
基于传递熵的复杂网络人体平衡特征分析
2022-08-03 07:39王哲远罗志增裘晟晨
中国生物医学工程学报 2022年2期
王哲远 罗志增 裘晟晨
1(杭州电子科技大学圣光机联合学院,杭州 310018)
2(杭州电子科技大学自动化学院智能控制与机器人研究所,杭州 310018)
3(浙江大学医学院附属第四医院,浙江 金华 322000)
引言
通过医学手段来确定被试者平衡能力等级的方法被广泛应用于运动康复、航空航天等领域,其目的是为患者制定合理的健康锻炼和康复方案。但目前主流医学评估方法多通过人为观测的量表法等来完成平衡等级的判断,带有较强的主观性[1-2]。 人体平衡能力是一种维持身体静止或运动状态的协调能力,由视觉、本体觉、前庭觉等组成的一套内源调节系统承担这一功能[3-5]。 其关键部分是通过大脑整合上述各系统中接受的外界环境、内部身体信息,并利用感觉运动相关脑区联合控制肢体的骨骼肌系统,通过不断改变人体姿势从而保持身体处于协调状态。
近来以脑电信号(electroencephalogram,EEG)以识别大脑状态及部分神经疾病成为一种较为流行的趋势[6]。 Puntkattalee 等[7]研究发现,静态站立平衡的脑电也会由于人体不同的平衡能力而显示出不同的特征,这为人体平衡估计提供了新的思路;Masakado 等[8]采用大脑皮质-肌肉相干性方法(corticalmuscular coherence, CMC)分析人体平衡但其方法局限于单个节点之间的耦合关系,缺乏如脑功能网络的整体性分析结论;金晟等[9]提出基于脑肌电相干性特征表征人体平衡等能力,但脑肌耦合中所涉及的频带转换会导致数据断层。 本研究采用脑-脑耦合的方式,力求避免上述问题。
人体的平衡维持往往依靠几次主导的平衡调节实现,每次调节指令的产生到下达再到完成调节动作一般持续0.5~2 s[10],而整个调节过程则会涉及到各脑区的协同机制。 神经生理学研究表明,信息在神经元间以离子交换形式来传递,这种交换伴随电位变化[11]。 单个神经元产生的电位变化较微弱,但多个神经元之间存在同步性时会使得电位相互叠加,呈现整体的外部效应,最终呈现形式是整体区域之间产生有规律的电位变化,因此相位同步是脑区间进行通信的重要机制,这种群体的信息传递同时也被头皮EEG所反映。 Roelfsema 等[12]对于运动区、视觉区的局部场电位记录得出结论:有效的信息传递行为须保持节点间高效且稳定的相位同步状态,要求信息传输时段EEG 信号间的同步锁相值强度至少超过平均静息态锁相位值(phase locking value, PLV)的24%,且持续时间大于46 ms。
根据上述大脑平衡生理机制,本研究提出一种基于脑区之间相位同步的数据筛选方法,从相位同步角度筛选出人体主导平衡调节数据。 同时为进一步获取筛选后数据的内在联系,搭建基于传递熵特征的平衡脑功能网络,试图从复杂网络角度寻找深层逻辑联系。 传递熵值作为一种可以衡量网络因果性的指标在生物医学中已经得到了较为广泛的应用,其本身对源间的信息量传递有较高的敏感度,而对于传递形式不作要求,因此对于大脑而言其具有较好的信息流通体现性[13]。 一方面,相位同步的数据筛选本身就具有内在的因果关系;另一方面,相较于其他序列因果性判断指标而言,传递熵需要相对较长的时间序列,目前在生物医疗领域也只有脑电信号与心电信号等能满足这一要求,本研究选用传递熵特征构造平衡脑功能网络。
1 材料与方法
本研究以EEG 为研究对象,首先从被试者处采集不同平衡范式下的EEG 信号,经过信号预处理和相位同步处理后完成数据筛选,再由传递熵特征构建不同范式下的网络连接图,通过网络连接图提取新型特征与全局聚类系数,最后将两者组合后输入支持向量机(support vector machine,SVM)中进行分类,整体流程见图1。

图1 利用EEG 信号进行平衡能力评估的流程Fig.1 Organization of the proposed method to assess balance
1.1 实验对象
数据来自20 名被试者,年龄为(30±10)岁。 被试者要求为:在一年内未受过下肢关节损伤,无前庭、视觉疾病史,无酗酒史等容易对人体平衡产生直接影响的疾病和行为,认为每个个体的平衡水平只与年龄有关。 共设计了4 组实验范式,范式编号为Q1 ~ Q4,平衡条件逐渐苛刻,具体要求如表1 所示。 实验前,由被试佩戴博瑞康公司生产的脑电帽,对电极注射PARKER 公司的导电凝胶,使阻抗值达到10 kΩ 以下。 EEG 信号采集通道分别被标记为O1、O2;P7、P3、Pz、P4、P8;T7、C3、Cz、C4、T8;F7、F3、Fz、F4、F8;FP1、FP2 共19 个通道。 在采集信号时被试保持镇静状态,同时为了避免引入疲劳因素,每人每次只采集一段约3 min 的信号,重复5次,在4 种范式下共获得20 min×3×5×4 =1 200 min数据,采集频率设置为1 000 Hz,每次采集为1 组数据,对每个被试共采集80 组数据。

表1 范式要求Tab.1 Experimental paradigms
1.2 信号预处理
根据文献[14],γ 频段EEG 用于平衡分析有较好的效果,首先采用巴特沃斯带通滤波器对原始EEG 进行γ 频段信号提取。 而后使用降噪源分离技术(denoising source separation,DSS) 和小波包滤波(wavelet packet filter) 对数据进行了二重去噪处理,分别去除了眼电伪迹噪声和高斯噪声。 形成预处理EEG 数据集ri(t),i =1,2,…,19 为通道标号。
1.3 相位同步处理
相位同步处理的目的是在ri(t) 中提取人体进行平衡协调过程的数据片段,并以此组成新的数据集合Ei,i =1,2,…,19,以供后续平衡脑功能网络的搭建。 采用时间窗口长度为0.30 s 的滑动窗函数来计算目标两通道之间的PLV,计算过程如下:
将输入信号转化为式(1) 所示带有相位信息的解析信号Ri(t),i =1,2,…,19;

式中,为输入时序信号ri(t) 经过Hilbert 变换后的结果,如式(2) 所示,有

式中,ξ为柯西主值,τ为积分变量。
将Ri(t) 转换为式(3) 所示指数形式,有

式中,Ai(t) 为输入信号的瞬时振幅,θi(t) 为瞬时相位,即

上述计算获得了单个通道i中信号的相位值。 PLV代表了两个时间序列之间同步程度大小的量化值,分布范围为[0,1] ,当PLV =0 时代表两者无相位同步关系,当PLV =1 时代表两者具有完全同步的相位关系,具体计算方法如下:
通道n,m的信号表示为rn(t) 与rm(t) ,利用Hilbert 变换计算得到各自相位θn(t) 与θm(t) ,则两者相位差表示为

假设该时段内采样点总数为N,则
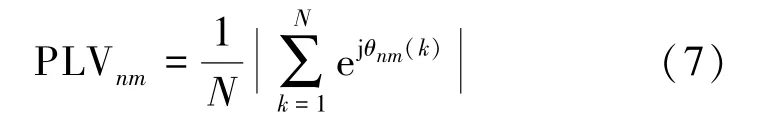
式(7) 最初的物理意义是用于评价两个非线性振子之间的耦合关系,而后研究发现神经元被激活后也存在类似的振荡行为,因而将其运用到生物信号处理,并在一些神经疾病(抑郁症、癫痫等)的预测和防治上产生了一定的效果[14]。
设置长度为2 s 的滑动时间窗,分别计算了枕叶区、额叶区和中央区中采集到的多路通道之间的PLVil,i =1,2,…,19;l =1,2,…,19;i≠l;由于本课题研究视觉和本体觉阻断情况下大脑对平衡信息的处理,因此将Fz-Pz、Fz-Cz 之间的相位同步水平作为筛选依据,当某时间段内信号相位特征符合所述筛选要求时,则提取该时段的所有通道信号加入到新的信号集合Ei,i =1,2,…,19 中,做重复但不重叠的时间窗推移直至序列结束。
1.4 传递熵网络的建立
以数据集Ei构建了一种基于传递熵值来映射人体实际神经连接的传递熵网络模型,各功能脑区节点之间的熵值变化体现了大脑的信息流通变化。大脑之间的信息流通行为具有非线性、因果性、无既定模式等特点,而传递熵网络能恰好契合该特点,传递熵值表达式为

式中,ri与rj分别为对应的两个时间序列。 已有研究表明大脑是Markov 系统[16],因此两序列的时延参数均为1。
依照式(8)对数据集Ei中两两节点进行计算,得到各个实验范式下维数为19×19 的熵值矩阵,据此矩阵构建节点连接网络图。 经过相位同步筛选后的Ei中包含了大量的平衡协调信息,所构建的网络可表征大脑内部在平衡调节时发生的改变。
1.5 人体平衡调节新特征的提取
与平衡调节密切相关的3 个脑区分别是枕叶区、中央区和额叶区,分别对应视觉感受区、运动感觉区和协同整合区,每个脑区的聚类系数的数值大小代表了该区域中整体神经元的激活程度,越大则活性越高。 为了把平衡能力统一在相同当量的信息整合能力标尺之下,计算枕叶区、中央区、额叶区的聚类系数分别为CC1、CC2、CC3,具体计算方法为

式中,ki为节点i的邻节点数量,ei为节点i与邻节点之间的实际存在边的数量,N=19 对3 个局部聚类系数求取平均值后获得全局聚类系数CC。
除此之外,以信息的协同整合区额叶区聚类系数CC3作为标尺,定义人体平衡协调新特征。 该特征也属于一种多区域协同系数,将其定义为D(下称协同系数)

式中, D1 = CC1/CC3为视觉协同系数, D2 =CC2/CC3为本体觉协同系数。
该特征在神经生理学上解释为:D1,D2 分别反映了视觉、本体觉在平衡调节中参与的程度,值越大则越紧密,其中额叶区的局部聚类系数为标准当量,以归一化不同个体的平衡能力。
1.6 实验数据的统计学验证方法
对于所采集的脑电实验数据,为排除偶然因素对实验结果的干扰,在构建传递熵网络前使用t检验对其进行了显著性检验,以确认两两范式数据间存在的差异是由于范式本身的设计所产生,而非偶然性因素。 除此之外,对分类中使用的高维组合特征采用多因素方差分析法进行了显著性检验,目的在于证明各范式下的高维数据仍然具有内在的差异性。P<0.05 被认为具有显著性差异。
2 结果
2.1 相位同步筛选结果分析
相位同步筛选结果见表2,表中数据为4 种范式下所筛选数据段中PLV 的平均值,前6 列数据分别代表了额叶区、中央区和枕叶区的内部平均同步数值,后2 列数据代表了本体觉感受区、视觉感受区与信息处理区之间的平均同步数值,可见在不同信息阻断下相应脑区的相位同步水平出现了明显的下降,与实际生理反应结果一致。

表2 4 种范式下平衡脑区PLV 表Tab.2 The PLV of different brain functional region in four paradigm
图2 则给出了某一经过筛选后数据PLV 片段。由图可知,在视觉被阻断情况下,如图2 中(b)和(d)所示,枕叶区与额叶区的同步程度波动较为剧烈,平均方差达到0.298,而中央区与额叶区的相位同步变化较为平缓,平均方差为0.171;因此得出与本体觉被阻断时相比视觉被阻断后额叶区信息流通性受到更严重的抑制,使得人体平衡调节更加困难。

图2 PLV 值波动图。 (a) Q1 范式;(b) Q2 范式;(c) Q3 范式;(d) Q4 范式Fig.2 PLV line chart.(a) Q1 paradigm; (b) Q2 paradigm; (c) Q3 paradigm; (d) Q4 paradigm
2.2 传递熵网络分析
2.2.1 网络连通性分析
如图3 所示的是基于脑区节点EEG 传递熵的连接网络图,弦图可以清晰的显示网络内部的节点集团化趋势,其中每一节点占据的圆环边缘面积体现了该节点在网络中的连通效率,占据面积越大代指由该节点产生的连通关系越多,表明其在网络中拥有更高的连通性,反之亦然;在绘制过程中已经充分的考虑到网络小世界特性,且保证了其原始邻接矩阵稀疏度为40%或更低。

图3 节点连接网络图。 (a) Q1 范式;(b) Q2 范式;(c) Q3 范式;(d) Q4 范式Fig.3 Nodes connection network diagram.(a) Q1 paradigm; (b) Q2 paradigm; (c) Q3 paradigm; (d) Q4 paradigm
分析发现,在Q1 范式下视觉相关的枕叶区O1、O2 和本体感觉相关的中央区C3、C4、Cz 附近的弦连接分布较密集,推测在获取外部信息能力正常时,大脑对于平衡的调节不需要额外的信息交互。在Q2 范式下视觉输入被阻断, O1、O2 节点以及附近节点的弦连接密集程度降低,而中央区内部节点C3、C4、Cz 等之间的联通性有增强的趋势,推测是视觉输入被阻断,枕叶区信息不足的与额叶区的信息流通程度降低,与此同时中枢神经系统增强了本体感觉等信息的整合能力,来弥补视觉缺失带来的平衡能力减弱。 在Q3 范式下本体觉输入被阻断,中央区C3、C4、Cz 节点以及附近的节点的弦连接密集程度显著降低,而枕叶区O1、O2 等节点之间的弦连接密集程度仍保持在与Q1 范式相似的较高水平。在Q4 范式下两种输入都被阻断,发现节点C3、C4、Cz 与节点O1、O2 的外部连接弦大幅减少,附近的集团化趋势减弱,表明大脑输入信息量的大幅度减少,导致脑区之间的交互性下降。
图4 显示了不同脑区在信号输入状态与输出状态下的传递熵值的分布特征。 结合两种状态发现,视觉阻断后枕叶区信号输入状态活跃程度下降,本体觉阻断后中央区信号输入状态活跃程度下降,而在两种信息都阻断的情况下额叶区的信号输出状态活跃程度有了大幅度的提升,并且显示出了较强烈的扩张趋势。

图4 传递熵特征值分布情况图。 图中(a)输入;(b)输出Fig.4 The transfer entropy box diagram.(a)Output; (b)Input
对同一脑区不同范式间的数据进行t检验以证明各组数据存在的独立性,P值如表3 所示,从中可见各范式之间存在着的显著差异(P <0.01)。

表3 t 检验的P 值Tab.3 P values in Student′s t-test
2.2.2 网络特征
对所构建传递熵网络提取了枕叶区、中央区、额叶区的聚类系数均值、方差和协同系数D1、D2 的均值和标准差如表4 所示;据表可知,在视觉输入被阻断后CC1系数显著下降, 但CC3系数则因为平衡信息补偿机制(应激效应)而得到一定提升,两者的趋势变化造成了D1 系数的不断缩小,由此可以判断D1 系数越小体现了神经中枢调节能力越强;同理当本体感觉阻断后D2 也呈现了类似的下降趋势,从生物系统的角度可以认为这是大脑的信息补偿机制起了作用。

表4 4 种实验范式下的网络指标数据(均值±标准差)Tab.4 Features value of brain network in different conditions(mean±SD)
除此之外,采用多因素方差分析法对4 种范式下的特征向量[CC,D1,D2] 进行了差异性分析,结果如表5 所示,根据结果显示各组间数据均存在显著差异(P <0.05)。 由此证明,将协同系数特征用于平衡能力评估当中是具有现实生理意义的。

表5 特征向量 [CC,D1,D2] 的差异性检测P 值表Tab.5 P-values of the feature vector [CC, D1, D2]
2.3 平衡分类结果
组合全局聚类系数CC与协同系数D1、D2 成为综合分类特征并将其作为特征输入到高斯内核的SVM 中进行分类。 图5 为各个实验范式下综合分类特征的三维空间分布图,图中Q1 范式具有较好区分性,而范式Q2、Q3 与Q4 表现出部分重合迹象。

图5 空间分类结果Fig.5 Classification result of three dimensions
利用上述特征进行分类,同时使用了两组其他分类特征进行对比,包含一组其他脑功能网络特征和一组传统足底压力中心(center of pressure, COP)特征[17]。 对比分类结果见表6,表中CC,L,D 分别代表:平均聚类系数、平均最短路径长度、平均节点度;Lng,Area,Lng/Area,DFB分别代表:中心轨迹移动长度、包络面积、单位轨迹移动长度、前后动摇轨迹长度。 结果显示,相较于第二组其他脑功能网络特征而言本文提出的方法在各个范式的分类准确度均具有更优的分类效果;与传统COP 特征相比较除了在Q2 范式低于其分类精度外,其他情况均显示出了更优的分类效果。 总体而言,人体平衡协调新特征的运用有效的提高了整体的分类精确度,印证了所提出的相位同步处理具有一定的积极效果。

表6 4 种实验范式的分类识别率(%)Tab.6 Classification rate (%) of four experimental paradigms
利用训练得到的SVM 模型对新采集的不施加干扰条件下10 组青年人数据,年龄为(23±4)岁,与10 组老年人数据,年龄为(60±10)岁,进行人体平衡评估,将人体平衡能力分为优秀( Q1)、良好(Q2,Q3)、较差(Q4)等3 类,结果如表7 所示。 [CC,L,D] 组把3 例青年组样本归为平衡能力较差,且把2例老年组样本归为优秀;传统COP 特征组把1 例老年组样本归为优秀;所提出的综合分类特征方法在青年、老年人群的平衡能力评估中,获得了与实际情况更接近的结果。 总体看来,以协同系数为基础的综合分类特征能较好地刻画不同平衡能力的人群各脑区活动规律,分类精度优于COP 特征参数,适用于不同人群的平衡评估。

表7 3 种特征下的平衡能力分类结果对比Tab.7 Comparison results of balance ability classification in three different groups features
3 讨论
相位同步水平是对大脑内部信息流通状态一种直观的评估指标[11],本研究首次将其作为大脑对于人体平衡调节参与程度的评估标准,并根据该特性对采集得到的EEG 进行筛选,获得了大量主导平衡调节的有效数据段,与未进行数据筛选的方法相比,在网络构建精度和特征区分度上体现了明显的优越性。 如图2 所示,研究不同范式下大脑的相位同步数据发现,与本体觉被阻断相比视觉被阻断后在同等长度的采样时间段内PLV 波动程度和频率显著增加,反映出大脑内部的信息流通程度的降低[12],对于同等水平的人体平衡维持需要激活更广泛的额叶区。 由此推断,视觉系统在人体平衡的调节过程中起到了更大的作用。 本研究中所用相位同步分析方法与格兰杰因果分析方法[18]相比,更加偏向对于全局脑电交互行为的考虑。 本研究计算了所有采集通道之间的锁相位值,从而可以十分清晰的判断每个时刻整体通道间的相干性行为,而每个通道之间存在的交互行为由传递熵来突出表现,因而对于网络构建的准确性具有较好的提升。
本研究首次通过传递熵方法构建了人体平衡调节下的脑功能网络模型,并将协同系数作为平衡能力评估特征之一,与COP 和脑肌电相干性特征相比,克服了单个通道脑电和肌肉电信号之间的相干性局限。 由表4 可知,协同系数在不同的实验范式下有较好的区分度,且该特征从平衡感觉信息的缺失与大脑应急机制这两个角度解释了平衡调节能力的高低,体现了在部分感觉被阻断的情况下额叶区作为信息整合区域将更大程度的参与到神经调节活动当中,从而证明将其作为平衡分类特征的研究方向是正确的。 根据图5 所示结果发现,Q1 范式与其他范式有着显著的区分度,而其他范式存在部分重叠情况,推测这是由于不同被试对于不同的感觉输入所拥有的处理能力存在差异造成的,一般拥有对某一感觉较好的调节能力的被试在缺失该感觉输入时依然能通过高效的整合协同能力维持自身的平衡,但总体的平衡表现依然呈现下降状态。
4 结论
本研究表明,在构建视觉、本体觉阻断的平衡脑功能网络时,与平衡相关的枕叶区和中央区出现活跃度下降趋势,但与平衡信息整合协同有关的额叶区活跃度增加,提示在视觉、本体觉被阻断时,大脑额叶区进一步参与到平衡调节中,并由此推断额叶区活跃度增强是维持人体平衡状态的应激反应。所定义的协同系数从平衡感觉输入和整体信息协同处理的角度较全面地反映了人体平衡能力,该特征的引入提高了人体平衡能力分类的正确率。
猜你喜欢
甘肃教育(2021年10期)2021-11-02
福建江夏学院学报(2021年6期)2021-08-10
分子影像学杂志(2021年4期)2021-07-21
浙江大学学报(理学版)(2021年1期)2021-01-26
大连民族大学学报(2020年2期)2020-06-16
中学科技(2018年9期)2018-12-19
英美文学研究论丛(2018年1期)2018-08-16
公务员文萃(2017年7期)2017-07-21
饮食科学(2017年4期)2017-05-03
健康管理(2017年3期)2017-04-20
