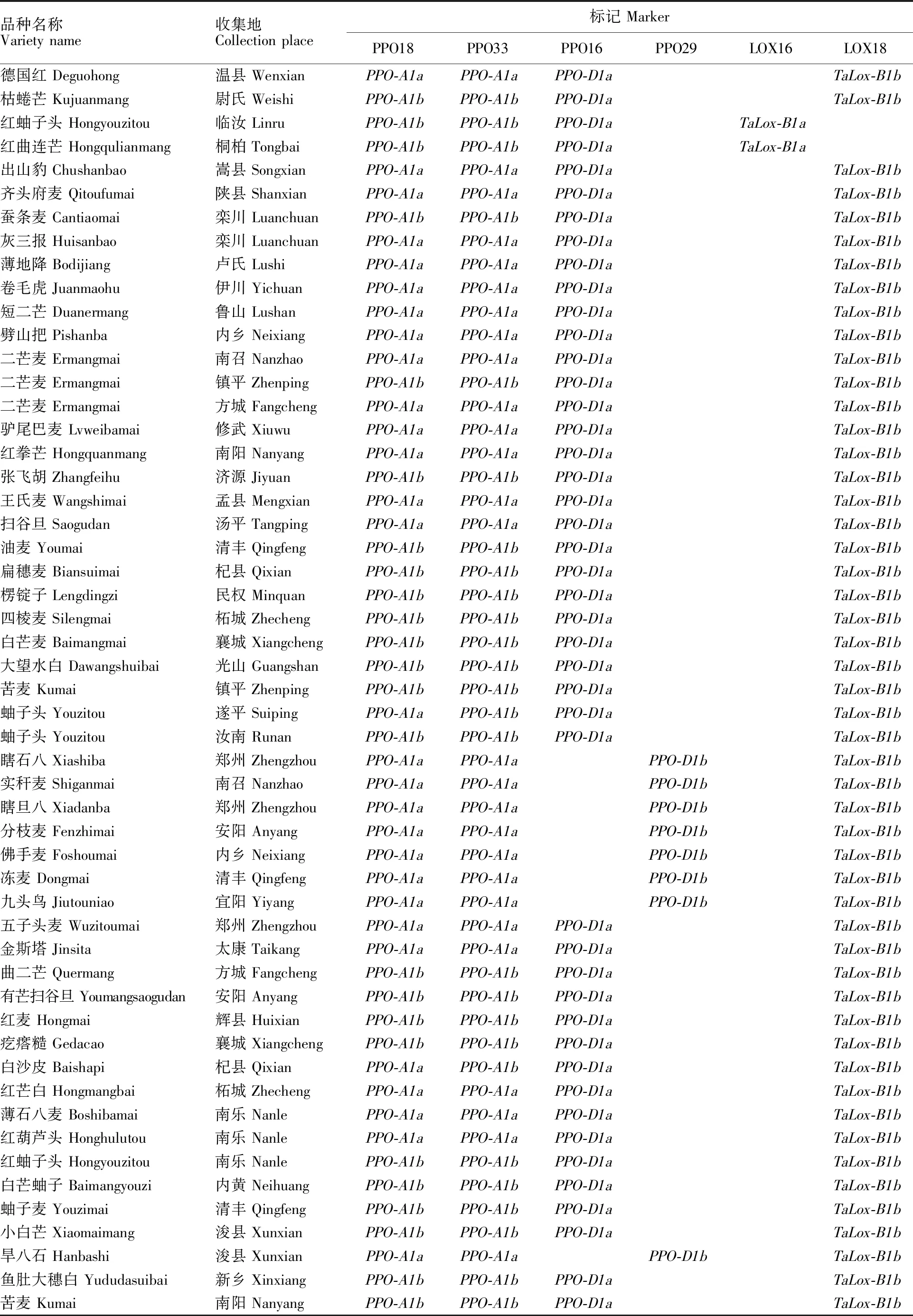
(续表1 Continued table 1)
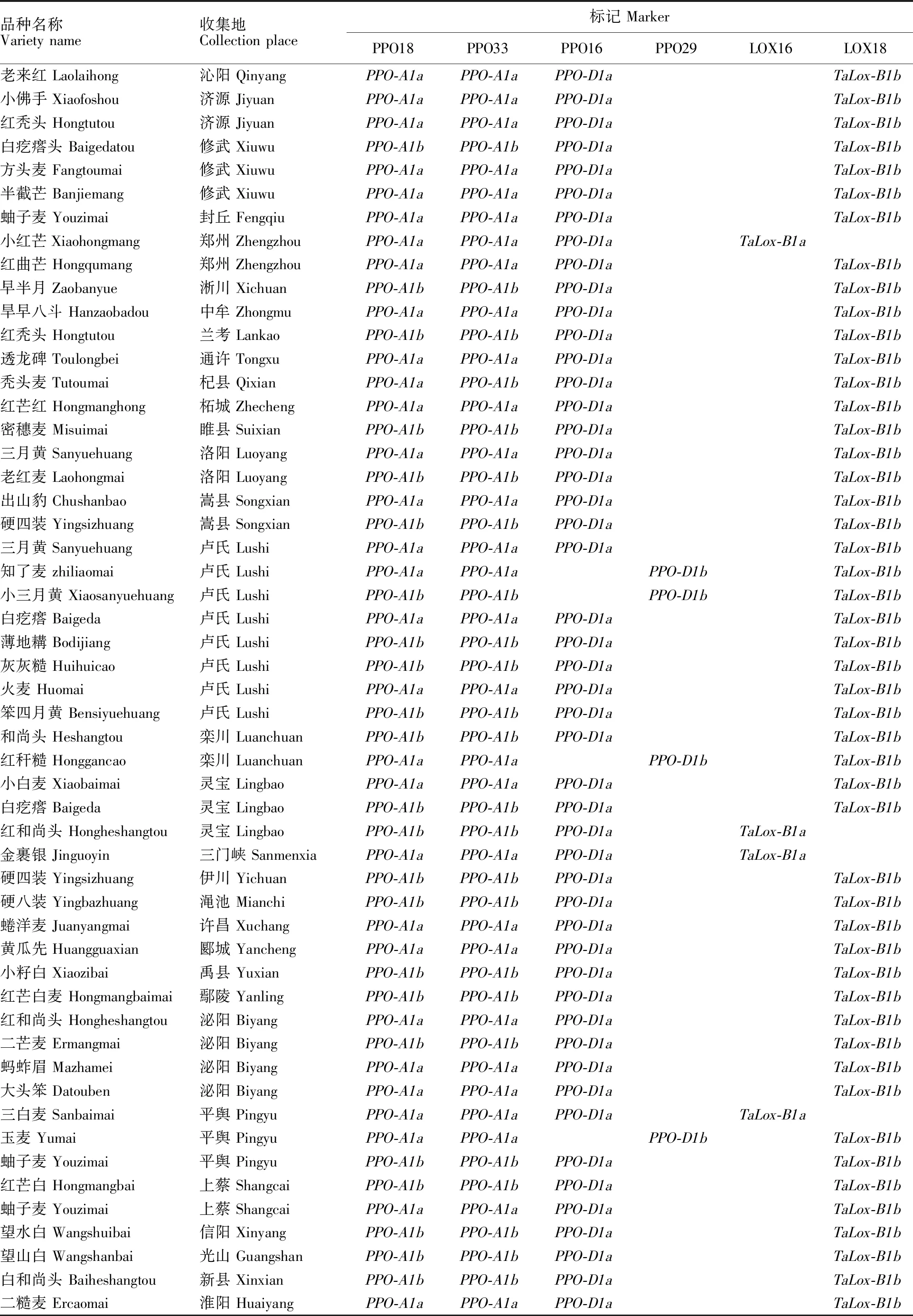
品种名称Varietyname收集地Collectionplace标记MarkerPPO18PPO33PPO16PPO29LOX16LOX18老来红Laolaihong沁阳QinyangPPO-A1aPPO-A1aPPO-D1aTaLox-B1b小佛手Xiaofoshou济源JiyuanPPO-A1aPPO-A1aPPO-D1aTaLox-B1b红秃头Hongtutou济源JiyuanPPO-A1aPPO-A1aPPO-D1aTaLox-B1b白疙瘩头Baigedatou修武XiuwuPPO-A1bPPO-A1bPPO-D1aTaLox-B1b方头麦Fangtoumai修武XiuwuPPO-A1aPPO-A1aPPO-D1aTaLox-B1b半截芒Banjiemang修武XiuwuPPO-A1aPPO-A1aPPO-D1aTaLox-B1b蚰子麦Youzimai封丘FengqiuPPO-A1aPPO-A1aPPO-D1aTaLox-B1b小红芒Xiaohongmang郑州ZhengzhouPPO-A1aPPO-A1aPPO-D1aTaLox-B1a红曲芒Hongqumang郑州ZhengzhouPPO-A1aPPO-A1aPPO-D1aTaLox-B1b早半月Zaobanyue淅川XichuanPPO-A1bPPO-A1bPPO-D1aTaLox-B1b旱早八斗Hanzaobadou中牟ZhongmuPPO-A1aPPO-A1aPPO-D1aTaLox-B1b红秃头Hongtutou兰考LankaoPPO-A1bPPO-A1bPPO-D1aTaLox-B1b透龙碑Toulongbei通许TongxuPPO-A1aPPO-A1aPPO-D1aTaLox-B1b秃头麦Tutoumai杞县QixianPPO-A1aPPO-A1bPPO-D1aTaLox-B1b红芒红Hongmanghong柘城ZhechengPPO-A1aPPO-A1aPPO-D1aTaLox-B1b密穗麦Misuimai睢县SuixianPPO-A1bPPO-A1bPPO-D1aTaLox-B1b三月黄Sanyuehuang洛阳LuoyangPPO-A1aPPO-A1aPPO-D1aTaLox-B1b老红麦Laohongmai洛阳LuoyangPPO-A1bPPO-A1bPPO-D1aTaLox-B1b出山豹Chushanbao嵩县SongxianPPO-A1aPPO-A1aPPO-D1aTaLox-B1b硬四装Yingsizhuang嵩县SongxianPPO-A1bPPO-A1bPPO-D1aTaLox-B1b三月黄Sanyuehuang卢氏LushiPPO-A1aPPO-A1aPPO-D1aTaLox-B1b知了麦zhiliaomai卢氏LushiPPO-A1aPPO-A1aPPO-D1bTaLox-B1b小三月黄Xiaosanyuehuang卢氏LushiPPO-A1bPPO-A1bPPO-D1bTaLox-B1b白疙瘩Baigeda卢氏LushiPPO-A1aPPO-A1aPPO-D1aTaLox-B1b薄地耩Bodijiang卢氏LushiPPO-A1bPPO-A1bPPO-D1aTaLox-B1b灰灰糙Huihuicao卢氏LushiPPO-A1bPPO-A1bPPO-D1aTaLox-B1b火麦Huomai卢氏LushiPPO-A1aPPO-A1aPPO-D1aTaLox-B1b笨四月黄Bensiyuehuang卢氏LushiPPO-A1bPPO-A1bPPO-D1aTaLox-B1b和尚头Heshangtou栾川LuanchuanPPO-A1bPPO-A1bPPO-D1aTaLox-B1b红秆糙Honggancao栾川LuanchuanPPO-A1aPPO-A1aPPO-D1bTaLox-B1b小白麦Xiaobaimai灵宝LingbaoPPO-A1aPPO-A1aPPO-D1aTaLox-B1b白疙瘩Baigeda灵宝LingbaoPPO-A1bPPO-A1bPPO-D1aTaLox-B1b红和尚头Hongheshangtou灵宝LingbaoPPO-A1bPPO-A1bPPO-D1aTaLox-B1a金裹银Jinguoyin三门峡SanmenxiaPPO-A1aPPO-A1aPPO-D1aTaLox-B1a硬四装Yingsizhuang伊川YichuanPPO-A1bPPO-A1bPPO-D1aTaLox-B1b硬八装Yingbazhuang渑池MianchiPPO-A1bPPO-A1bPPO-D1aTaLox-B1b蜷洋麦Juanyangmai许昌XuchangPPO-A1aPPO-A1aPPO-D1aTaLox-B1b黄瓜先Huangguaxian郾城YanchengPPO-A1aPPO-A1aPPO-D1aTaLox-B1b小籽白Xiaozibai禹县YuxianPPO-A1bPPO-A1bPPO-D1aTaLox-B1b红芒白麦Hongmangbaimai鄢陵YanlingPPO-A1bPPO-A1bPPO-D1aTaLox-B1b红和尚头Hongheshangtou泌阳BiyangPPO-A1aPPO-A1aPPO-D1aTaLox-B1b二芒麦Ermangmai泌阳BiyangPPO-A1bPPO-A1bPPO-D1aTaLox-B1b蚂蚱眉Mazhamei泌阳BiyangPPO-A1aPPO-A1aPPO-D1aTaLox-B1b大头笨Datouben泌阳BiyangPPO-A1aPPO-A1aPPO-D1aTaLox-B1b三白麦Sanbaimai平舆PingyuPPO-A1aPPO-A1aPPO-D1aTaLox-B1a玉麦Yumai平舆PingyuPPO-A1aPPO-A1aPPO-D1bTaLox-B1b蚰子麦Youzimai平舆PingyuPPO-A1bPPO-A1bPPO-D1aTaLox-B1b红芒白Hongmangbai上蔡ShangcaiPPO-A1bPPO-A1bPPO-D1aTaLox-B1b蚰子麦Youzimai上蔡ShangcaiPPO-A1aPPO-A1aPPO-D1aTaLox-B1b望水白Wangshuibai信阳XinyangPPO-A1bPPO-A1bPPO-D1aTaLox-B1b望山白Wangshanbai光山GuangshanPPO-A1bPPO-A1bPPO-D1aTaLox-B1b白和尚头Baiheshangtou新县XinxianPPO-A1bPPO-A1bPPO-D1aTaLox-B1b二糙麦Ercaomai淮阳HuaiyangPPO-A1aPPO-A1aPPO-D1aTaLox-B1b
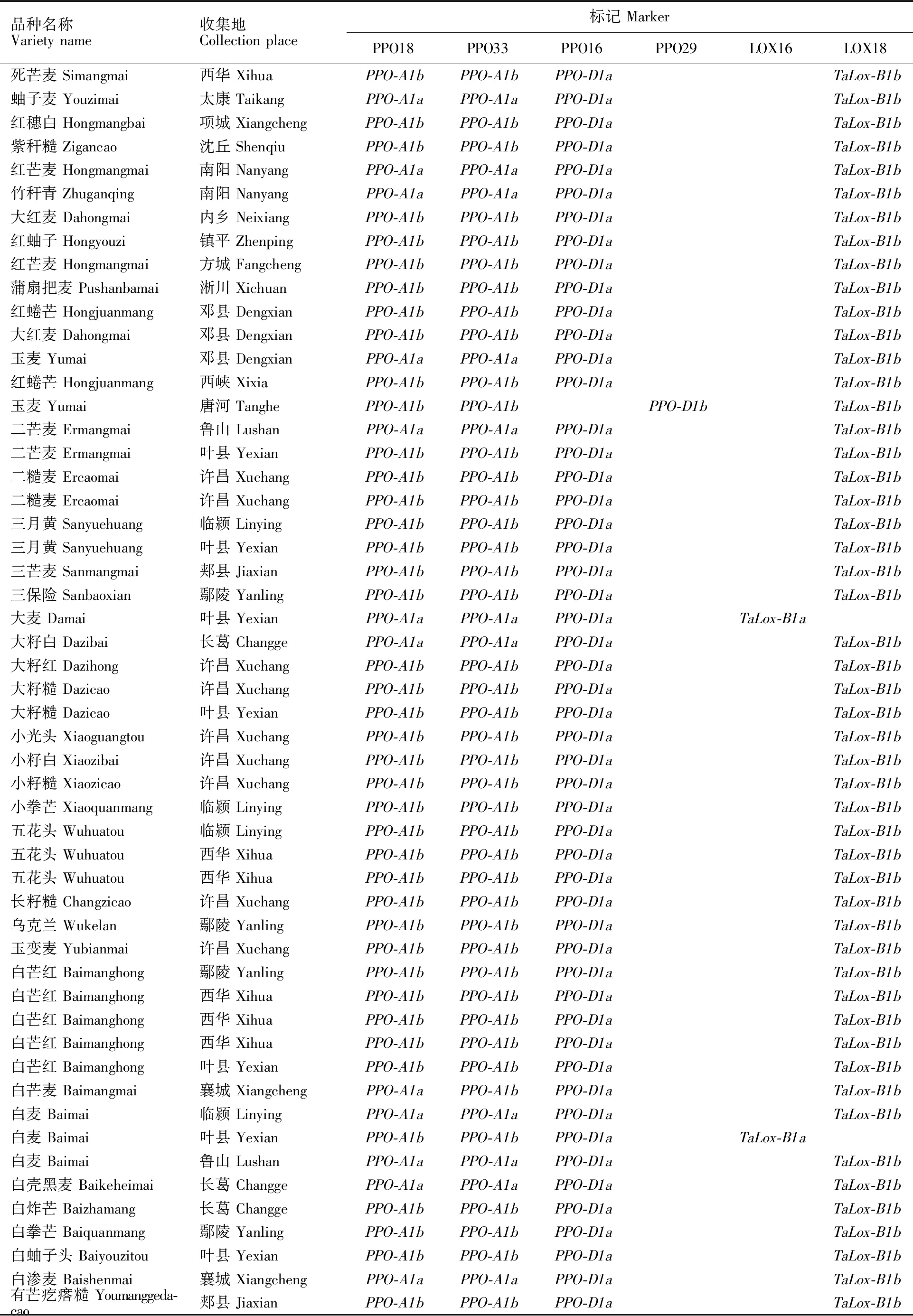
(续表1 Continued table 1)
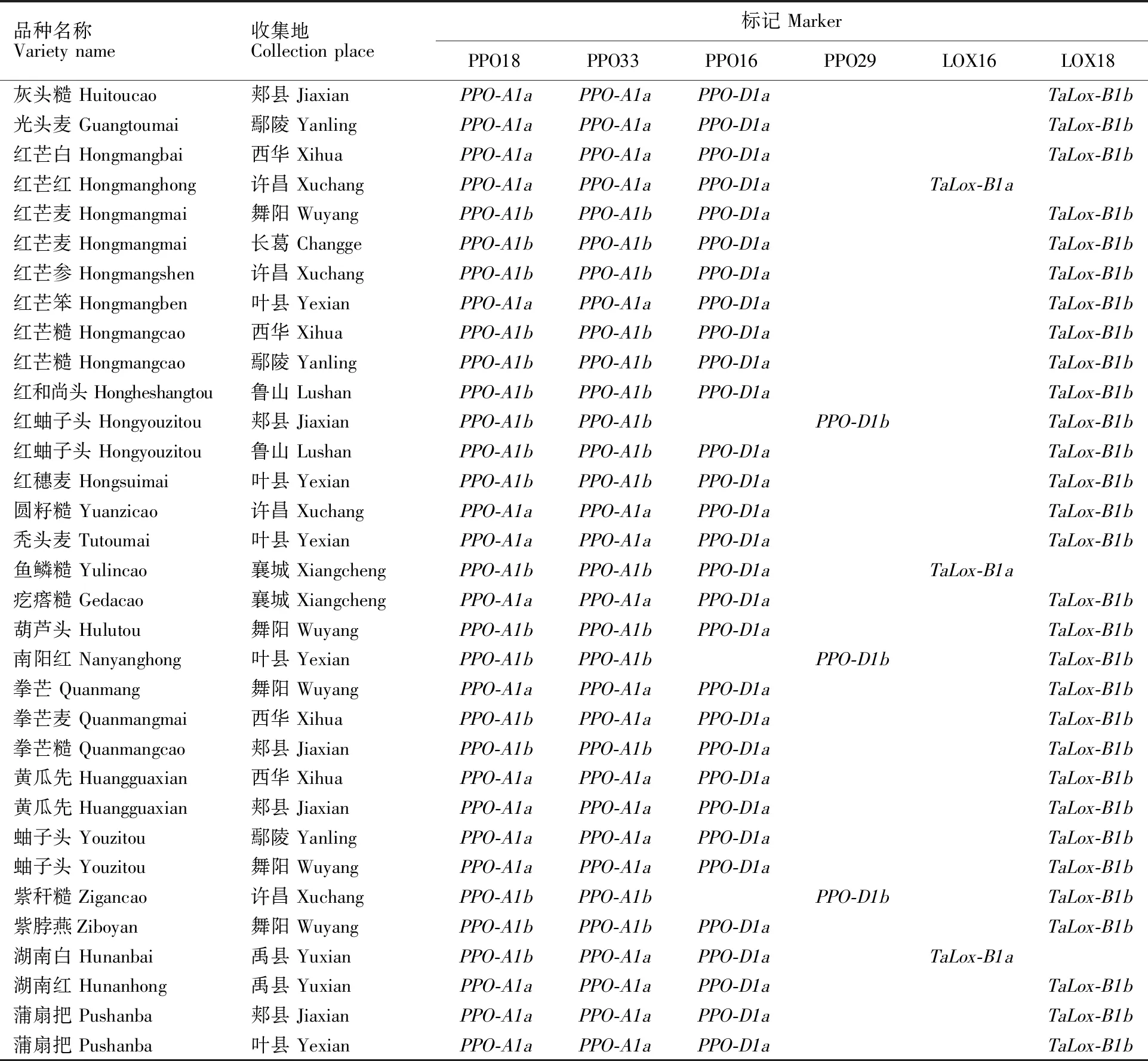
(续表1 Continued table 1)
1.2 试验方法
1.2.1 基因组DNA的提取
每份材料挑选饱满种子5粒,放置于玻璃培养皿中20 ℃发苗培养,7~10 d后,剪取生长健康叶片参照Lagudah等方法[14]提取DNA,用1%琼脂糖检测DNA提取质量,所有样品DNA均稀释为100 ng·μL-1备用。
1.2.2 面粉色泽相关基因检测
试验用PPO-A1的功能标记PPO18、PPO33和PPO-D1位点的互补型功能标记PPO16、PPO29检测品种间多酚氧化酶基因;用TaLox-B1的两个互补型功能标记LOX16和LOX18检测脂肪氧化酶基因,所有引物均由生工生物工程(上海)股份有限公司合成,具体序列见表2。
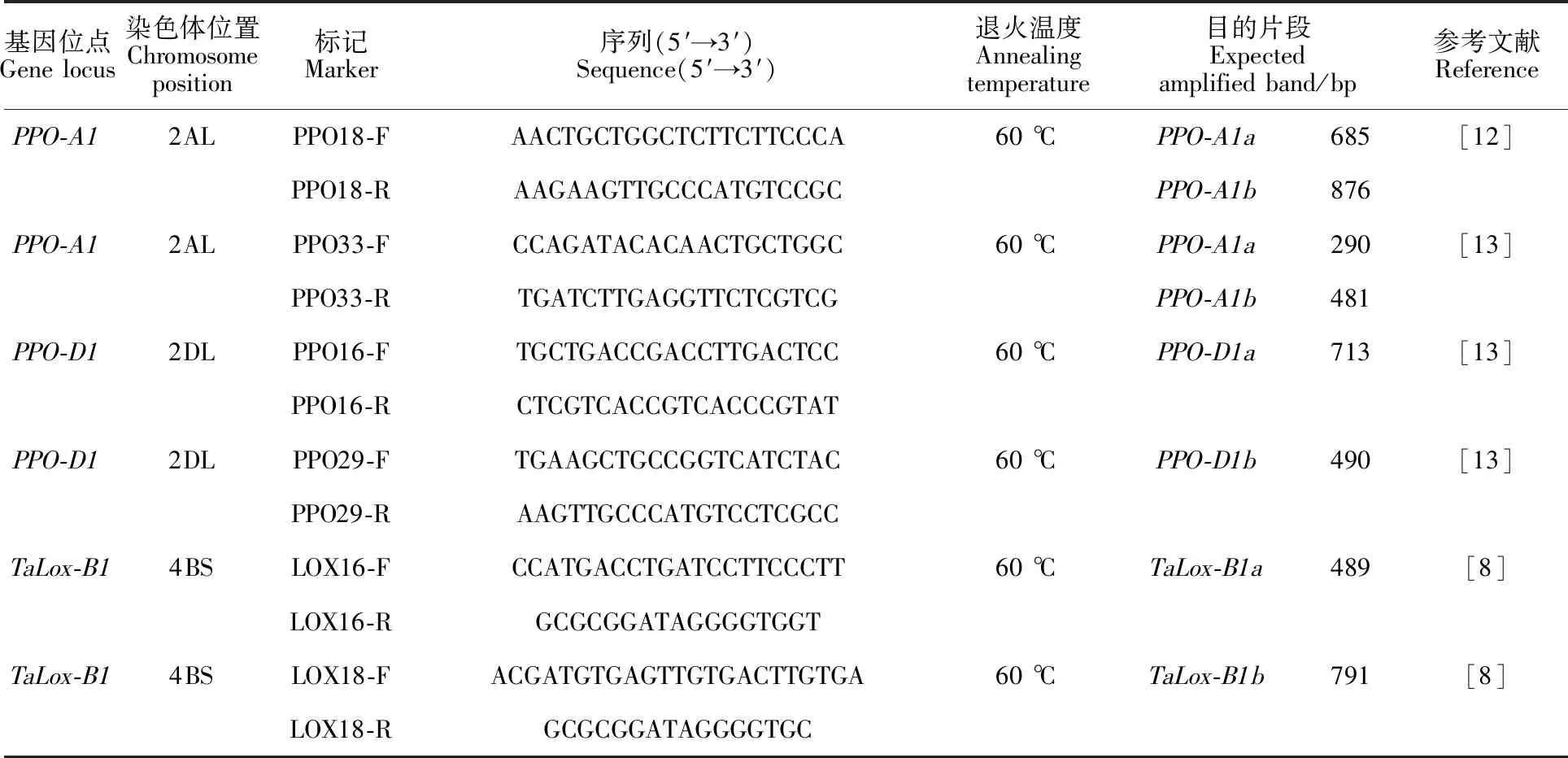
表2 基因检测所用分子标记引物序列、退火温度、预期扩增条带大小及相应等位基因Table 2 Primer sequence,annealing temperature,expected amplified band size and corresponding allele of the molecular markers used in the genes detection
PCR扩增在Mastercycler Pro S(Eppendorf AG,Hamburg,Germany)上进行,PCR反应体系为10 μL,包括:2 μL DNA模板,1.0 μL 10 × buffer(Mg2+),0.2 μL dNTPs(10 pmol·μL-1),上、下游引物各0.2 μL(10 pmol·μL-1) ,0.06 UTaqDNA polymerase(5 U·μL-1),6.34 μL 无菌水。PCR扩增程序为:95 ℃变性 5 min;94 ℃变性30 s,60 ℃退火30~50 s,72 ℃延伸 45~60 s,36个循环;72 ℃ 10 min。PCR扩增产物利用1%琼脂糖凝胶电泳检测。用EXCEL对电泳结果进行统计分析。
2 结果与分析
2.1 多酚氧化酶基因检测结果
PPO18是共显性功能标记,可以精确检测高多酚氧化酶活性基因PPO-A1a和低多酚氧化酶基因PPO-A1b[15],其扩增片段分别为685 bp和876 bp(图1)。在308份地方品种中,检测到携带PPO-A1a基因型的共有133个品种,频率为 43.18%,携带PPO-A1b基因型的品种共175个,频率为56.82%(表1);同一个位点的另一个标记PPO33扩增结果(图2)显示,携带PPO-A1a基因的共有131个品种,出现频率为 42.53%,携带PPO-A1b基因的品种共177个,频率为57.47%。两个标记扩增结果比较显示,308个不同地方品种中,两个品种(拳芒麦和白芒麦)扩增结果不一致,标记PPO18对两个品种的扩增结果为PPO-A1a、PPO-A1b,而标记PPO33对两个品种的扩增结果为PPO-A1b、PPO-A1a,两个标记扩增结果符合度为99.35%。

M:DNA ladder 2000;1~12:品种。下同。
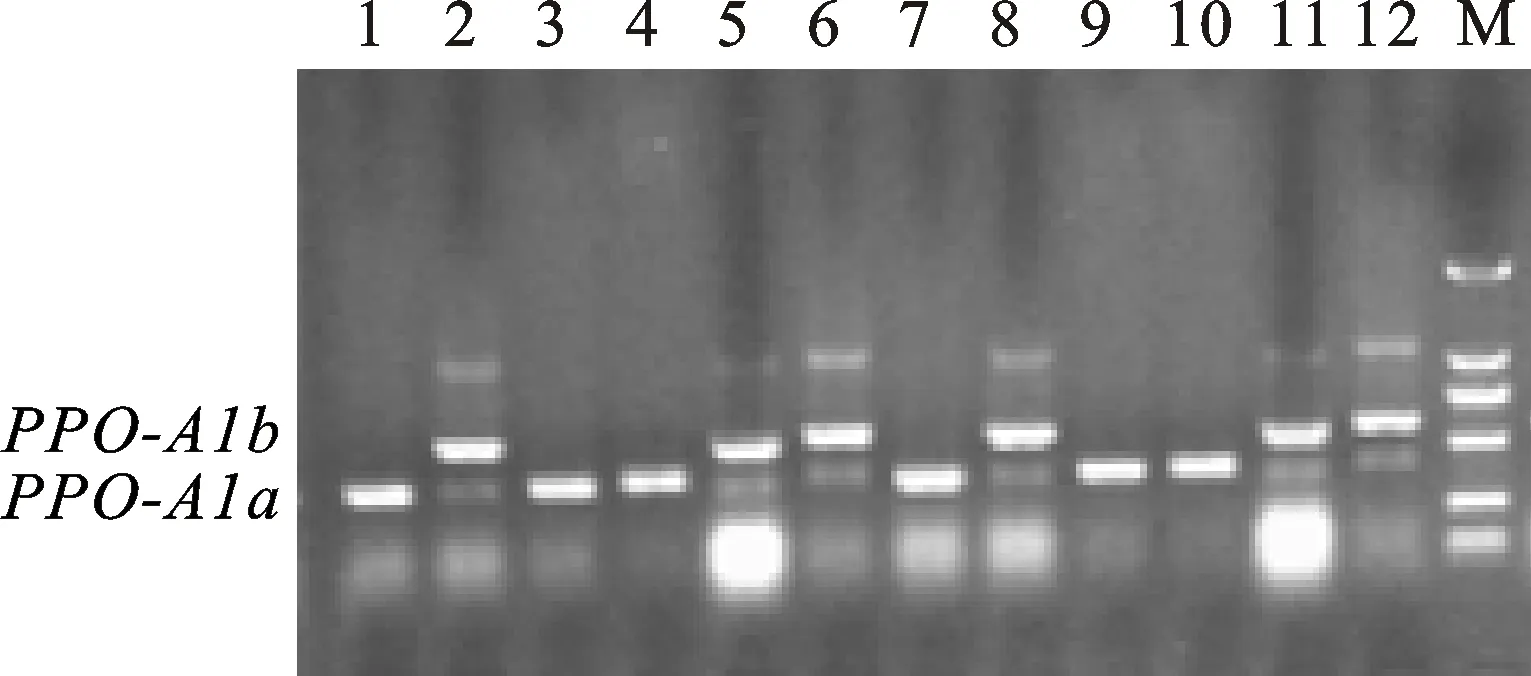
图2 标记PPO33对部分品种的扩增结果Fig.2 PCR amplification from partial landraces with marker PPO33
多酚氧化酶基因位点PPO-D1的两个标记PPO16和PPO29也是互补型标记,分别用于检测等位基因PPO-D1a(中等低多酚氧化酶活性)和PPO-D1b(中等高多酚氧化酶活性)[15],其相应扩增片段为713 bp(图3)和490 bp(图4)。利用标记PPO16对308个地方品种扩增,检测出目标基因282个,频率为91.56%;利用PPO29检测到含有目的基因的品种数为26个,频率为 8.44%,两个标记扩增结果完全吻合。
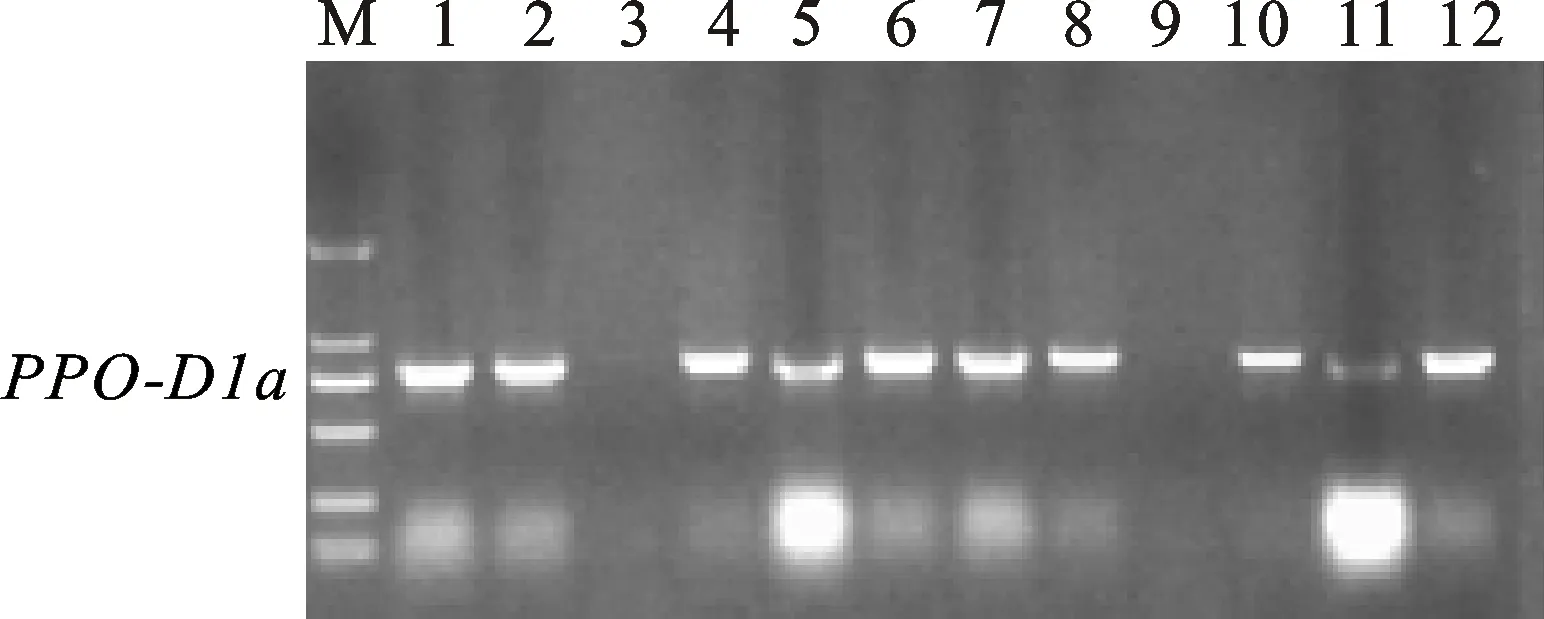
图3 标记PPO16对部分品种的扩增结果Fig.3 PCR amplification from partial landraces with marker PPO16
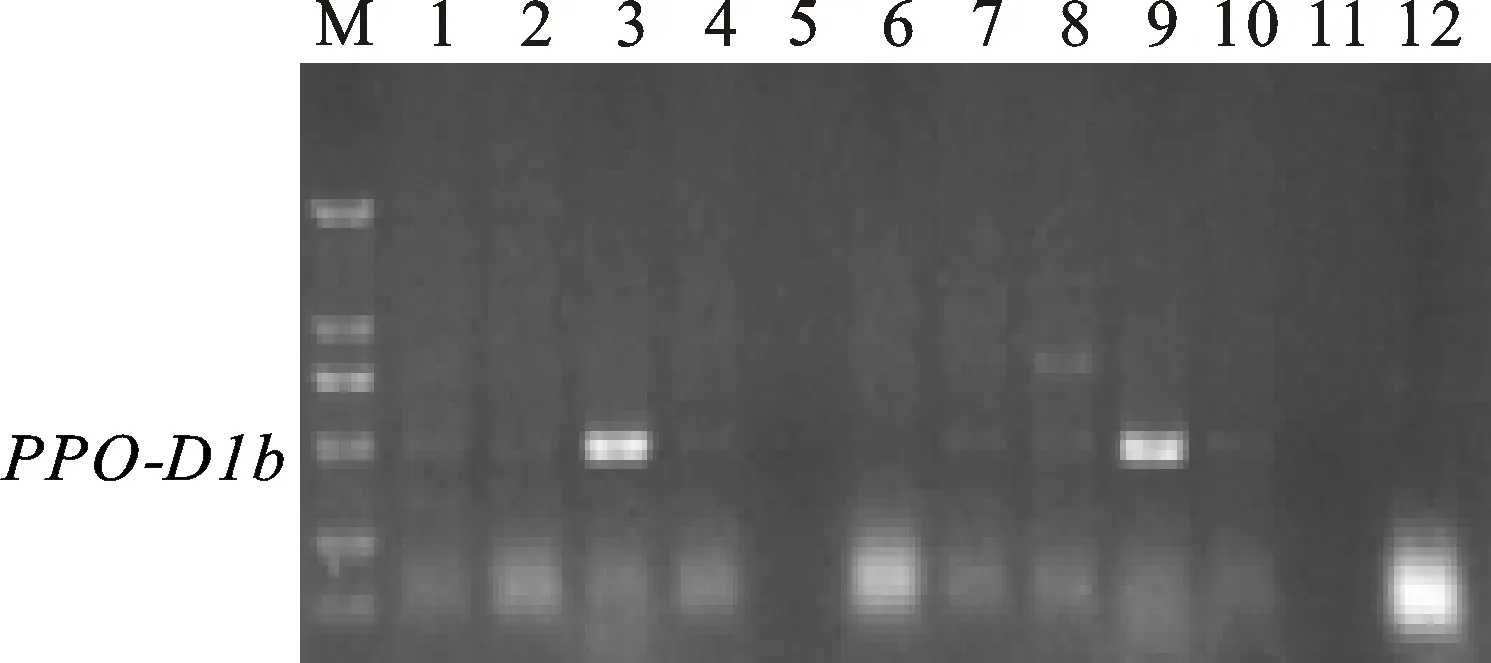
图4 标记PPO29对部分品种的扩增结果Fig.4 PCR amplification from partial landraces with marker PPO29
2.2 脂肪氧化酶基因检测结果
两个互补型分子标记LOX16和LOX18所检测的目标是脂肪氧化酶基因TaLox-B1a和TaLox-B1b。其中,TaLox-B1a的扩增片段为489 bp(图5),是高活性脂肪氧化酶,TaLox-B1b的扩增片段为791 bp(图6),是低活性脂肪氧化酶基因[9]。在308个品种中,携带TaLox-B1b和TaLox-B1a的品种数量分别为287和21,其频率分别为93.18%和6.82%。
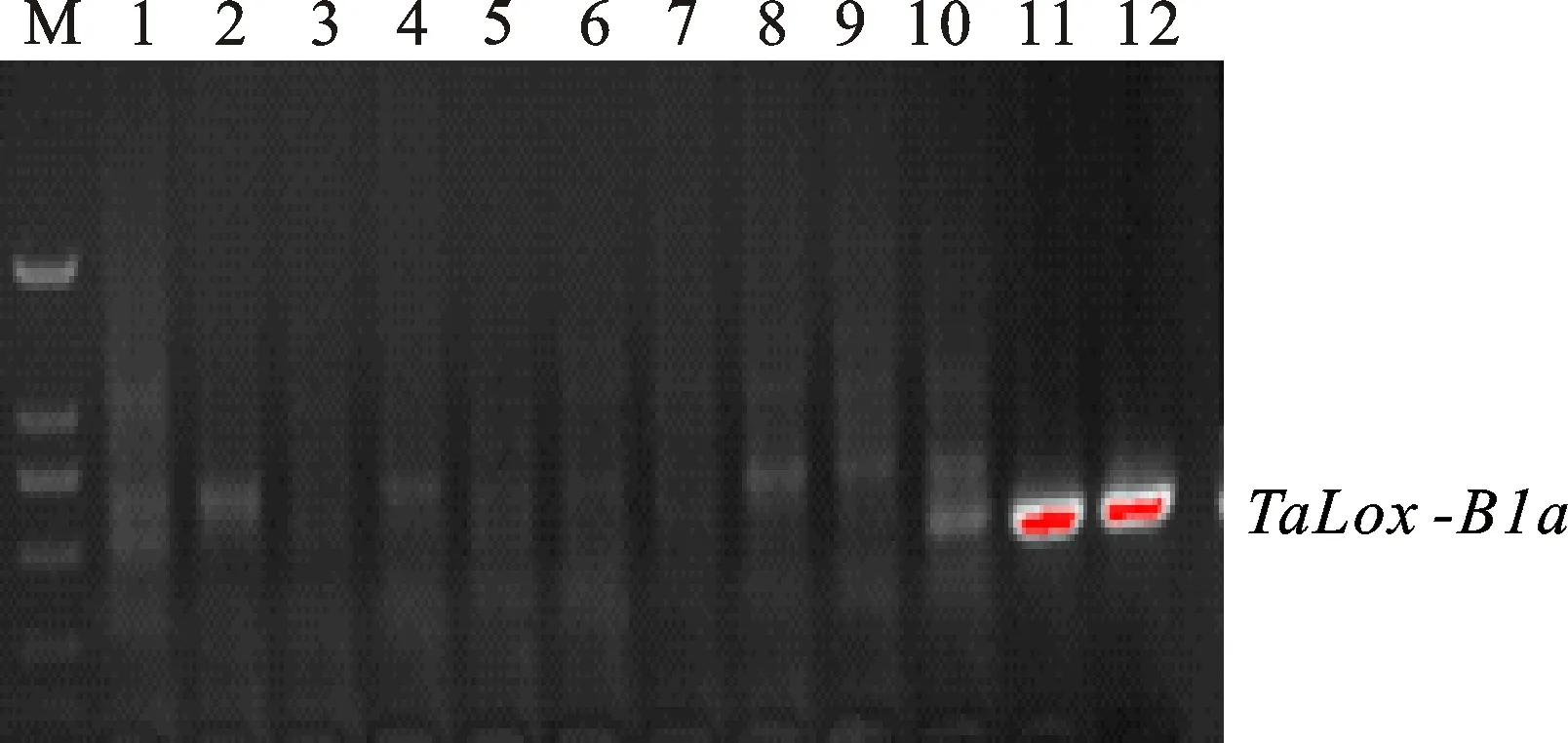
图5 标记LOX16对部分品种的扩增结果Fig.5 PCR amplification from partial landraces with marker LOX16

图6 标记LOX18对部分品种的扩增结果Fig.6 PCR amplification from partial landraces with marker LOX18
2.3 小麦面粉色泽性状优异基因组合分析
在306个供试品种中(不包括拳芒麦和白芒麦),低活性多酚氧化酶基因型组合(PPO-A1b/PPO-D1a)、较低活性多酚氧化酶基因型组合 (PPO-A1a/PPO-D1a)、较高活性多酚氧化酶基因型组合(PPO-A1b/PPO-D1b)和高活性多酚氧化酶基因型组合(PPO-A1a/PPO-D1b)在306个地方品种中品种数分别为166、10、114和16。
低活性多酚氧化酶和高活性脂肪氧化酶可增加面粉及其制品的色泽品质,不同活性多酚氧化酶基因和脂肪氧化酶基因组合代表了不同的面粉色泽性状表现,表3是根据分子标记检测结果所统计的8种不同基因型组合及其携带品种的数量。因为尚无多酚氧化酶和脂肪氧化酶基因各自对面粉色泽表型解释值比较的报道,因此,无法对8种不同基因型组合所代表的色泽性状表型进行精确排序,推测最优组合为PPO-A1b/PPO-D1a/TaLox-B1a,最差组合为PPO-A1a/PPO-D1b/TaLox-B1b,在306个品种中,两种基因型组合数量为11和16。

表3 PPO和LOX两个性状不同基因型组合及其品种数量Table 3 The variety number with different genotypecombinationfor PPO and LOX
3 讨 论
小麦多酚氧化酶和脂肪氧化酶作为影响小麦面粉及其制品色泽的重要性状,相关基因的分子标记在现代品种中的应用报到较多[3-6,9],但是对河南省地方品种的检测尚未见报道。曹 东等[4]对甘肃近10年的审定品种进行了PPO和LOX基因检测,发现TaLox-B1b的频率是TaLox-B1a的3倍以上,4个PPO等位基因频率基本接近;这与吴培培等[6]对黄淮地区306份品种的LOX基因检测结果基本一致;陈 泠等[17]在对全国277份育成品种的PPO等位基因检测后,发现PPO-A1b在河南品种中频率只有28.6%,亟需在后续品种中加强PPO-A1b利用。本研究首次对作为黄淮小麦亲本重要来源的地方品种的相关基因进行了检测,结果显示,携带PPO-A1b的品种数量比PPO-A1a的品种高13.72%(拳芒麦和白芒麦在等位基因PPO-A1a、PPO-A1b统计时去除),与前人报道的黄淮地区现代品种中的PPO-A1b比例偏低相比改善明显,而PPO-D1a、PPO-D1b在品种中分布极不均匀,低多酚氧化酶活性基因PPO-D1a占支配地位,出现频率是PPO-D1b的10.8倍,因此,低活性多酚氧化酶基因型组合 (PPO-A1b/PPO-D1a)在306份品种中有166个,活性占比为54.25%,表明河南小麦地方品种的低活性多酚氧化酶基因分布整体表现较为 理想。
有研究认为,PPO-A1的效应显著大于PPO-D1[11,16],较低活性多酚氧化酶基因型组合 (PPO-A1b/PPO-D1b)、较高活性多酚氧化酶基因型组合(PPO-A1a/PPO-D1a)和高活性多酚氧化酶基因型组合(PPO-A1a/PPO-D1b)在306个地方品种中分别有114、10和16个,频率分别为37.25%、3.27%和5.23%,高和较高活性多酚氧化酶基因型组合仅为8.50%,说明河南省地方品种的多酚氧化酶性状是以中、低活性基因型为 主体。
脂肪氧化酶基因主要影响面粉贮藏期间的白度[9],是小麦育种一个重要品质指标,LOX16和LOX18是LOX位点主要的检测标记,本研究检测结果显示,携带低活性脂肪氧化酶基因TaLox-B1b的品种数量是高活性脂肪氧化酶基因TaLox-B1a品种的13.67倍,即河南省地方品种中面粉白度优异基因型频率较低,需要在亲本组合选配时加入高活性脂肪氧化酶基因品种。综合考虑脂肪氧化酶和多酚氧化酶基因,优异基因型组合为PPO-A1b/PPO-D1a/TaLox-B1a,在306个品种中,只有11个品种具有这种基因型组合,其频率为3.59%,而最差组合为PPO-A1a/PPO-D1b/TaLox-B1b,共有16个品种携带此基因型组合,这表明河南省的地方品种中面粉色泽基因型组合整体呈现类正态分布的状态,最优和最差基因型组合的品种数量少,而中间类型品种占主体地位(表3)。
本试验中,除标记PPO18和PPO33在品种拳芒麦和白芒麦中不一致外,其他结果均相互吻合。对于标记PPO18和PPO33在个别品种中的检测差异,我们推断可能是两个地方品种在长期自交过程中,在PPO-A1位点出现了自然变异,在基因扩增区段发生序列/结构的改变,从而导致了检测结果的差异,更直接的证据需要通过进一步的试验进行验证。
猜你喜欢
传染病信息(2022年4期)2022-11-23
农产品加工(2022年15期)2022-09-26
安徽农学通报(2022年6期)2022-04-07
昆明医科大学学报(2022年1期)2022-02-28
载人航天(2021年5期)2021-11-20
新课程·下旬(2018年8期)2018-11-10
热带作物学报(2018年5期)2018-05-14
农业研究与应用(2017年2期)2017-07-01
中学生理科应试(2016年4期)2016-11-19
江苏农业科学(2015年2期)2015-03-12
