
HIF1α/HIF2α经EGF调控胶质瘤细胞化疗抵抗机制研究*
2022-05-07 11:10郭海燕
重庆医学 2022年8期
杜 军,汪 攀,龚 胜,赵 璐,郭海燕,廖 彬△
(1.重庆市人民医院神经外科 401147;2.重庆市黔江中心医院神经外科 409000)
胶质瘤作为颅内常见的肿瘤,预后差并且易复发,既往研究认为这与肿瘤组织的缺氧微环境关系密切[1-2]。肿瘤组织缺氧微环境主要由低氧诱导因子1α(hypoxia inducible facto-1α,HIF1α) 和低氧诱导因子2α(hypoxia inducible facto-2α,HIF2α)参与调控[3-5],随着肿瘤的进展,HIF2α对肿瘤生长的影响减弱,而HIF1α变得越来越重要[6-7]。因此针对HIF1α的药物开发成为研究热点之一,旨在抑制患者肿瘤的生长[8]。不幸的是,这种靶向治疗并不成功,它并不能显著抑制肿瘤体积,因此,本研究通过HIF1α和HIF2α单独敲除以及同时敲除来探究其原因。
表皮生长因子(Epidermal growth factor,EGF)在缺氧条件下高水平表达,并通过EGF受体(EGFR)和磷脂酰肌醇激酶/丝氨酸苏氨酸激酶(PI3K/AKT)信号通路调控胶质瘤生长[9-10]。有研究认为,缺氧微环境下肿瘤高表达EGF,其与EGFR结合,从而激活PI3K/AKT信号通路,诱导HIF1α稳定表达,促进肿瘤恶性进展[11-12]。然而,很少有研究HIF1α/HIF2α与EGF之间的相互关系,本研究旨在通过体外实验探讨HIF1α和HIF2α经EGF调控胶质瘤细胞化疗抵抗的机制。
1 材料与方法
1.1 材料
U87细胞和293T细胞购于中国科学院上海细胞库。DMEM高糖培养基、0.25%胰酶和胎牛血清购于美国Gibco公司。乳酸脱氢酶细胞毒性检测试剂盒和细胞凋亡检测试剂盒购于上海碧云天生物技术有限公司。替莫唑胺(temozolomide,TMZ)购于中国阿拉丁公司。鼠抗人β-actin单克隆抗体购自上海生工生物科技有限公司,鼠抗人HIF1α单克隆抗体和鼠抗人HIF2α单克隆抗体购于美国Abcam公司。人EGF细胞因子购于美国R&D Systems公司。LentiCRISPRv2、pMD2.G及psPAX2质粒购于美国Addgene公司。
1.2 细胞培养及HIF1α和HIF2α基因敲除
U87细胞采用含10%胎牛血清DMEM高糖培养基,于饱和湿度、5% CO2、95% N2、37 ℃条件下培养。缺氧培养条件为饱和湿度、1% O2、5% CO2、94% N2、37 ℃。在https://chopchop.cbu.uib.no/网站查得CRISPR/CAS9敲除靶序列,HIF1α的靶序列为:5′-GAA CTC ACA TTA TGT GGA AGT GG-3′,HIF2α的靶序列为:5′-CTT GGA GGG TTT CAT TGC CGT GG-3′。根据LentiCRISPRv2质粒说明书合成带BsmBⅠ酶切位点的靶序列DNA片段,进一步克隆至LentiCRISPRv2质粒中,对上述质粒测序并验证插入片段,测序结果正确后将其与pMD2.G和psPAX2质粒共转染至293T细胞行慢病毒包装。浓缩纯化的病毒感染细胞后添加嘌呤霉素予以药物抗性筛选。根据敲除情况分为HIF1α单独敲除组、HIF2α单独敲除组、HIF1α/HIF2α同时敲出组,另设未转染的U87细胞为对照组。
1.3 Western blot检测HIF1α和HIF2α表达
各组细胞常氧或缺氧培养24 h后,收集细胞提取总蛋白并用BCA法测浓度,十二烷基硫酸钠-聚丙烯酰胺凝胶(SDS-PAGE)电泳1 h,再冰浴电转2 h至聚偏氟乙烯(PVDF)膜。10%脱脂奶粉室温封闭2 h。一抗β-actin(1∶2 000)、HIF1α(1∶1 000)和HIF2α(1∶1 000)4 ℃孵育过夜,二抗(1∶5 000)室温孵育2 h,洗涤后化学发光拍照,分析蛋白条带灰度值,以β-actin为内参。
1.4 实时荧光定量PCR(RT-qPCR)检测EGF mRNA表达变化
各组细胞常氧或缺氧培养24 h后,收集细胞使用TRIzol提取总RNA,反转录为cDNA后,以cDNA为模板进行RT-PCR,以β-actin作为内参。引物序列为,EGF正向引物5′-GCC CCC CTG CCT CCT CCA AGT G-3′,反向引物5′-GGG GGT GGA GTA GAG TCA GGG CAA -3′;β-actin-正向引物5′-ACC CGC CGC CAG CTC ACC-3′,反向引物5′-GGG GGG CAC GAA GGC TCA TC-3′。反应条件:95 ℃,5 min;95 ℃ 30 s,60 ℃ 30 s,72 ℃ 30 s,40个循环:反应体系:2×SYBR Mix(10.0 μL)、正向引物(0.4 μL)、反向引物(0.4 μL)、cDNA(50 ng)、ddH2O补至20 μL。通过Bio-Rad CFX Manager Software软件分析得到EGF mRNA表达水平。
1.5 ELISA检测培养上清液中EGF水平变化
各组细胞常氧或缺氧培养72 h,收集上清液500×g离心5 min去掉细胞碎片。标准品及标本100 μL加入孔中,37 ℃孵育1 h;吸取上清液后加入100 μL检测溶液A,37 ℃孵育1 h;洗板3次后,加入100 μL检测溶液B,37 ℃孵育30 min;洗板3次后,加入90 μL 3,3′,5,5′-四甲基联苯胺(TMB)底物溶液,37 ℃避光显色10~20 min,加入50 μL终止液后用酶标仪检测450 nm处吸光度(A)值。根据标准品A值,建立浓度-A值标准曲线,计算出各组细胞培养上清液中EGF浓度。
1.6 细胞毒性检测
各组细胞按1×104/孔细胞接种96孔板,加入不同浓度TMZ(400、800 μmol/L)再常氧或缺氧培养72 h再进行乳酸脱氢酶(LDH)释放检测,96孔板400×g离心5 min,取120 μL上清液于另一96孔板,各孔加入60 μL LDH检测工作液,室温避光孵育30 min,酶标仪检测490 nm处A值。计算各组细胞毒性:细胞毒性率(%)=(处理样品A值-样品对照孔A值) /(细胞最大酶活性A值-样品对照孔A值)×100。细胞最大酶活性A值为TMZ药物处理的细胞孔经裂解后检测LDH的A值。
1.7 细胞凋亡率检测
各组细胞缺氧培养24 h后,添加TMZ(200 μmol/L)再缺氧培养72 h,通过流式细胞仪检测细胞凋亡。细胞经消化为单个细胞后取1×105个细胞重悬于100 μL预冷PBS中, 300×g离心5 min,弃上清液;加入195 μL结合缓冲液和5 μL Annexin V-FITC重悬细胞,室温避光孵育15 min,300×g离心5 min,弃上清液;加入190 μL结合缓冲液及10 μL碘化丙啶(PI)溶液重悬细胞,室温避光孵育10 min后冰上放置并进行流式检测,保存数据并进行统计分析。
1.8 统计学处理

2 结 果
2.1 Western blot验证U87细胞HIF1α、HIF2α敲除效果
缺氧条件下,HIF1α单独敲除组HIF1α表达水平降低,HIF2α单独敲除组HIF2α表达水平降低,HIF1α/HIF2α同时敲除组2个蛋白表达水平均降低,见图1。

A:对照组;B:HIF1α单独敲除组;C:HIF2α单独敲除组;D:HIF1α/HIF1α同时敲除组。
2.2 U87细胞HIF1α和HIF2α敲除后EGF表达检测
U87细胞EGF mRNA表达水平缺氧条件较常氧条件显著升高(P<0.05);缺氧培养24 h后,HIF1α、HIF2α单独敲除组及HIF1α/HIF2α同时敲除组EGF mRNA表达水平低于对照组(P<0.05),且HIF1α/HIF2α同时敲除组也低于HIF1α和HIF2α单独敲除组(P<0.05);缺氧培养72 h后,HIF1α、HIF2α单独敲除及HIF1α/HIF2α同时敲除组细胞培养上清液中EGF浓度低于对照组(P<0.05),同时HIF1α/HIF2α同时敲除组EGF浓度也低于HIF1α和HIF2α单独敲除组(P<0.05)。见图2。
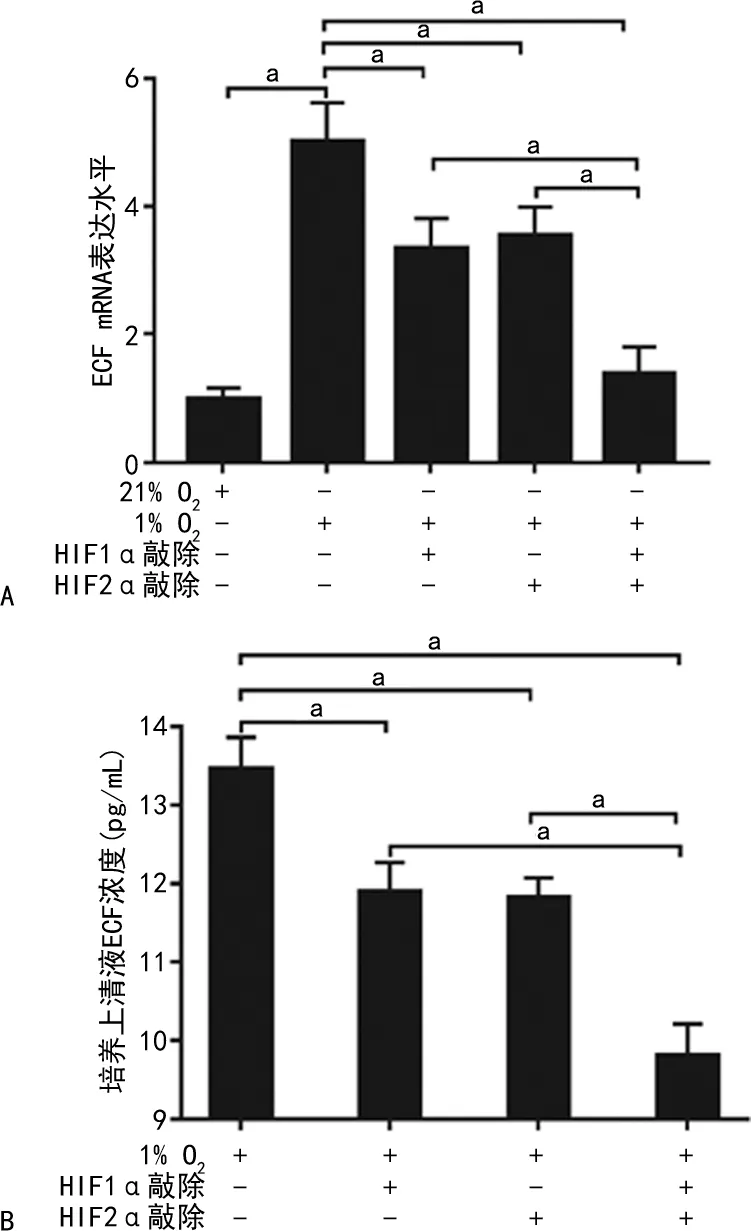
A:PCR检测EGF mRNA结果分析;B:ELISA检测培养上清液中EGF浓度结果分析;a:P<0.05。
2.3 U87细胞HIF1α和HIF2α敲除后TMZ对其细胞毒性的检测
TMZ(400、800 μmol/L)处理对U87细胞毒性率缺氧培养比常氧培养明显降低(P<0.05);缺氧培养72 h,HIF1α、HIF2α单独敲除组细胞毒性率较对照组差异无统计学意义(P>0.05),HIF1α/HIF2α同时敲除细胞毒性率较对照组及HIF1α、HIF2α单独敲除组升高(P<0.05)。TMZ(200 μmol/L)处理并缺氧培养72 h,HIF1α、HIF2α单独敲除组及HIF1α/HIF2α同时敲除细胞凋亡率均高于对照组(P<0.05),且HIF1α/HIF2α同时敲除组细胞凋亡率高于HIF1α、HIF2α单独敲除组(P<0.05)。见图3。
2.4 EGF对HIF1α和HIF2α同时敲除组细胞毒性率和凋亡率的影响
在TMZ(400、800 μmol/L)处理并缺氧培养72 h的HIF1α/HIF2α同时敲除组细胞加入EGF(20 ng/mL),细胞的细胞毒性率和细胞凋亡率(TMZ 200 μmol/L)明显低于未加入EGF的细胞(P<0.05)。见图4。

A~D:细胞凋亡检测图;E:细胞凋亡检测统计分析结果;F:细胞毒性检测统计分析结果;a:P<0.05。

a:P<0.05;A、B:细胞凋亡检测图;C:细胞凋亡检测统计分析结果;D:细胞毒性检测统计分析结果。
3 讨 论
由于肿瘤组织血管无序化和肿瘤细胞高代谢,所以胶质瘤细胞始终处于一个缺氧的微环境中[13]。已有研究证实,缺氧微环境可促进肿瘤细胞增殖及维持胶质瘤干细胞干性从而抑制凋亡及产生化疗抵抗[14]。而在缺氧微环境中,HIF1α和HIF2α起主要调控作用[5]。因此,抑制HIF1α或HIF2α可抑制肿瘤的发生发展及复发。例如,抑制HIF1α表达可增加胶质瘤细胞对化疗药物的敏感性[7],而靶向胶质瘤干细胞中的HIF2α可降低肿瘤的复发[15]。因此,这些研究强烈支持开发HIF抑制剂治疗胶质瘤,因为在理论上可以抑制肿瘤的进展。一些HIF抑制剂已经被批准进入了临床试验阶段。然而,截至目前,还没有HIF靶向治疗成功治愈胶质瘤的报道。本研究在对HIF1α和HIF2α单独敲除之后又进行了同时敲除,均检测了在缺氧条件下TMZ处理对细胞毒性和凋亡的影响。结果证实单独敲除HIF1α和HIF2α缺氧条件下TMZ处理后的细胞毒性率与对照组比较差异无统计学意义(P>0.05),而同时敲除组较两单独敲除组和对照组细胞毒性率和细胞凋亡率均显著增高。笔者认为HIF1α和HIF2α均参与调控胶质瘤细胞化疗抵抗。结合课题组前期研究,缺氧条件下HIF1α敲除之后HIF2α蛋白表达水平升高,同时HIF2α敲除之后HIF1α蛋白表达水平升高[16],所以单独敲除其中一个蛋白后另一个蛋白表达升高弥补了敲除蛋白的作用继续产生化疗抵抗。
在胶质瘤中已有报道EGF是HIF1α的上游蛋白[17]。而在本研究中,敲除HIF1α和HIF2α之后,EGF表达水平下降。同时HIF1α和HIF2α同时敲除细胞培养基中添加EGF因子,缺氧条件下TMZ对细胞的毒性和细胞凋亡均降低。HIF1α和HIF2α具有高度的同源性,并具有一些相同的靶基因,这表明它们可能与相同的HRE序列结合。HIF1α和HIF2α都含有一个保守的DNA结合区域,称为bHLH-PAS结构域,其与靶基因启动子里面HREs结合后促进该基因表达[18]。因此,笔者前期研究中,已验证了在缺氧胶质瘤细胞中HIF1α和HIF2α可通过结合EGF启动子上HRE序列AGGCGTGG和GGGCGTGG影响EGF启动子活性。因此,HIF1α和HIF2α与EGF之间存在反馈调控。EGF促进HIF1α的表达,相反,HIF1α和HIF2α的上调促进EGF的表达。总之,HIF1α和HIF2α可通过EGF调控胶质瘤的发生发展和复发,这为胶质瘤的治疗提供了新的靶点和策略。
猜你喜欢
临床神经外科杂志(2022年6期)2022-11-14
中国现代医生(2022年19期)2022-08-25
临床医学工程(2022年7期)2022-08-01
新疆农业科学(2022年6期)2022-07-13
中国农学通报(2022年12期)2022-06-01
临床外科杂志(2022年2期)2022-03-23
作物学报(2022年4期)2022-02-10
中国药学药品知识仓库(2021年11期)2021-11-25
中小学德育(2020年11期)2020-03-18
中学生物学(2019年7期)2019-10-17
