
lncRNA NKILA靶向miR-22-3p调控高糖诱导的人足细胞增殖和凋亡
2022-05-26 09:45张艳秋翟铁刘丹郝凤杰王晓军承德市中心医院河北承德067000
中国老年学杂志 2022年10期
张艳秋 翟铁 刘丹 郝凤杰 王晓军 (承德市中心医院,河北 承德 067000)
足细胞在糖尿病肾病(DN)的发病机制中起关键作用,高糖可诱导足细胞凋亡和脱落,最终导致肾小球硬化〔1〕。研究足细胞凋亡和损伤机制有助于治疗DN。核因子(NF)-κB相关长链非编码RNA(lncRNA NKILA)在10 mg/脂多糖( LPS)诱导8 h的脓毒症肾损伤的HK-2细胞中表达显著升高〔2〕。而本研究通过StarBase预测显示,miR-22-3p可能是lncRNA NKILA的靶基因。miR-22-3p在肾小管损伤模型细胞〔3〕和脓毒症诱导的急性肾损伤〔4〕中表达下调。且有研究表明miR-22-3p可通过磷脂酰肌醇-3-激酶(PI3K)/蛋白激酶B(AKT)途径影响肾癌细胞活力和迁移〔5〕。根据上述研究结论和预测结果本研究假设,在高糖诱导的人足细胞HPC中,lncRNA NKILA靶向miR-22-3p通过PI3K/AKT途径影响高糖诱导的人足细胞HPC增殖和凋亡,并对此假设进行验证。
1 材料与方法
1.1材料 人永生性肾脏足细胞系HPC购自北京北纳生物;McCoy 5a培养基和胎牛血清(FBS)购自美国Gibco公司;胰蛋白酶Trypsin购自Sigma-Aldrich公司;细胞周期蛋白(cyclin)D1抗体、cleaved-caspase-3抗体、磷酸化(p)-PI3K、p-AKT抗体和β-actin抗体购自英国Abcam公司;Lipofectamine 2000转染试剂、总RNA提取试剂Trizol、 real-time PCR试剂盒、反转录试剂盒(RT-PCR)购自美国Invitrogen公司;流式细胞凋亡检测试剂盒购自美国BD公司;lncRNA NKILA抑制剂(anti-lncRNA NKILA)、lncRNA NKILA过表达载体(pcDNA-NKILA)、miR-22-3p mimics、miR-22-3p抑制物(anti-miR-22-3p)、阴性对照(si-NC、pcDNA-NC、miR-NC和anti-miR-NC)、lncRNA NKILA的野生型(WT-NKILA)和突变型(MUT-NKILA)双荧光报告载体购自上海吉玛基因;双荧光素酶报告系统购自美国Promega公司;流式细胞仪购自美国BD公司,real-time PCR仪购自美国Bio-Rad公司。
1.2方法
1.2.1细胞培养、分化和分组〔6〕细胞培养和分化:将HPC培养在含10%FBS、100 U/ml青霉素和100 mg/ml链霉素的McCoy 5a培养基中,置37℃ 5%CO2、湿度95%的培养箱中培养,2 d传代一次,细胞传代时将细胞培养于33℃ 5%CO2培养箱中,细胞基本融合为一层时,将传代的HPC细胞转入37℃ 5% CO2培养箱培养10~14 d,细胞分化成熟。
实验分组:正常对照(NC)组:McCoy 5a培养基+D-葡萄糖11.1 mmol/L培养48 h;高糖刺激(HG)组:McCoy 5a培养基+D-葡萄糖25 mmol/L培养48 h;高糖刺激+转染(HG+转染)组:转染48 h后用McCoy 5a培养基+D-葡萄糖25 mmol/L处理48 h。
1.2.2细胞转染 转染前24 h将HPC以5×105个细胞/孔接种于6孔板中,培养至细胞融合度为50%~60%时根据转染说明书进行转染。转染分组:lncRNA NKILA低表达组(转染si-NKILA,以si-NC为对照),miR-22-3p高表达组(转染miR-22-3p,以miR-NC为对照),lncRNA NKILA和miR-22-3p双抑制组(转染si-NKILA+anti-miR-22-3p,以si-NKILA+anti-miR-NC为对照)。将无血清培养液稀释的等体积脂质体和各组载体混合,室温孵育20 min,加入到培养好的HPC细胞中,培养6 h,弃无血清培养液,加入McCoy 5a完全培养基,转染48 h,收集细胞。
1.2.3real-timePCR检测lncRNA NKILA和miR-22-3p 的表达 收集NC组、HG组或转染的各组HPC细胞,按照总RNA提取试剂盒的说明书提取细胞总RNA,反转录合成cDNA,取cDNA为模板按照 real-time PCR的说明书进行反应合成lncRNA NKILA和miR-22-3p,反应程序为:95℃ 2 min;95℃30 s、60℃ 40 s、72℃ 45 s,40个循环;72℃ 5 min。用2-ΔΔCt方法进行数据分析。
1.2.4CCK-8实验检测细胞存活率 收集转染后和(或)高糖处理的HPC细胞,消化并稀释,以2×103个细胞/孔(100 μl细胞/孔)接种于96微孔板中,在细胞培养至48 h进行 CCK-8实验,每孔加入10 μl CCK-8溶液,置于37℃继续培养2 h,酶标仪检测450 nm 处的吸光度(A)值。细胞存活率(%)=实验组A值/阴性对照组A值×100%。
1.2.5Western印迹检测蛋白表达 收集各组HPC细胞,裂解破碎细胞,收集细胞中的蛋白,测定蛋白浓度。将蛋白样本进行十二烷基硫酸钠-聚丙烯酰胺凝胶电泳(SDS-PAGE),转聚偏氟乙烯(PVDF)膜,封闭2 h,加入稀释的各个一抗(cyclinD1抗体1∶2 000、cleaved-caspase-3抗体1∶1 000、p-PI3K抗体1∶1 000、p-AKT抗体1∶1 000、和β-actin抗体1∶4 000),4℃过夜孵育,洗膜2次,加入稀释的酶标记二抗,室温孵育2 h,以β-actin为内参,分析蛋白表达水平。
1.2.6流式细胞术测定细胞凋亡率 收集经高糖或(和)转染后各组HPC细胞,胰酶消化,离心收集细胞,按照膜联蛋白(Annexin)V/碘化丙啶(PI)试剂盒说明书进行操作,Annexin V-FITC和PI各5 μl,混匀,室温避光15 min,流式细胞仪检测凋亡率。
1.2.7双荧光素酶报告实验 根据1.2.2进行HPC细胞培养和转染,将构建的WT-NKILA和MUT-NKILA双荧光素酶报告载体分别与miR-NC或miR-22-3p共转染HPC细胞,转染后培养48 h,收集细胞并进行裂解,离心收集上清,根据试剂盒说明书进行操作,以海肾荧光素酶活性为内参照,检测分析相对萤火虫荧光素酶活性。
1.3统计学处理 采用SPSS21.0软件进行单因素方差分析和t检验。
2 结 果
2.1NC组和HG组中lncRNA NKILA和miR-22-3p表达情况 与NC组相比,HG组的HPC细胞lncRNA NKILA含量显著升高(P<0.05),miR-22-3p显著降低(P<0.05),见表1。

表1 NC组和HG组中lncRNA NKILA和 miR-22-3p表达情况
2.2低表达NKILA对高糖处理的HPC增殖和凋亡影响 与NC组相比,HG组的HPC细胞存活率和cyclinD1含量降低,凋亡率和cleaved-caspase-3蛋白含量上升,差异均具有统计学意义(P<0.05);与HG+si-NC组相比,HG+si-NKILA组HPC细胞中NKILA含量降低,细胞存活率和cyclinD1含量升高,凋亡率和cleaved-caspase-3蛋白含量降低,差异均具有统计学意义(P<0.05),见图1、图2和表2。
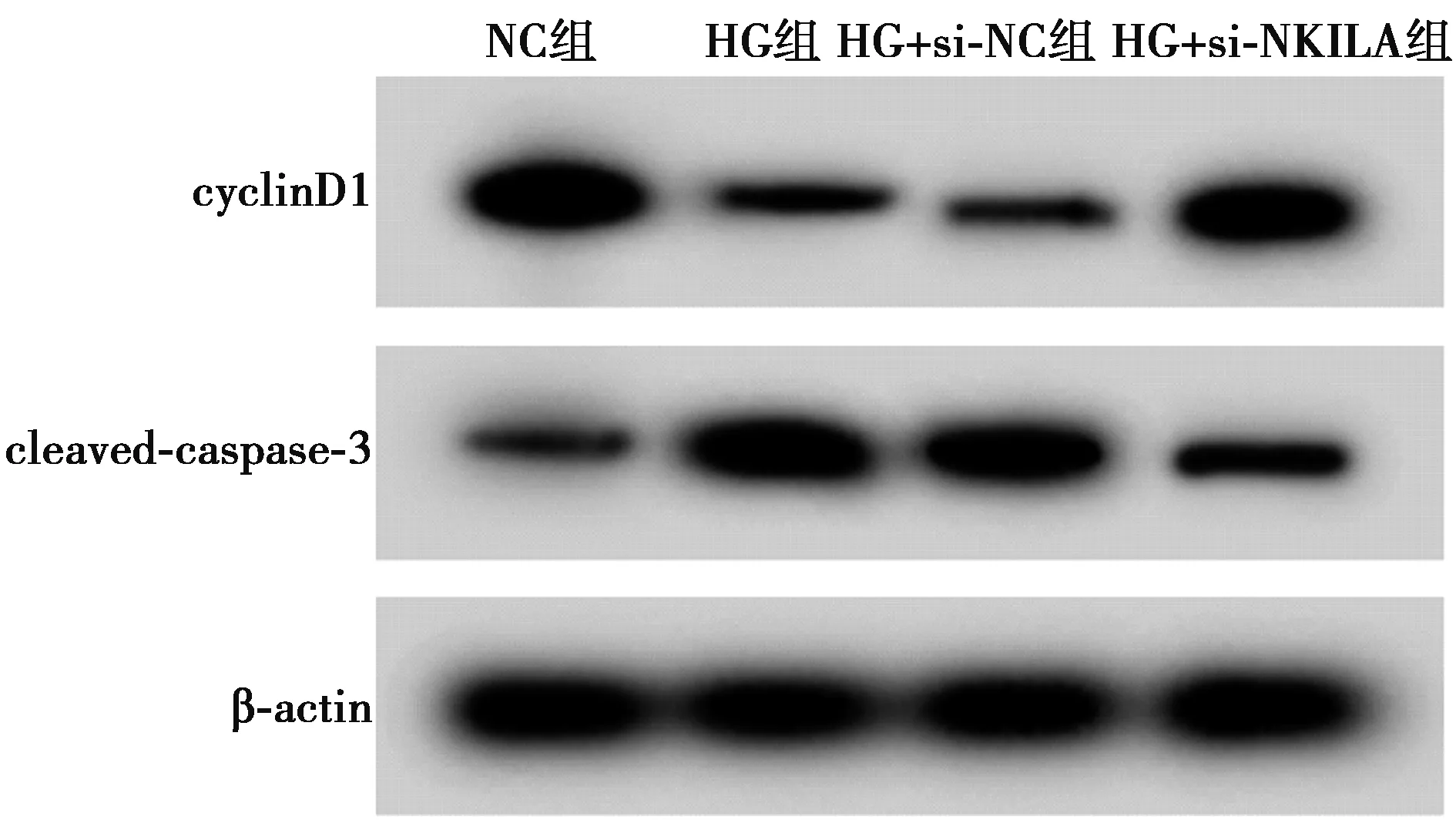
图1 Western印迹检测cyclinD1和cleaved-caspase-3 蛋白表达
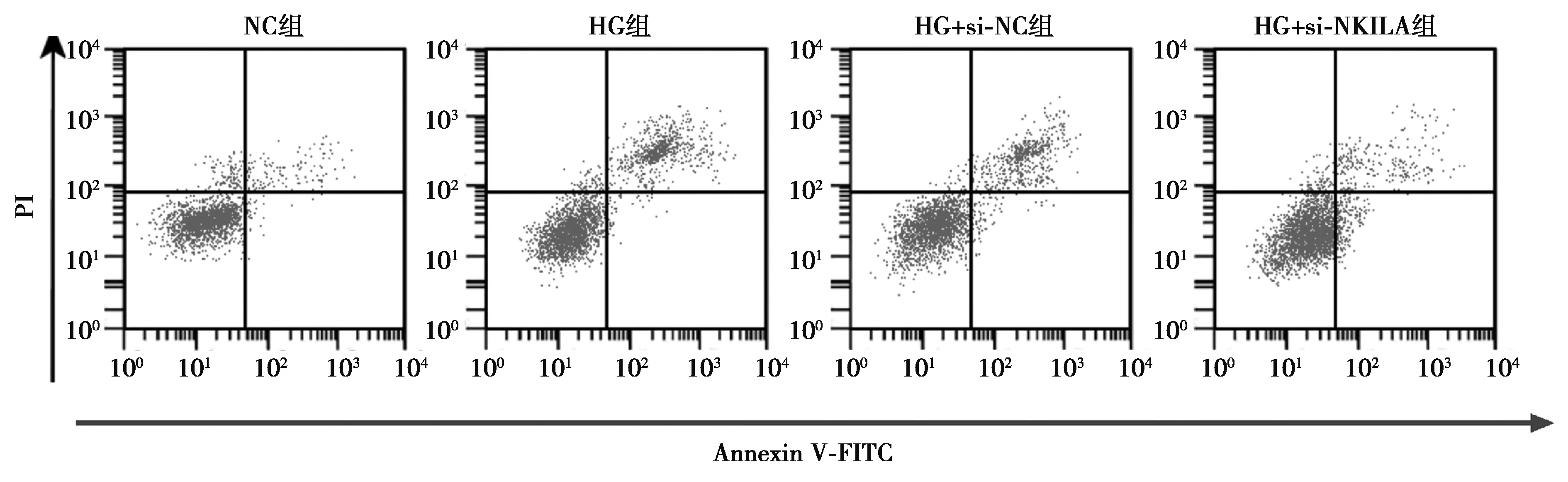
图2 流式细胞仪检测细胞凋亡
表2 低表达NKILA对高糖诱导的HPC增殖和凋亡影响
2.3高表达miR-22-3p对HPC增殖和凋亡的影响 与HG+miR-NC组相比,HG+miR-22-3p组HPC细胞中miR-22-3p含量、细胞存活率和cyclinD1含量升高,凋亡率和cleaved-caspase-3蛋白含量降低,差异均具有统计学意义(P<0.05),见图3和表3。
2.4lncRNA NKILA靶向调控miR-22-3p的表达 通过StarBase预测显示,miR-22-3p序列中含有与lncRNA NKILA互补的位点,见图4。与miR-NC组相比,miR-22-3p组WT-NKILA的萤火虫荧光素酶相对活性显著降低(P<0.05),而MUT-NKILA的荧光素酶相对活性无明显变化(P>0.05)。见表4。qRT-PCR结果表明,si-NC组、si-NKILA组、pcDNA-NC组、pcDNA-NKILA组miR-22-3p分别为:1.02±0.11、2.69±0.27、1.05±0.10、0.42±0.04。抑制lncRNA NKILA可上调miR-22-3p含量,过表达lncRNA NKILA可下调miR-22-3p含量(F=353.702,P<0.001)。
2.5低表达miR-22-3p可以部分逆转NKILA低表达对HPC增殖和凋亡的影响 与HG+si-NKILA+anti-miR-NC组相比,HG+si-NKILA+anti-miR-22-3p组HPC细胞中miR-22-3p、细胞存活率和cyclinD1含量均降低,凋亡率和cleaved-caspase-3蛋白含量升高,均具有统计学意义(P<0.05),见图5和表5。
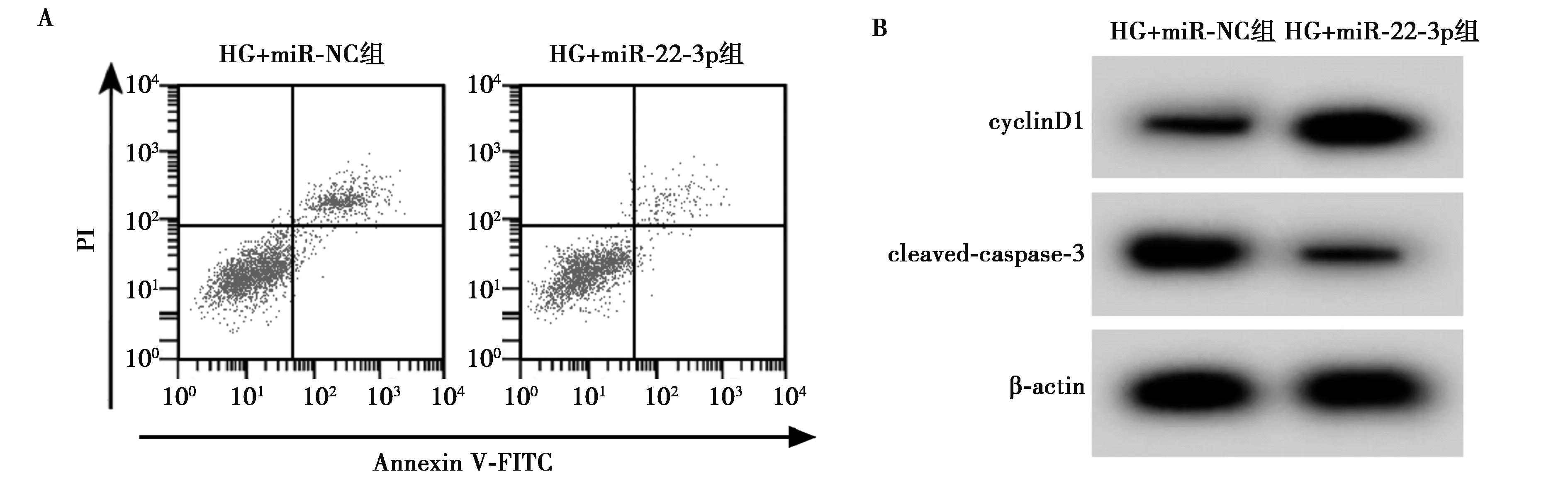
A:流式细胞仪检测细胞凋亡;B:Western印迹检测cyclinD1和cleaved-caspase-3蛋白的表达图3 高表达miR-22-3p对HPC的增殖和凋亡的影响
表3 高表达miR-22-3p对HPC的增殖和凋亡的影响

图4 通过Starbase预测miR-22-3p和 NKILA结合示意图
表4 miR-NC或miR-22-3p与报告质粒共转染HPC 细胞后双荧光素酶活性检测
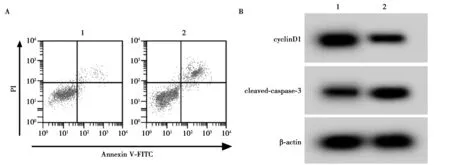
A:流式细胞仪检测细胞凋亡;B:Western印迹检测cyclinD1和cleaved-caspase-3蛋白的表达;1、2:HG+si-NKILA+anti-miR-NC组、HG+si-NKILA+anti-miR-22-3p组图5 低表达miR-22-3p可以部分逆转NKILA低表达对HPC增殖和凋亡的影响
表5 低表达miR-22-3p可以部分逆转NKILA低表达对HPC增殖和凋亡的影响
2.6PI3K/AKT信号通路相关蛋白的表达 与HG+si-NC组相比,HG+si-NKILA组HPC细胞中p-PI3K和p-AKT含量均显著升高(P<0.05);与HG+si-NKILA+anti-miR-NC组相比,HG+si-NKILA+anti-miR-22-3p组p-PI3K和p-AKT含量均显著下降(P<0.05),见图6和表6。

1~4:HG+si-NC组、HG+si-NKILA组、HG+si-NKILA+anti-miR-NC组、HG+si-NKILA+anti-miR-22-3p组图6 Western印迹检测p-PI3K、p-AKT蛋白的表达
表6 PI3K/AKT信号通路相关蛋白的表达
3 讨 论
大量lncRNA在DN中表达失调,可能通过转化生长因子(TGF)-β1/NF-κB/信号转导及转录激活蛋白(STAT)3/糖原合成酶激酶(GSK)-3β信号通路参与DN的发展和进展,可能是疾病的早期诊断或预后的标志物或作为减缓DN进展的治疗靶点〔7,8〕。
lncRNA NKILA通过抑制NF-κB活动,使T细胞对活化诱导的细胞死亡(AICD)敏感而促进肿瘤免疫逃避〔9〕。NKILA在非小细胞肺癌(NSCLC)〔10〕、恶性黑色素瘤〔11〕、乳腺癌〔12〕和食管鳞状细胞癌(ESCC)〔13〕中均可通过阻断NF-κB信号通路抑制癌细胞的迁移、侵袭或上皮间质转化。目前针对NKILA的主要研究集中肿瘤等方面,还有研究表明,lncRNA NKILA对脓毒症肾损伤后自噬具有调节作用〔2〕。本研究说明抑制lncRNA NKILA对足细胞具有潜在的保护作用。
本研究通过StarBase预测显示,miR-22-3p与lncRNA NKILA存在结合位点,可能参与lncRNA NKILA对足细胞凋亡和增殖的调控。miR-22-3p在糖尿病心肌病(DCM)大鼠模型中表达下调,通过MIAT/miR-22-3p/DAPK2信号通路调控心肌细胞的凋亡〔10,14〕。且miR-22-3p在脓毒症急性肾损伤中表达下调〔4〕,在草酸钙诱导肾结石模型中参与肾小管上皮细胞的凋亡〔3〕。但其对DN足细胞中的表达和作用尚不清楚。本研究结果表明低表达miR-22-3p可部分逆转抑制lncRNA NKILA对高糖诱导的HPC增殖和凋亡的作用。
PI3K/Akt/NF-κB轴可能在足细胞损伤中起关键作用,PI3K/AKT途径的激活可阻止高糖诱导的足细胞凋亡〔15,16〕。本研究发现,抑制lncRNA NKILA表达可激活PI3K/AKT信号通路,下调miR-22-3p则逆转了抑制lncRNA NKILA对高糖诱导的HPC细胞PI3K/AKT信号通路的激活作用。说明lncRNA NKILA靶向miR-22-3p可能通过PI3K/AKT信号通路发挥对足细胞的保护作用。
综上,在高糖诱导的HPC细胞中,lncRNA NKILA通过调控miR-22-3p激活PI3K/AKT信号通路,抑制高糖诱导的足细胞凋亡,提高细胞存活率。lncRNA NKILA可能是DN潜在的靶点。
猜你喜欢
眼科新进展(2022年2期)2022-03-11
中国老年学杂志(2022年2期)2022-02-17
现代临床医学(2021年5期)2021-11-02
天津医科大学学报(2021年4期)2021-08-21
天津医科大学学报(2021年4期)2021-08-21
建材发展导向(2021年11期)2021-07-28
广东蚕业(2021年1期)2021-03-18
山东医药(2020年9期)2020-05-20
祝您健康(2020年5期)2020-05-14
当代水产(2020年10期)2020-03-17
