
雪茄烟Beinhart 1000-1赤星病抗性的主基因+多基因混合遗传分析
2022-04-25 02:50:30童治军卢桂萍陈学军曾建敏方敦煌
西南农业学报 2022年3期
童治军,卢桂萍,陈学军,曾建敏,方敦煌
(1.云南省烟草农业科学研究院/烟草行业烟草生物技术育种重点实验室/国家烟草基因工程研究中心,昆明 650021;2.云南省烟草公司昆明市公司,昆明 650051)
【研究意义】烟草赤星病是由链格孢菌[Alternariaalternata(Frires) Keissler]引起的真菌性病害,主要发生在烟草叶片上,特别是生理成熟的叶片[1-2],各烟草种植区均有分布,是威胁世界烟草生产的毁灭性病害[3-5]。在中国各大烟区,烟草赤星病对产量、质量影响较大,一般年份发病率为20%~30%,严重的发病率达90%,可减少产值达50%以上,损失达9205.433万元[6-8]。【前人研究进展】烟草赤星病的发生流行与主栽烟草品种的抗病性密切相关,目前生产上以化学防治为主、生物防治为辅,但收效甚微[8-10]。培育抗烟草赤星病的优良品种仍是预防该病最经济、有效的途径之一,而其核心是深入明确抗赤星病烟草品种的遗传规律。研究表明,烟草赤星病的抗性是属于受多基因控制的数量性状。郭永峰等[11]对不同烟草品种赤星病抗源的抗性比较分析后,得出Beinhart 1000-1与净叶黄对烟草赤星病的抗性是不同源的,且Beinhart 1000-1的抗病性是由显性多基因控制,而净叶黄的抗性基因大部分是隐性的。通过进一步采用双列杂交分析其抗性遗传因素可知,Beinhart 1000-1与净叶黄的赤星病抗性遗传均符合加性—显性模型[12]。与之类似,冯莹等[13]利用高抗赤星病烟草品种净叶黄、Beinhart 1000-1和感病品种NC82为材料分别构建了2个杂交组合的P1、P2、F1、F2世代群体,并采用主基因+多基因混合遗传模型进行抗性遗传分析,结果表明,两者的赤星病抗性均受2对加性—完全显性主基因+加性—显性多基因混合遗传模型(2 MX-CE-AD)控制。但Ramm等[1]对Beinhart 1000-1赤星病抗性遗传研究结果表明Beinhart 1000-1赤星病抗性是显性单基因控制的典型质量性状。由于针对雪茄烟品种Beinhart 1000-1赤星病抗性遗传规律的研究结果存在较大差异,极不利于利用该抗源培育抗性品种,故此有待进一步对其进行遗传分析研究和挖掘。【本研究切入点】本研究以抗病雪茄烟品种Beinhart 1000-1为父本、以感病烤烟品种红花大金元为母本杂交、自交构建的341个株系重组自交系群体F7为研究材料,试验主基因+多基因混合遗传模型方法评价亲本P1、P2和F7世代群体材料的赤星病抗性基因遗传规律,确定其遗传模型、估算遗传率。【拟解决的关键问题】以期为下一步开展赤星病抗性基因QTL定位,开发与之紧密连锁的分子标记进行分子标记辅助选择进而培育烟草抗赤星病新品种提供理论基础。
1 材料与方法
1.1 材料
以抗赤星病的雪茄烟品种Beinhart 1000-1(B1000-1)为父本(P1)、以感病烤烟品种红花大金元(HD)为母本(P2),通过杂交、自交及单粒传法构建了341个株系的烟草重组自交系群体F7。2020年将双亲、重组自交系群体及抗、感病对照品种净叶黄、G140种植于云南省玉溪市研和试验基地大田中,采用完全随机区组设计,每个株系种植4个重复,每个重复种植10株。
1.2 烟草赤星病抗性表型测定
按人工病圃轻简化鉴定烟草赤星病抗性的方法[14]选择病圃进行田间布局、控肥栽培、较低浓度大面喷雾接种形成人工病圃、保湿诱发赤星病,接种后第7、14、21天进行3次调查,统计病情指数[15](DI:Disease Index)。以每次调查的平均病情指数和病害流行曲线下面积(AUDPC:Area Under Disease Progress Curve)[16]作为烟草赤星病抗性表型测定数据(即接种后第7、14、21天的平均DI值和AUDPC值4类数据,分别记为T1、T2、T3和T4)。
1.3 数据统计分析
采用文献[17]的方法进行。
2 结果与分析
2.1 抗性表型数据分析
亲本红花大金元和Beinhart 1000-1的赤星病病情指数(DI)均呈现出显著(P<0.05)或极显著差异(P<0.01)。同样,烟草重组自交系群体F7中的DI值分别出现了远低于抗病亲本Beinhart 1000-1和远高于感病亲本红花大金元,表现出超亲遗传。烟草重组自交系群体F7的DI值变异系数(CV)存在丰富的变异(318.42%~100 396.70%),适合遗传分析(表1)。此外,除T1外,烟草重组自交系群体F7DI值的偏斜度和峰度绝对值均大于1,但小于1.5,概率密度函数回归曲线表明该重组自交系群体的赤星病抗性表型呈连续性的正态分布或偏正态分布(图1),属于典型的数量性状遗传。
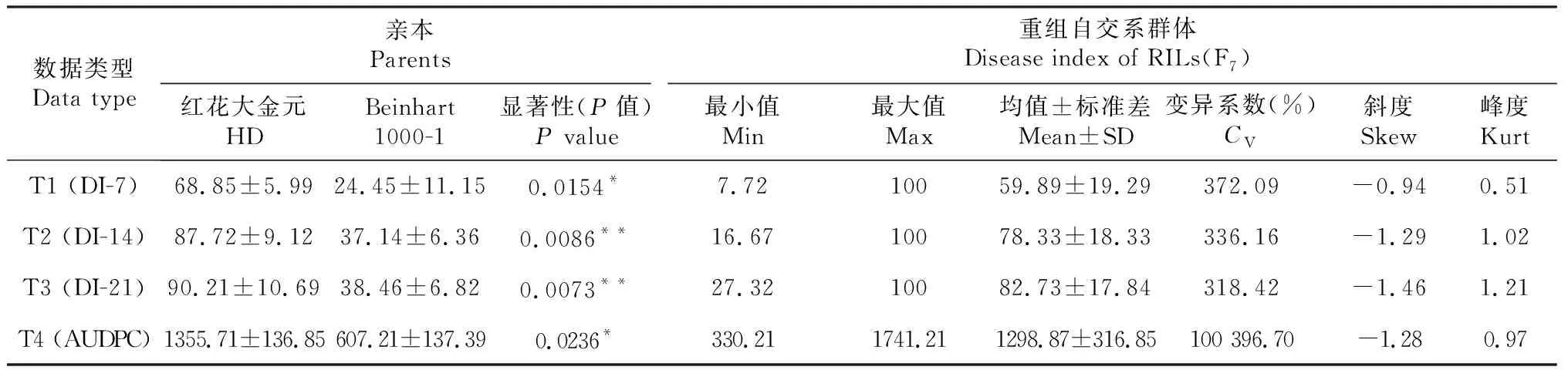
表1 亲本及重组自交系群体的赤星病抗性表型统计
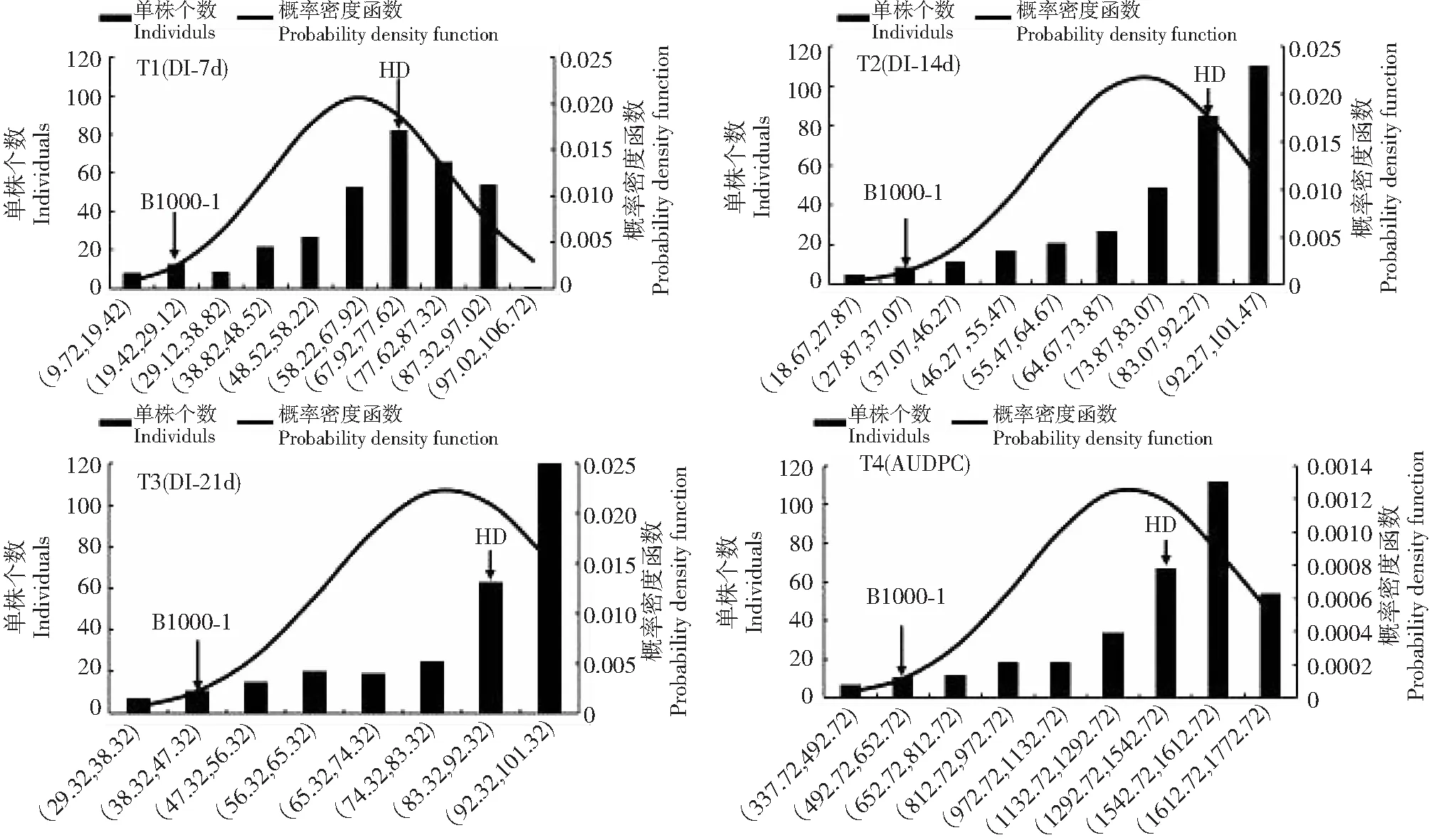
图1 3个不同调查时期烟草重组自交系群体的赤星病抗性表型分析Fig.1 Phenotype statistic values of resistance brown spot of tobacco RILs in 3 different disease investigation periods
2.2 候选模型的选择及适合性检测
对3个不同调查时期各世代(P1、P2和F7)的赤星病抗性表型分别进行主基因+多基因混合遗传模型分析,根据AIC值最小原则初选模型,选取AIC值最小的3组遗传模型候选(表2),对3个不同调查时期进行适合性检验(U12、U22、U32、nW2和Dn),以AIC值最小或相对小且统计量达到显著性水平(P<0.05)个数最少的模型作为最优遗传模型(表3)。结果表明T3(即接种21 d后)模型是烟草赤星病抗性性状的最优遗传模型,由4对加性—上位性主基因遗传控制的4 MG-AI模型。
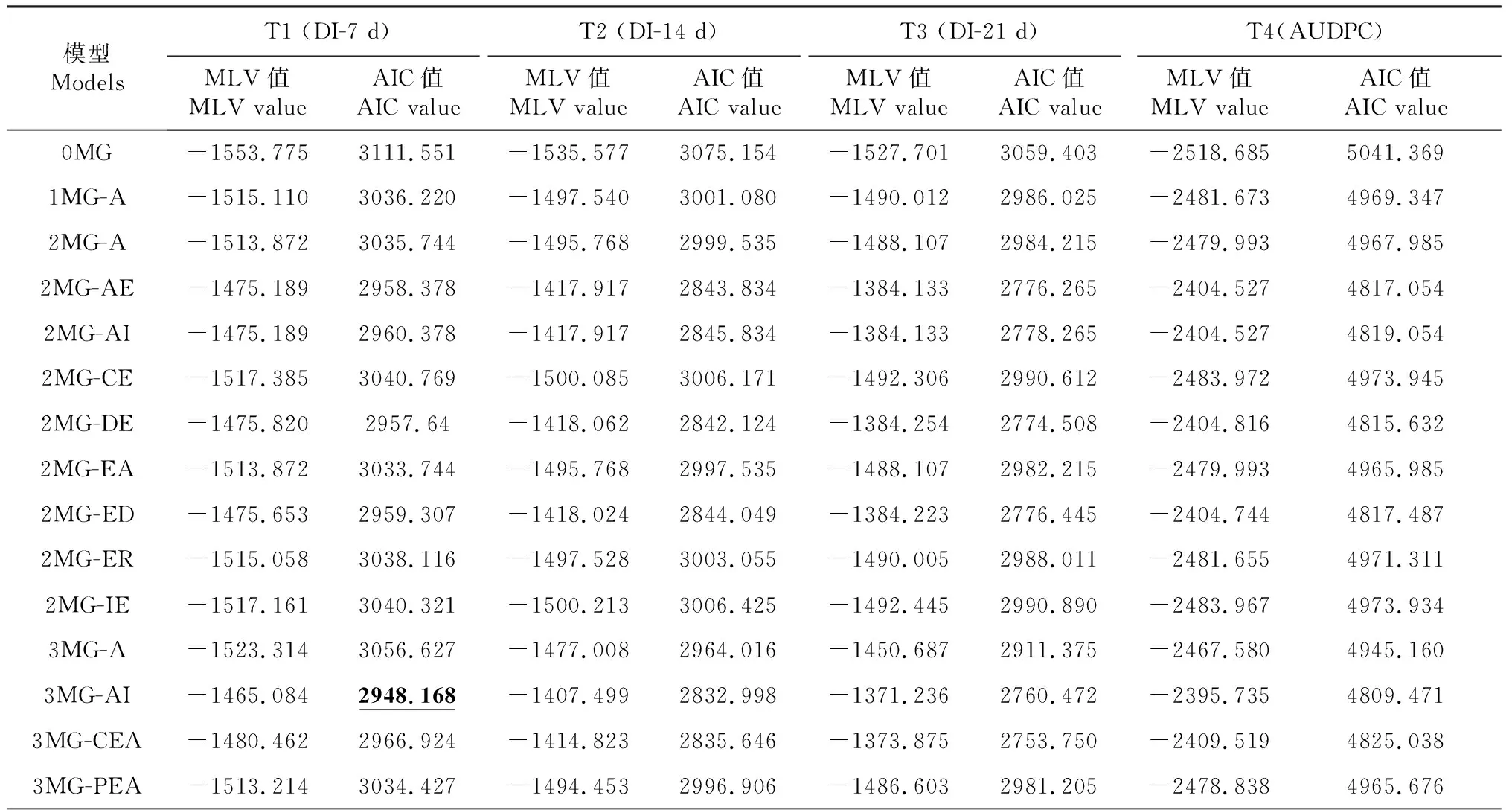
表2 3个不同调查时期各遗传模型的AIC值和极大似然函数MLV值
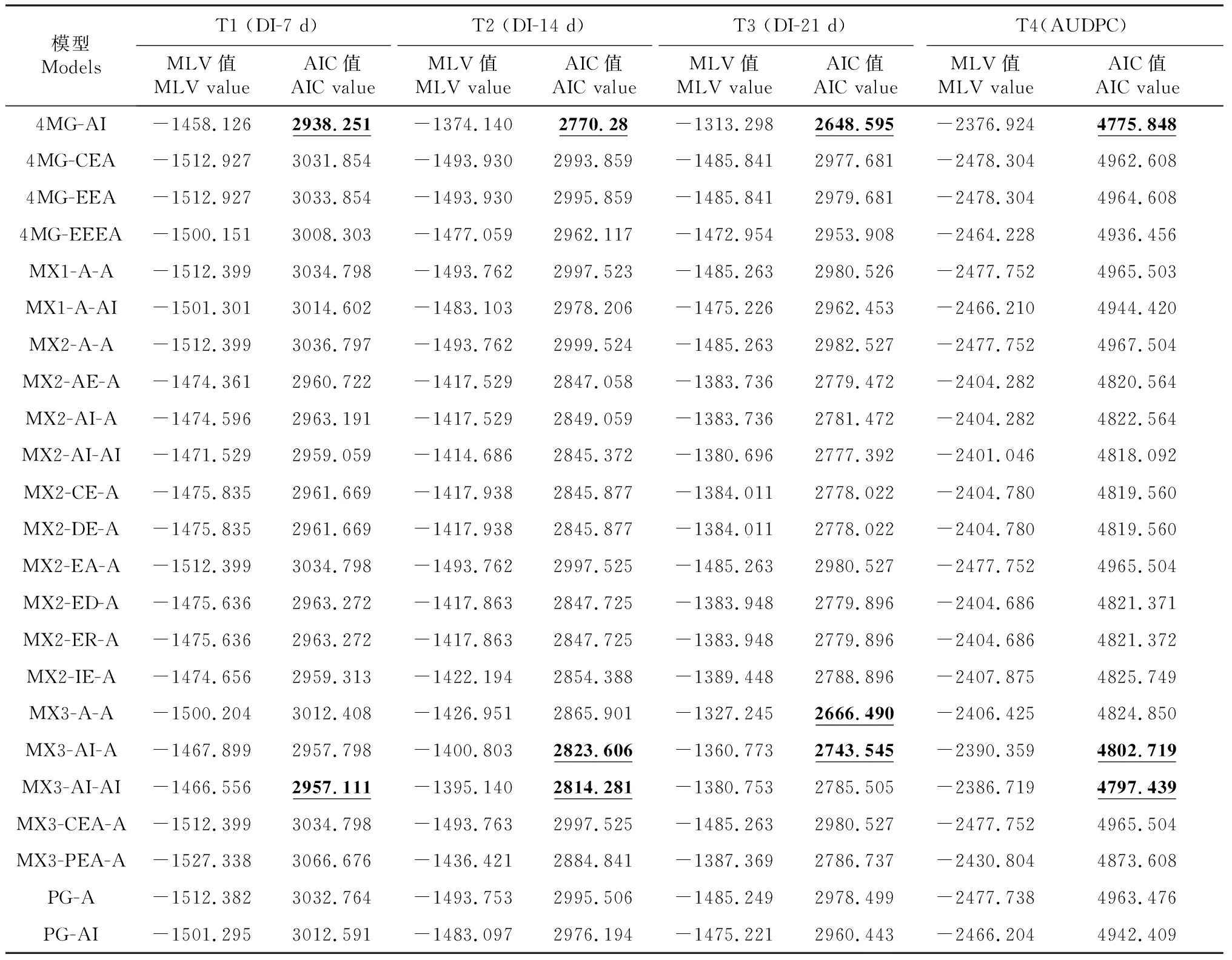
续表2 Continued table 2
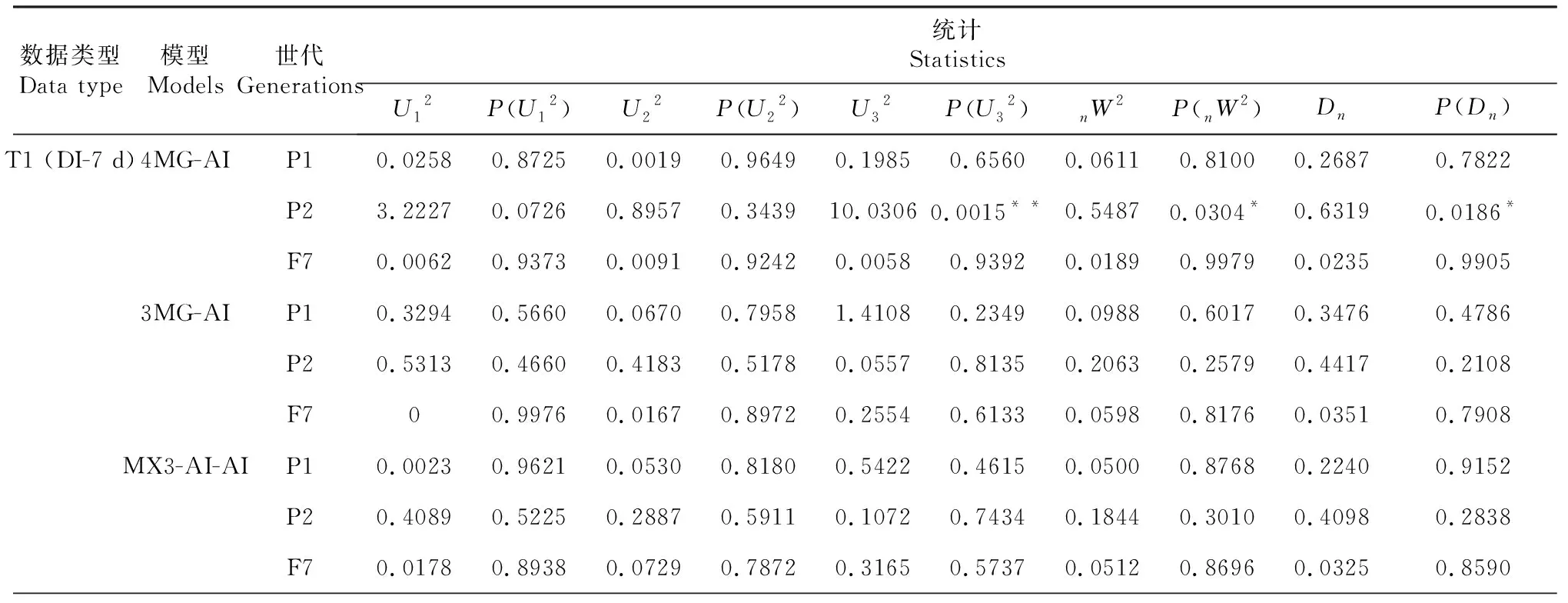
表3 候选模型的适合性检验
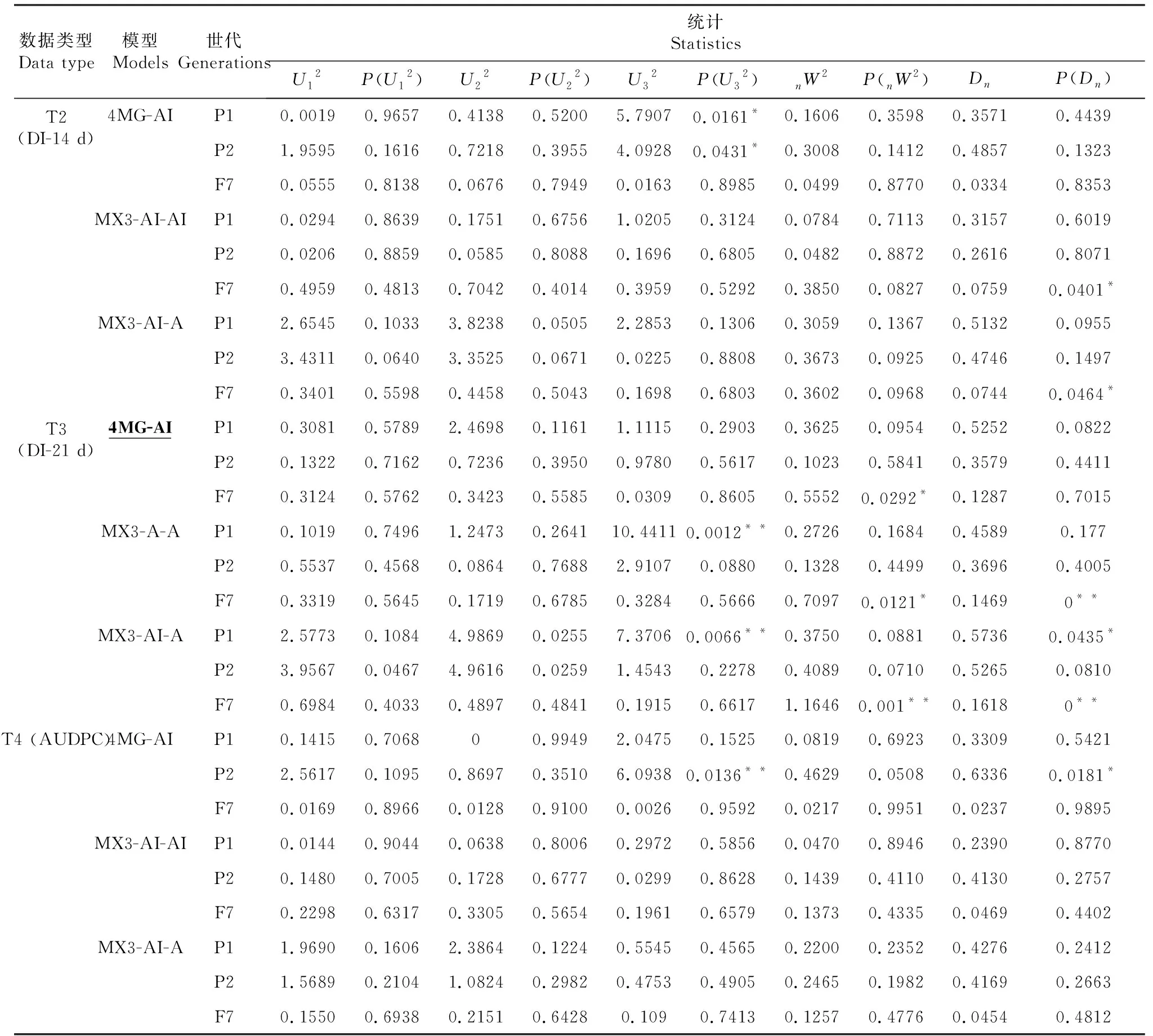
续表3 Continued table 3
2.3 最优模型的遗传参数估计
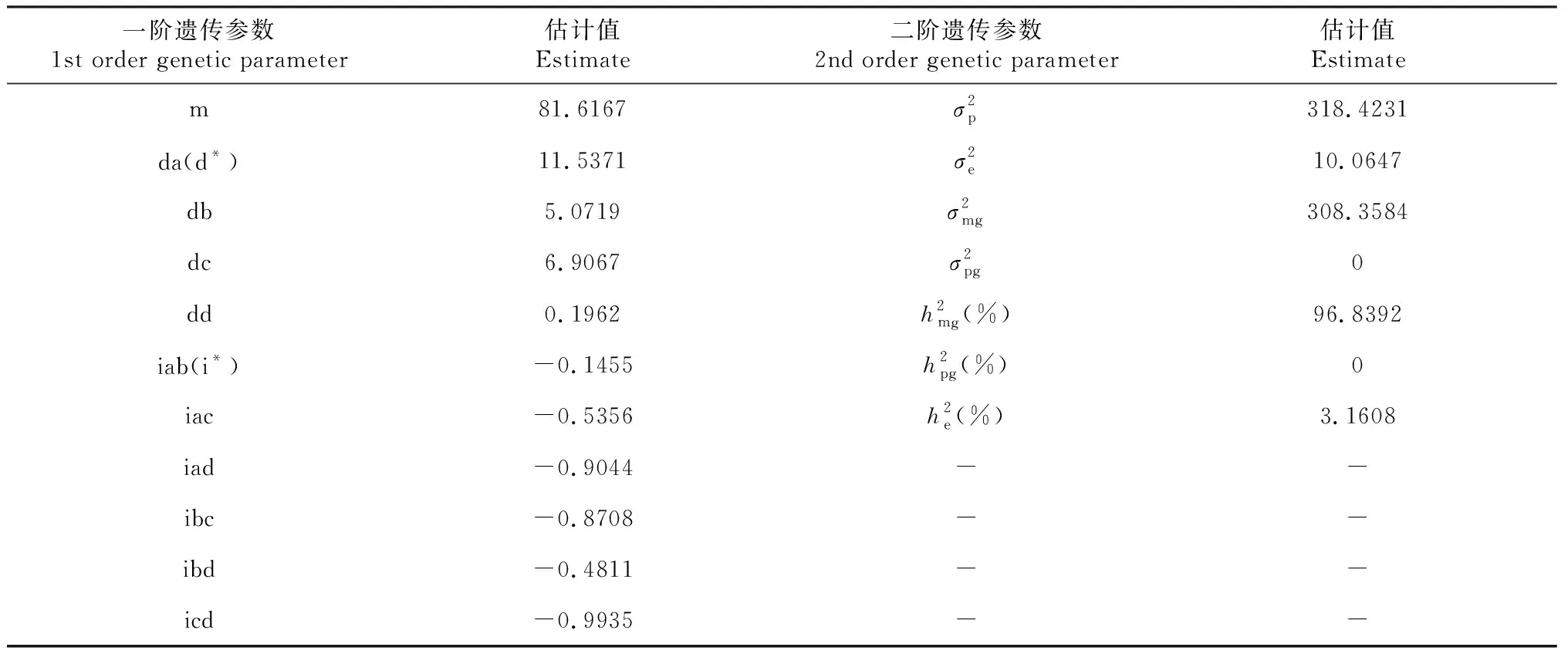
表4 最适遗传模型4MG-AI的烟草赤星病抗性遗传参数估计值
3 讨 论
目前已报道的烟草赤星病抗源主要有3种[1,18],即雪茄烟品种Beinhart 1000-1、烤烟品种净叶黄和野生烟草N.suaveolens,其中Beinhart 1000-1和净叶黄是国内外烟叶生产上主要利用的赤星病抗源。针对抗源净叶黄,大量的研究表明其赤星病抗性由显性主基因+多基因共同控制,主基因的显性效应和加性效应在遗传中起主导作用,有研究者基于上述遗传分析结果并结合不同遗传分离群体对源自净叶黄的赤星病抗性基因进行了QTL定位分析[19-21]。针对雪茄烟品种Beinhart 1000-1的赤星病抗性遗传分析研究表明,其抗性性状是受微效多基因控制和环境影响的数量性状,而Ramm等[1]的研究结果却表明Beinhart 1000-1的抗性受1对显性单基因控制,属于典型的质量性状。究其原因,Ramm等[1]虽采用多个感病烤烟品种作为亲本与Beinhart 1000-1进行组合配制,但其在早期的研究中仅使用多个组合的杂交子一代(F1)并简单统计各F1组合烟株的赤星病发病情况,缺乏进一步利用遗传分离群体的遗传分析。此外,郭永峰等[11-12]通过双列杂交分析Beinhart 1000-1与感病品种NC82配制杂交组合的赤星病抗性,结果表明,Beinhart 1000-1的赤星病抗性遗传符合主基因与多基因加性—显性模型。基于相同亲本组合,冯莹等[13]利用P1、P2、F1和F2世代群体,采用主基因+多基因混合遗传模型对Beinhart 1000-1进行赤星病抗性遗传分析,结果表明,其赤星病抗性受两对加性—完全显性主基因+加性—显性多基因混合遗传模型(2MX-CE-AD)控制,且多基因的显性效应大于加性效应,即Beinhart 1000-1的赤星病抗性遗传以主基因效应为主,但受环境影响较大。与之类似,高婷婷等[22]基于Beinhart 1000-1与G140构建F2遗传群体的研究结果显示,Beinhart 1000-1的赤星病抗性在田间表现为多个主基因控制。童治军等[23]利用Beinhart 1000-1与HD构建的重组自交系群体,对Beinhart 1000-1赤星病抗性的遗传分析结果也表明,Beinhart 1000-1的抗性受主基因控制,其主基因的加性效应起主导作用,环境效应的影响较小。本研究将抗赤星病的雪茄烟品种Beinhart 1000-1与感病烤烟品种红花大金元组合P1、P2、F7世代3个不同调查时期赤星病抗性表型调查结果采用主基因+多基因混合遗传模型进行了多世代联合分析,较全面分析了Beinhart 1000-1的赤星病抗性遗传效应,结果表明Beinhart 1000-1的赤星病抗性受4对加性—上位性主基因控制,为4 MG-AI遗传模型,4对主基因加性效应远大于他们间的互作效应,表明主基因加性遗传的贡献率起主导作用;主基因的遗传效应高达96.8392%,无多基因遗传效应,而环境效应仅为3.1608%,即环境因素的影响可以忽略。该结果与高婷婷等[22]和童治军等[23]对Beinhart 1000-1抗性遗传分析的结果相似,但却与郭永峰等[11-12]和冯莹等[13]的研究结果不尽相同,主要原因是采用的遗传分析群体不同,永久性分离群体如重组自交系高世代群体的表型数据利用的是单株系群体,而暂时性分离群体如F2表型是单株,理论及实践上均证明利用群体比单株获得的试验结果更准确、更可靠。
4 结 论
采用植物数量性状“主基因+多基因”混合遗传模型联合分析方法,分析了雪茄烟品种Beinhart 1000-1的赤星病抗性遗传规律,明确了Beinhart 1000-1赤星病抗性受主基因控制,且主基因的加性效应起主导作用,环境效应影响小。因此,在烟草抗病育种过程中要重视利用主基因,应在早期世代对赤星病抗性性状进行选择。
猜你喜欢
安徽农业科学(2022年6期)2022-04-11 21:49:49
广西植物(2021年1期)2021-03-24 18:30:12
北京农学院学报(2019年1期)2019-02-22 05:39:46
山西农经(2018年17期)2018-04-03 11:30:26
消费导刊(2017年24期)2018-01-31 01:29:31
农学学报(2017年12期)2018-01-04 12:37:48
辽宁大学学报(哲学社会科学版)(2017年3期)2017-06-21 21:16:59
河南农业科学(2017年4期)2017-04-12 09:55:45
西南农业学报(2016年5期)2016-05-17 05:42:20
西南农业学报(2016年6期)2016-04-16 05:12:47
