
乙酰哈巴苷通过Wnt信号通路诱导结肠癌HCT116细胞凋亡
2022-07-07 09:25蔡晓明
天然产物研究与开发 2022年6期
周 兴,尹 倩,黄 蓉,简 兵,蔡晓明
川北医学院 基础医学院创新实验室,南充 637100
结肠癌(colon cancer)是世界第四大致命癌症,属于常见的消化系统恶性肿瘤[1,2],遗传、肥胖、缺乏锻炼和吸烟等是患结肠癌的危险因素[3]。目前临床上主要依靠手术治疗结肠癌,由于结肠癌在早期不易发现,被确诊时已错过最佳手术时机,此时患者往往需要放化疗来改善预后[4,5]。但传统化疗药物不良反应多,而天然药物的不良反应少,且更为廉价[6]。
筋骨草(AjugaciliataBunge.)是多年生唇形科草本植物,主要分布于中国河北、山东、陕西、甘肃、四川等地。筋骨草全草皆可入药,在中国民间广泛被用于治疗肺热咯血、跌打损伤、扁桃腺炎、咽喉炎等疾病,而筋骨草中乙酰哈巴苷(8-O-acetylharpagide,8-OA)的含量最高。根据You等[7]前期研究显示,乙酰哈巴苷具有强大的抗炎作用,可以减轻毛细血管早期的炎性渗出,由于炎症介质本身可以在肿瘤进展中发挥调节作用,因此乙酰哈巴苷可能通过炎症途径发挥抗肿瘤作用,此外,Konoshima等[8]则发现乙酰哈巴苷对一氧化氮诱发的小鼠皮肤癌有明显的抑制作用,并可抑制N-亚硝基二乙胺和苯巴比妥诱发的肝癌细胞增殖,表明乙酰哈巴苷可能是一种潜在的抗肿瘤药物。
本研究旨在通过乙酰哈巴苷作用于结肠癌HCT116细胞后的一系列生物学行为改变来探讨乙酰哈巴苷在抗肿瘤方面所发挥的作用。
1 材料和方法
1.1 材料
1.1.1 细胞及药物
人结肠癌细胞株HCT116购自中国科学院上海细胞库,培养条件为含10%胎牛血清的DMEM培养基,37 ℃,5% CO2和95%湿度。乙酰哈巴苷(成都曼思特生物技术有限公司,纯度≥97%,货号:6926-14-3),溶解于无菌PBS中配置成浓度为2.5 mol/L的储存液。
1.1.2 试剂
DMEM培养基、胰蛋白酶(美国HyClone公司);胎牛血清(美国Gibco公司);培养瓶、96孔板(美国Corning公司);CCK8(上海东仁化学科技有限公司);细胞凋亡试剂盒(中国碧云天公司);Total RNA Extractor(上海生工生物公司);cDNA反转录试剂盒(美国Thermo Fisher Scientific公司);SYBR Green PCR Master Mix(北京索莱宝公司);引物合成(上海生工生物公司);XAV939(中国MCE公司);兔抗人IgG一抗、羊抗兔IgG二抗(中国ProteTech公司)。
1.1.3 仪器设备
3111 CO2培养箱(美国Thermo Fisher Scientific公司);SW-0J-1FD 超净工作台(中国安泰空气技术有限公司);AE 2000 倒置显微镜(日本奥林巴斯公司);BCD-272WDGD-20 ℃冷冻冰箱(中国海尔公司);EasyCyte6-2L流式细胞仪(美国Guava公司);CFX connect荧光定量PCR仪(美国Bio-Rad公司);5804R高速低温离心机(美国Thermo Fisher Scientific公司);LUX5187139全波长酶标仪(美国Thermo Fisher Scientific公司);Chemidoc XRS+凝胶成像仪(美国BIO-RAD公司)。
1.2 方法
1.2.1 CCK8
收集HCT116细胞,离心计数后调整密度为1×105个/mL,每孔100 μL铺入96孔板,待细胞贴壁,实验组分7组,每组含乙酰哈巴苷的浓度分别为0.125、0.25、0.5、0.75、1、1.25、1.5 mmol/L,对照组加入与实验组等体积的PBS溶液,全程避光加样,完成后放入37 ℃,5% CO2培养箱中培养24、48、72 h;第二天每孔加入10 μL CCK8试剂,用酶标仪测定OD450值,计算抑制率。
1.2.2 平板克隆形成实验
在6孔板中每孔加入HCT116细胞500个,每组3个复孔。实验组以0.5 mmol/L乙酰哈巴苷处理,对照组加入与实验组等体积的PBS溶液,在作用HCT116细胞7天后,吸掉培养基,PBS洗涤细胞3次,4%多聚甲醛固定细胞5 min,再用PBS洗涤细胞3次。500 μL结晶紫染色细胞20 min,PBS洗涤后拍照记录。
1.2.3 流式细胞仪检测细胞周期和凋亡
将细胞计数后调整密度为1×105个/mL,每孔2 mL铺入6孔板,实验孔药物浓度为(0.25、0.5、0.75 mmol/L),对照组加入与实验组等体积的PBS溶液,培养48 h后收集培养液,PBS洗涤细胞一次,加入胰酶消化2 min,加入之前收集的细胞培养液,转移到离心管内,1 000 r/min离心5 min,弃上清,用PBS重悬并计数,周期实验加入1 mL冰浴70%乙醇,混匀后4 ℃固定24 h,1 000 r/min离心5 min,弃上清,加入1 mL PBS离心后弃上清,每管加0.5 mL碘化丙啶染料37 ℃避光孵育30 min;凋亡实验取5×104个细胞,1 000 r/min离心5 min,弃上清,加入195 μL Annexin V-FITC结合液重悬细胞,再加入5 μL AnnexinV-FITC染色液,混匀,加入10 μL碘化丙啶染色液,混匀,室温避光孵育10 min,置于冰上,立即用流式细胞仪检测。
1.2.4 转录组测序
将细胞计数后调整密度为1×105个/mL,每孔2 mL铺入6孔板中培养,实验组(B1、B2、B3)加入乙酰哈巴苷的终浓度为0.5 mmol/L,对照组(A1、A2、A3)加入与实验组等体积的PBS溶液,培养48 h后收集细胞,用Trizol法提取总RNA,以Illumina HiseqTM测序平台进行测序分析。
1.2.5 乙酰哈巴苷对HCT116细胞mRNA表达的影响
将细胞计数后调整密度为1×105个/mL,每孔2 mL铺入6孔板中培养,细胞贴壁后弃原培养基,实验组加入乙酰哈巴苷的终浓度分别为0.25、0.5 mmol/L,对照组加入与实验组等体积的PBS溶液,培养48 h后收集细胞,用Trizol法提取总RNA,取1 μg总RNA为模板进行逆转录反应,合成cDNA,反应体系为20 μL。先将RNA模板和Random primer加在一起,补充RNase-free ddH2O至12 μL,进行热变性(条件:65 ℃,5 min,冰上1 min),再加入其他组分,在漩涡混合器上充分混匀,4 000 r/min,15 s离心;放入PCR扩增仪中,设置反应程序:25 ℃ 5 min,42 ℃ 60 min,70 ℃ 5 min,冰上1 min进行逆转录合成cDNA,模板稀释20倍后进行PCR扩增;体系:SYBR Mixture 12.5 μL,F-primer 1 μL,R-primer 1 μL,cDNA 5 μL,DDH2O 5.5 μL;PCR仪器设置为:95 ℃预变性10 min,95 ℃变性15 s,60 ℃退火60 s,共进行40个循环。引物序列见表1。
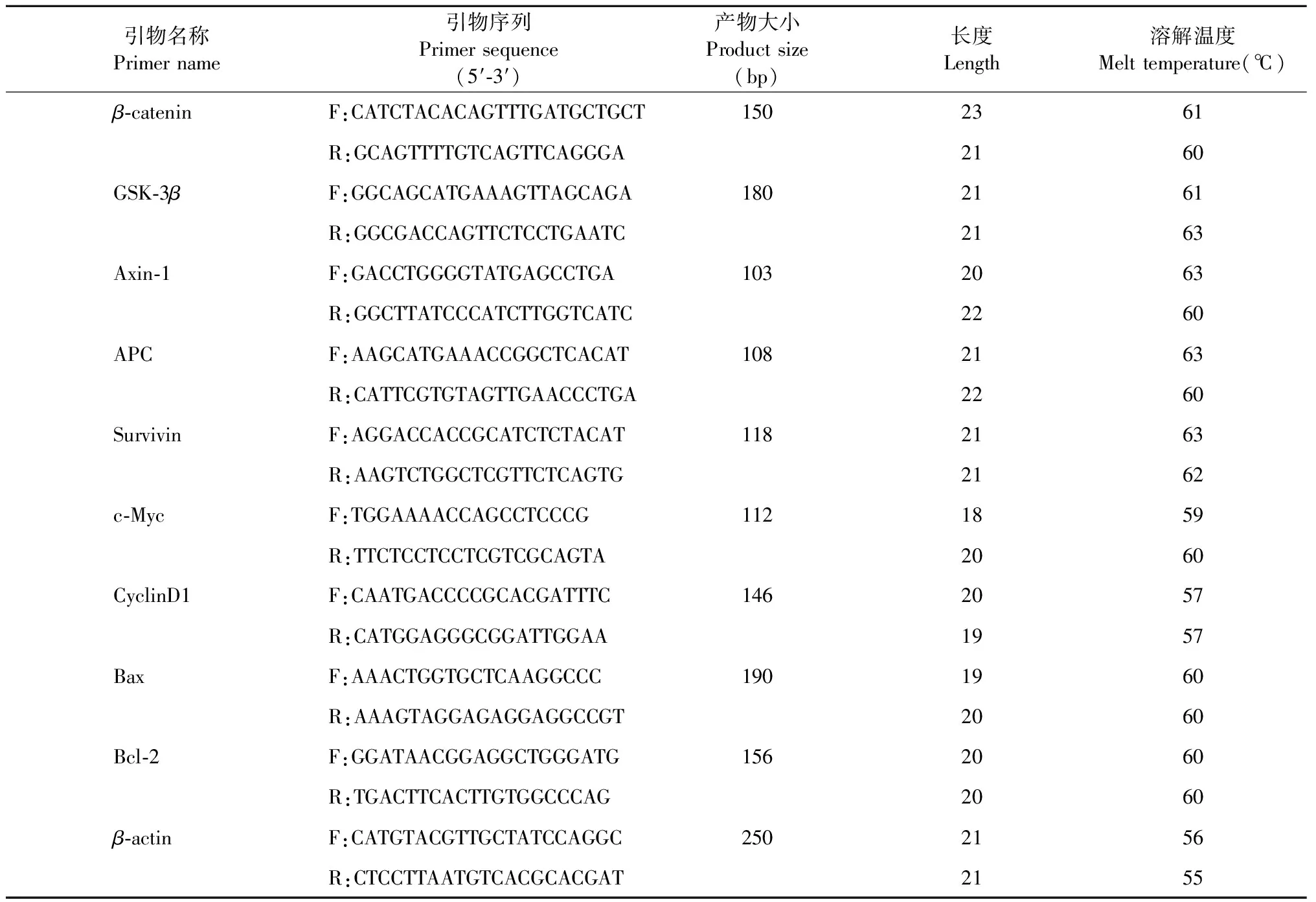
表1 引物序列
1.2.6 乙酰哈巴苷对HCT116细胞蛋白表达的影响
培养细胞方法和分组同“1.2.5”,48 h后收集细胞加入RIPA裂解液提取总蛋白。配置SDS-PAGE凝胶,100 V电泳2 h左右,直至Marker清晰分离开即可终止电泳,80 V转膜1.5 h后将PVDF膜置于5%脱脂奶粉封闭液中,常温封闭4 h,用TBST洗3次。在4 ℃冰箱中摇动孵育一抗过夜,用TBST在室温下洗膜3次,室温下振摇孵育二抗1 h,用TBST洗3次,用Bio-Rad凝胶成像仪显影。
1.2.7 统计学处理
流式数据采用FlowJo 7.6分析,Western blot采用Image J对条带光密度进行定量,所有数据均采用SPSS 20.0进行统计分析处理,采用单因素方差分析,计量数据采用均数±标准差表示,P<0.05表示差异显著,P<0.01表示差异极显著。
2 结果
2.1 乙酰哈巴苷对结肠癌细胞增殖情况的影响
CCK8法检测结果显示,结肠癌HCT116细胞的抑制率随乙酰哈巴苷浓度的升高而显著增加(P<0.05),乙酰哈巴苷浓度分别为0.125、0.25、0.5、0.75、1、1.25、1.5 mmol/L时,作用HCT116细胞24 h后的抑制率如图1所示,分别为(2.22±0.82)%、(8.24±0.90)%、(31.18±6.48)%、(37.12±4.79)%、(52.71±1.96)%、(60.76±3.79)%、(68.65±3.22)%。结果表明乙酰哈巴苷可呈浓度-时间依赖性地抑制结肠癌细胞体外增殖。作用24、48、72 h的半数抑制浓度(IC50)分别为0.91、0.39、0.39 mmol/L,由于48 h至72 h其IC50值相同,表明乙酰哈巴苷主要在最初的48 h内发挥作用,因此后续取48 h为最佳作用时间。

图1 乙酰哈巴苷以时间依赖的方式抑制HCT116细胞活性
2.2 乙酰哈巴苷对结肠癌细胞克隆形成的影响
用0.5 mmol/L乙酰哈巴苷作用HCT116细胞7天后,对照组细胞形成了明显的克隆(197±16),而药物处理组的克隆率明显减少(0.33±0.57)。由图2所示,差异有统计学意义(P<0.05),表明乙酰哈巴苷能显著降低HCT116细胞的分裂能力,干扰细胞的克隆形成。

图2 乙酰哈巴苷显著抑制HCT116细胞增殖
2.3 乙酰哈巴苷对结肠癌细胞周期和凋亡的影响
通过流式细胞仪对染色细胞进行分群,从而判断细胞所处的周期相和凋亡相。结果如图3所示,随着乙酰哈巴苷浓度的逐渐增加,G0/G1期细胞比例由(59.40±0.98)%增加到(85.75±0.84)%,细胞周期被阻滞在G1期。

图3 乙酰哈巴苷以剂量依赖的方式抑制HCT116细胞周期
细胞凋亡实验结果如图4,显示0.75 mmol/L乙酰哈巴苷处理HCT116细胞48 h后,早期凋亡细胞从(1.21±0.38)%增加到(17.84±0.41)%,差异有统计学意义(P<0.05),表明乙酰哈巴苷可以诱导HCT116细胞发生凋亡反应。
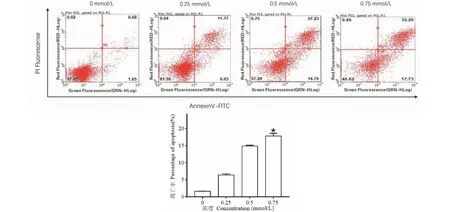
图4 乙酰哈巴苷对HCT116细胞凋亡的影响
2.4 转录组测序筛选乙酰哈巴苷作用HCT116细胞的靶向信号通路
转录组测序结果数据采用DESeq进行分析,结果如图5所示,对照组与药物治疗组相比存在4 129个差异基因,其中下调基因2 208个,上调基因1 921个。
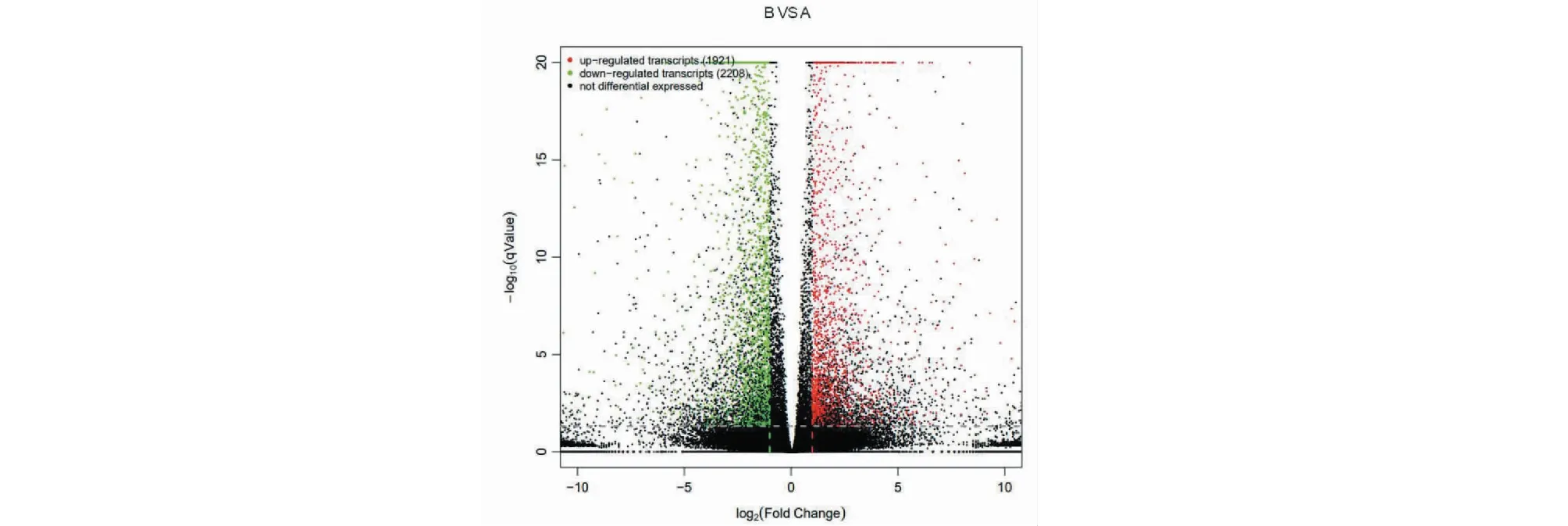
图5 转录组测序火山图
通过KEGG数据库功能富集,结果如图6,发现Wnt通路是有显著差异的通路,表明Wnt通路可能是乙酰哈巴苷的靶向信号通路。

图6 KEGG通路富集结果
显著富集功能蛋白质互作分析结果如图7,显示乙酰哈巴苷影响的主要差异蛋白分子有PLK1、KIF11、CDC20、WNT16、NOTHCH3、BUB1、NOXA1、MAPK15等,这些分子的功能主要是参与调节细胞周期、有丝分裂、DNA氧化应激损伤应答和细胞代谢等。

图7 蛋白质互作网络
2.5 乙酰哈巴苷对HCT116细胞Wnt信号通路基因mRNA表达的影响
为验证乙酰哈巴苷对HCT116细胞Wnt信号通路的影响,对Wnt信号通路相关基因进行检测,实验结果如图8。与对照组相比,乙酰哈巴苷组(0.25、0.5 mmol/L)显著降低了HCT116细胞β-catenin、c-Myc mRNA的表达(P<0.05)。与细胞周期、凋亡相关基因CyclinD1、Survivin、Bcl-2显著降低,Bax mRNA显著上调(P<0.05),以上结果表明乙酰哈巴苷在一定程度上抑制了Wnt通路的转录活性,同时上调了促凋亡基因的转录并下调了凋亡抑制基因的转录。
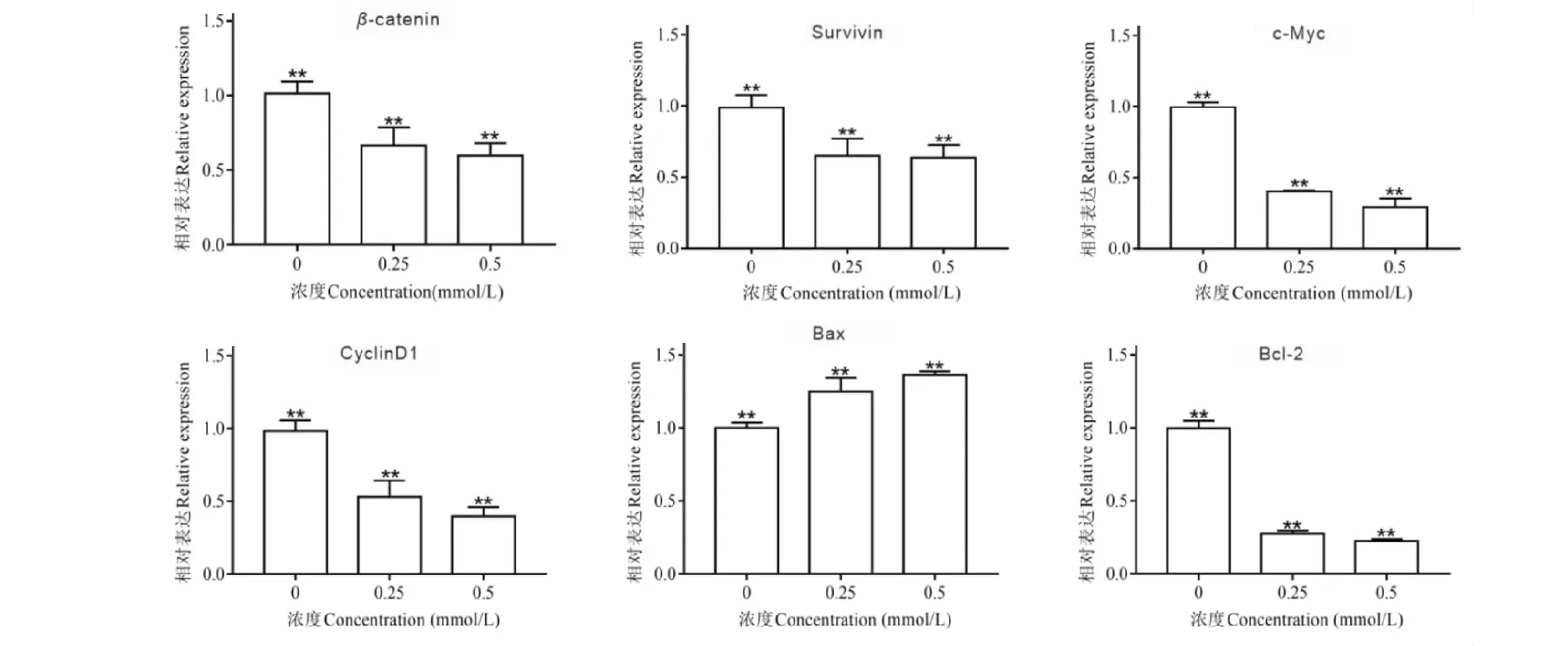
图8 乙酰哈巴苷对HCT116细胞中Wnt信号通路及细胞凋亡相关基因mRNA水平的影响
2.6 乙酰哈巴苷对HCT116细胞Wnt通路蛋白的影响
为验证乙酰哈巴苷对HCT116细胞Wnt信号通路蛋白的影响,用乙酰哈巴苷作用HCT116细胞,检测结果如图9,显示乙酰哈巴苷引起Wnt信号通路及细胞周期相关蛋白β-catenin、c-Myc、Survivin、CyclinD1、Bcl-2、pro-caspase3显著下调(P<0.05),乙酰哈巴苷引起细胞凋亡相关蛋白APC、Bax、cleaved-caspase3显著上调(P<0.05)。

图9 乙酰哈巴苷处理后Wnt信号通路相关蛋白的表达
同时,用XAV939和乙酰哈巴苷各自单独用药后对HCT116细胞进行检测发现都能显著降低β-catenin和c-Myc蛋白的表达,二者联合用药后如图10所示,能进一步降低β-catenin和c-Myc蛋白的表达并降低了HCT116细胞的存活率,差异有统计学意义(P<0.05),这些结果从蛋白水平上表明乙酰哈巴苷抑制了Wnt通路的活性,同时激活了凋亡通路。乙酰哈巴苷与XAV939具有协同作用,能进一步抑制HCT116细胞的Wnt/β-catenin信号通路。
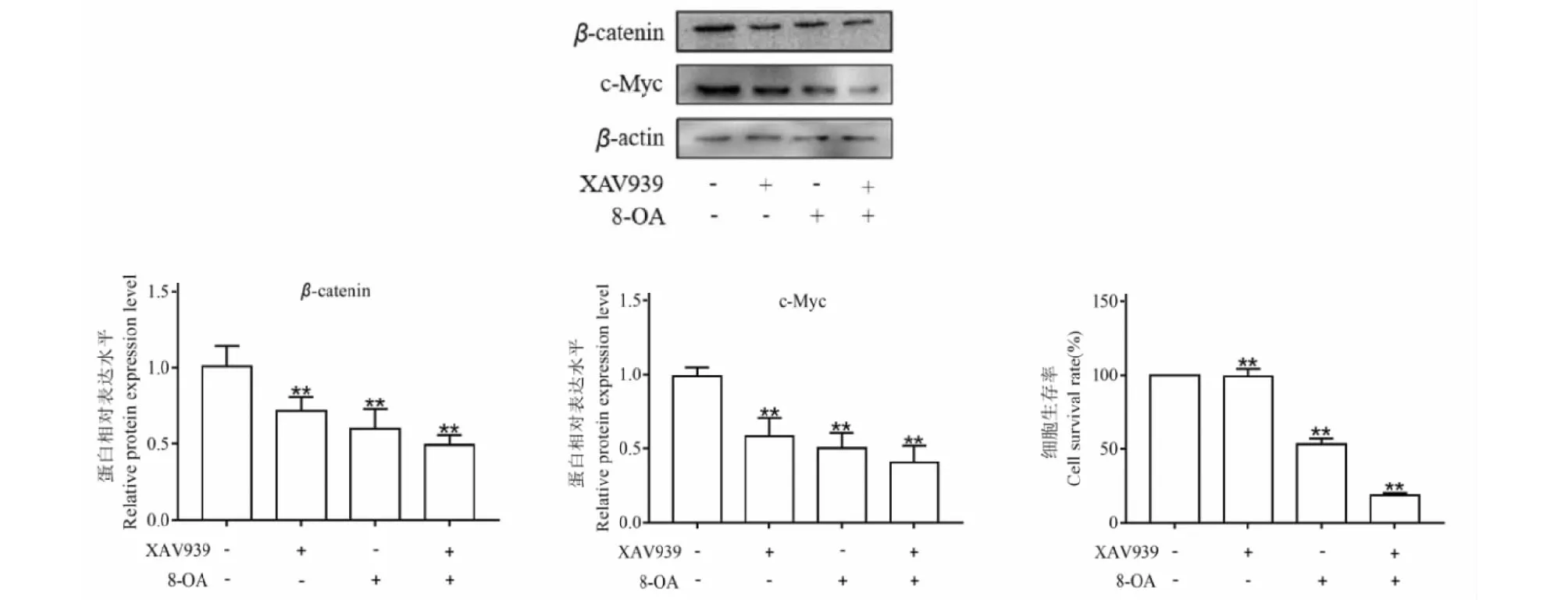
图10 乙酰哈巴苷联合XAV939处理对β-catenin、c-Mcy表达的影响
3 讨论与结论
结肠癌约占全世界每年癌症相关死亡人数的10%[9]。目前结肠癌的治疗主要以手术为主,化疗和放疗为辅,对于晚期化疗患者来说,化疗的有效率低且副作用明显,生存期只有18个月左右[10]。近年来,在天然植物中寻找能抗肿瘤药物是一大研究热点,因为一些传统的化疗药物,包括紫杉醇和长春新碱[11]均来源于自然植物。
抗增殖是开发抗癌药物的重要切入点,CCK8法显示了乙酰哈巴苷对HCT116细胞的抑制作用呈浓度-时间依赖性,三个时间点的IC50分别为0.91、0.39、0.39 mmol/L,说明乙酰哈巴苷主要在最初的48 h内发挥作用。此外,通过平板克隆形成实验证明了乙酰哈巴苷可导致HCT116细胞的克隆形成能力明显减弱。
肿瘤细胞的增殖和生存能力远强于正常细胞[12],大多数抗癌药物能通过诱导细胞周期阻滞和细胞凋亡来抑制肿瘤细胞增殖[13]。细胞在死亡过程常发生活性氧水平升高、半胱天冬氨酸蛋白酶激活和染色质凝聚等现象[14],其中线粒体跨膜电位丢失诱导细胞色素c从线粒体释放到胞质是诱导凋亡的关键步骤[15],而Bax和Bcl2对细胞色素c的释放和下游caspase蛋白的激活至关重要[16]。对此我们通过流式细胞仪检测发现,乙酰哈巴苷可以剂量依赖性地影响结肠癌HCT116细胞的细胞周期,引起细胞周期阻滞于G1期,同时导致细胞的凋亡率明显升高。
研究证实Wnt信号通路与结肠癌发展高度相关[17],其中,β-连环蛋白(β-catenin)是Wnt经典通路的关键分子,E-钙黏蛋白/β-连环蛋白复合物有助于细胞间粘附稳定,从而减少细胞的侵袭和转移[18]。在肿瘤细胞中,β-catenin可激活与癌细胞增殖、周期、凋亡、迁移、侵袭和耐药性相关的基因转录,从而导致肿瘤的恶性增殖[19]。此外,β-catenin已被证明是一种有前途的癌症预防和治疗靶点,许多天然产物可作为β-catenin信号传导的抑制剂,其机制主要是通过磷酸化、泛素化和抑制其核移位实现的,并且天然产物抑制剂在体内外的各种肿瘤模型中显示出了较好的预防和治疗效果[20]。β-catenin在细胞核中可促进下游分子Myc的表达,Myc属原癌基因家族,在核内编码细胞必需的核转录因子,主要参与调节细胞的生长、增殖、分化、周期、代谢、凋亡等[21]。在哺乳动物中,Myc家族蛋白质包含c-Myc、n-Myc和l-Myc三类,c-Myc与基因组不稳定性和肿瘤发生有关。在静止细胞中,c-Myc的表达水平较低,一旦细胞进入细胞周期,c-Myc的表达迅速增加数倍[22]。此外,在约70%的结肠癌中发现c-Myc蛋白显著上调,表明c-Myc与结肠癌的恶性程度呈正相关[23]。
本实验采用高通量转录组测序发现乙酰哈巴苷共引起HCT116细胞产生4 129个差异基因,其中分别为上调基因1 921个,下调基因2 208个,差异基因主要与细胞生长、细胞周期、细胞侵袭、细胞免疫、氧化应激、细胞代谢、分子转运和细胞死亡途径有关,在差异基因结合KEGG数据库分析后发现,乙酰哈巴苷显著影响了HCT116细胞的Wnt信号通路,通过乙酰哈巴苷与对照组基因表达和蛋白质检测,证实了β-catenin和c-Myc的显著下调。乙酰哈巴苷联合Wnt小分子抑制剂XAV939进一步抑制了HCT116结肠癌细胞的生长,表明Wnt/β-catenin通路还介导了HCT116的耐药性,抑制Wnt/β-catenin通路可使乙酰哈巴苷的抗肿瘤活性明显增强。
本研究显示,乙酰哈巴苷能有效抑制结肠癌细胞HCT116的增殖活性,基因组检测发现细胞的Wnt/β-catenin信号通路发生显著差异,推测乙酰哈巴苷可能通过调控Wnt/β-catenin信号通路使细胞正常的生长受到抑制,进一步诱导细胞凋亡和周期阻滞,从而抑制肿瘤细胞生长。然而,该推测需要佐证,其具体作用途径尚需进一步深入研究。
猜你喜欢
分子催化(2022年1期)2022-11-02
中国典型病例大全(2022年9期)2022-04-19
皮肤性病诊疗学杂志(2021年5期)2021-11-27
现代牧业(2021年4期)2021-04-14
智慧健康(2021年33期)2021-03-16
三农资讯半月报(2021年1期)2021-01-27
大自然探索(2017年12期)2018-03-02
中国医药导报(2016年33期)2017-03-06
现代养生·下半月(2015年8期)2015-11-16
中国民族民间医药·下半月(2014年4期)2014-09-26
