
益生菌Nissle 1917鞭毛蛋白高变区随机缺失及鞭毛展呈外源抗原的构建
2022-04-25 02:50:48金正雨邓永军区炳明
西南农业学报 2022年3期
杨 颖,金正雨,邓永军,区炳明,文 明
(1.贵州大学动物科学学院,贵阳 550025;2.贵州省动物生物制品工程技术研究中心,贵阳 550025;3.肇庆学院生命科学学院,广东 肇庆 526061)
【研究意义】大肠杆菌Nissle 1917(EscherichiacoliNissle 1917,EcN)作为益生菌使用已具有100多年的历史[1],其不含肠毒素、溶血素、细胞毒素等致病因子,无致突变、遗传毒性及DNA损伤活性[2],能合成细菌素和具有六铁摄取系统以利于该菌株在肠道中与其他菌株竞争[3],此外,EcN可以利用复杂的机制靶向定殖肿瘤部位进而用于肿瘤靶向治疗[4]。因为EcN的益生特性,其一直是德国等国家医用药品(Mutaflor®)的活性成分[5],主要用于治疗人类消化道菌群失调的相关疾病[6-7]。随着分子生物学和全基因组测序技术的发展,现已完整注释EcN的基因组序列[8],基于此,更多EcN基因重组的研究构建出具有新性质的EcN衍生物,这将促进其在未来成为生物活性分子载体或用于口服活疫苗开发的载体系统[9-10]。【前人研究进展】细菌鞭毛表面展呈技术是将外源抗原融合到鞭毛蛋白高变区,随后构建的嵌合鞭毛组装成鞭毛丝,进而实现细菌对外源抗原的展呈[11-12]。由于细菌鞭毛丝由成千上万个鞭毛蛋白单体组装而成[13],把外源抗原嵌合到鞭毛蛋白高变区,有利于外源抗原的大量表达,并且能利用鞭毛蛋白的佐剂效应[14-15],促进机体对外源抗原的捕获、加工和提呈。【本研究切入点】EcN具有发达的鞭毛系统,前期研究发现其鞭毛蛋白可以容许10个氨基酸的缺失和6个氨基酸外源抗原的插入[16]。【拟解决的关键问题】为寻找外源抗原的其他可能插入位点,并尝试插入更大片段的外源抗原,选用EcN无质粒菌株(EcN cured of its two cryptic plasmids pMUT1 and pMUT2,EcNc),构建EcNc鞭毛蛋白高变区随机缺失质粒库,筛选可缺失位点和插入外源抗原进行展呈,为EcN靶向投递外源抗原提供操作策略。
1 材料与方法
1.1 材料
1.1.1 菌株、质粒和细胞 供试菌株、质粒和细胞的特征及来源见表1。

表1 供试菌株、质粒和细胞
1.1.2 主要试剂和培养基 限制性内切酶、T4DNA连接酶、Marker购自TaKaRa公司;TransStart FastPfu DNA Polymerase购自北京全式金生物技术有限公司;MutExpress TM ⅡFast Mutagenesis Kit、DNA凝胶回收试剂盒购自南京诺唯赞生物科技有限公司;BAL-31核酸酶购自NEB(北京)有限公司;胰蛋白胨(Tryptone)、酵母提取物(Yeast extract)均购自Oxoid公司;RPMI1640-F12(1∶1)细胞培养基、新生牛血清(NCS)均购自Gibco(美国)公司。
1.2 方法
1.2.1 EcNc fliC高变区随机缺失质粒库的构建SpeⅠ为fliC基因中部高变区内唯一的酶切位点,选择SpeⅠ单酶切pBR322-fliC重组质粒,回收单一线性片段后,用Nuclease BAL-31经不同时间(6、7、10、20 min)消化黏性末端,回收各时间段消化产物,用DNA聚合酶Ⅰ(Klenow)大片段补齐黏性末端后,经T4DNA ligase环化连接,转化DH5α化学感受态,37 ℃培养1 h后加入10 mL LB(Amp)培养过夜,提取质粒进行琼脂糖凝胶电泳检测。
1.2.2 fliC高变区缺失区域的筛选 将fliC高变区随机缺失质粒库提取质粒,电转化EcNc ΔfliC感受态后,涂布LB(Amp)固体培养基,37 ℃培养过夜,挑取单菌落接种半固体运动性平板[17],检测各菌株运动性能,并设计引物P1: 5′-ATGGCACAAGT CATTAATACCAACA,P2: TTAACCCTGCAGCAGAGACA GA-3′对相应fliC基因进行测序,分析缺失区域。
1.2.3 EcNc FliC高变区展呈外源抗原的构建 设计引物P3: 5′-GCTGGGCAAACTGCTAGT-3′,P4:5′-TGCTACATCACCTGCTTTTGCT-3′,以pUC18-fliC质粒为模板,进行反向PCR扩增缺失fliC基因缺失910~927处的碱基序列;设计引物P5:5′-CAAAAGCAGGTGATGTAGCAAATGCATCTTATGCCGGTGT-3′,P6: 5′-CCACTAGCAGTTTGCCCAGCTACTCTTTGAA TCTGTCCGA-3′, 以F18菌株基因组为模板扩增其FedF基因178~327位碱基序列;上述2种扩增产物回收后混合,在Exnase®催化下,37 ℃反应30 min完成重组反应,实现2个线性化DNA的体外环化,构建pUC18-fliCF(fliCF片段为fliC基因910~927位插入F18FedF基因178~327位碱基序列的重组片段),再将fliCF片段连接到自杀载体pRE112,构建重组自杀质粒pRE112-fliCF并转化EcNc,参照文献[18]方法进行重组菌株EcNcfliCF的筛选,并用引物P1、P2进行fliCF序列扩增和测序鉴定。
1.2.4 透射电镜和运动性观察 将EcNc与EcNcfliCF过夜培养的菌液分别1∶100转接于新鲜的LB中,37 ℃震摇培养至OD600nm约为1.0,各细菌均调整为1.0,参照文献[18]的方法进行电镜观察和运动能力检测。
1.2.5 IPEC-J2 细胞黏附试验 调整对数生长期的细菌浓度至1×106CFU,以10∶1的比例加入96孔板中的单层IPEC-J2 细胞,在细胞培养箱中共孵育1 h,用PBS洗涤孵育孔3次,去除未黏附的细菌,然后加入0.5%的Triton X-100裂解液,37 ℃作用20 min裂解细胞获得其表面黏附的细菌,梯度稀释涂布LB平板,培养过夜后计算黏附细菌数目,利用Graphpad软件作图并分析差异显著性。
2 结果与分析
2.1 fliC高变区随机缺失质粒库的构建
提取质粒进行琼脂糖凝胶电泳,检测结果(图1)表明,Nuclease BAL-31经不同时间(6、7、10、20 min)消化EcNcfliC高变区、再经补齐黏性末端环化转化DH5α化学感受态后,质粒提取物在琼脂糖凝胶电泳中呈现弥散条带,表明EcNcfliC高变区出现不同程度的缺失,获得EcNcfliC高变区随机缺失质粒库。

M: λ-EcoT14 I digest; 1~4: 经Nuclease BAL-31分别消化6、7、10、20 min构建的质粒库; 5: pBR322-fliC; 6: pBR322M: λ-EcoT14 I digest; 1-4: The plasmids library is constructed by digestion of Nuclease BAL-31 for 6, 7, 10 and 20 minutes respectively; 5: pBR322-fliC; 6: pBR322图1 EcNc fliC高变区随机缺失质粒库鉴定结果Fig.1 Identification of random deletion plasmid library in hypervariable region of EcNc fliC
2.2 fliC基因高变区缺失区域的筛选
从图2看出,EcNcfliC高变区随机缺失质粒库电转化EcNc ΔfliC感受态后,挑取过夜培养的单菌落接种半固体运动性平板,运动性检测显示,大部分菌株未呈运动性(图2-A,2-B);扩增相应fliC序列测序结果显示,大部分高变区随机缺失的区域为非3的整数倍缺失(图3);试验筛选到一株fliC高变区缺失菌株具有运动性(图2-C),经测序其fliC基因缺失910~927 bp处碱基序列(图3)。
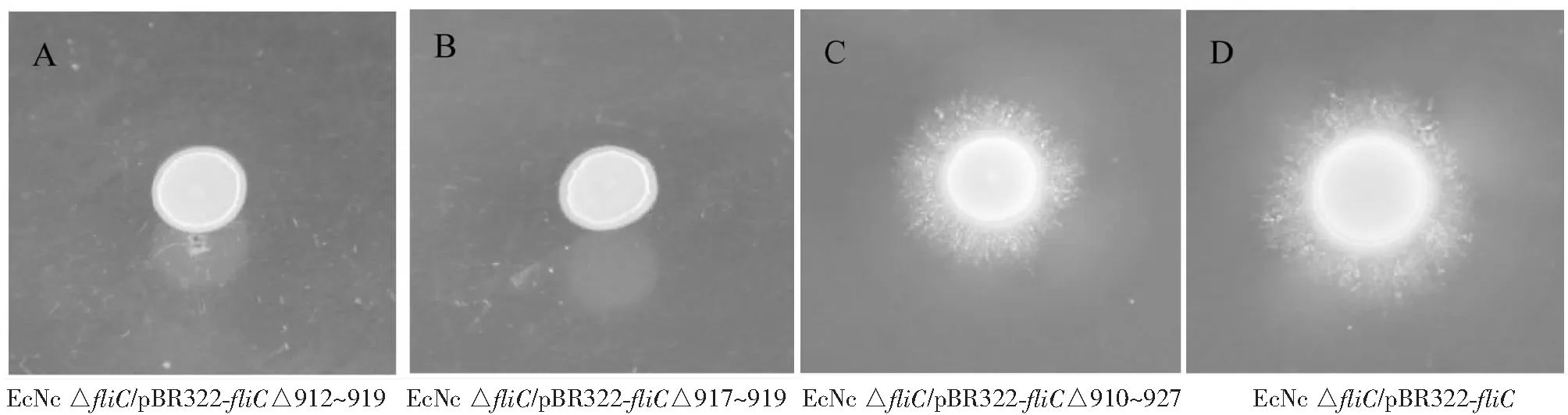
图2 部分菌株运动能力检测结果Fig.2 Motility detection of some EcNc strains

图3 部分EcNc fliC高变区缺失区域Fig.3 Hypervariable region deleted area of fliC of some EcNc strains
2.3 重组菌株EcNcfliCF的构建
利用 MutExpress TM ⅡFast Mutagenesis Kit,构建fliC基因910~927位嵌合F18 FedF基因178~327位碱基序列的重组片段fliCF,将fliCF连接入自杀载体pRE112中转化到EcNc,2次重组后经PCR和测序鉴定,结果显示,重组菌EcNcfliCF构建正确。
2.4 重组菌株EcNcfliCF表型的检测
从图4可知,EcNcfliCF重组菌株在电镜下能观察到鞭毛丝,但在运动平板上无运动性能。
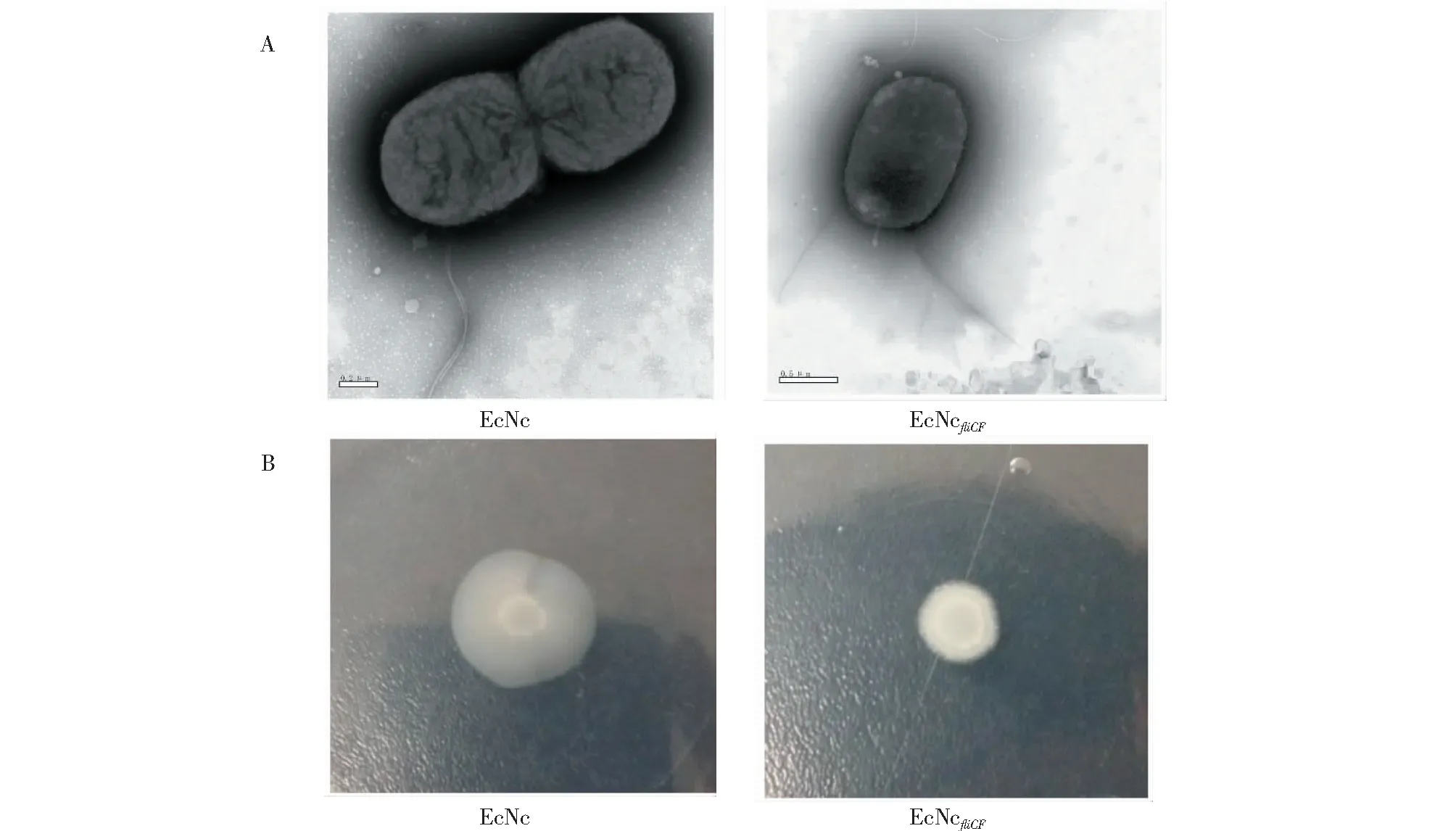
图4 重组菌株EcNc和EcNcfliCF的表型Fig.4 Phenotypes of EcNc and EcNcfliCF
2.5 IPEC-J2细胞黏附检测
从图5可知,相对EcNc,EcNcfliCF对 IPEC-J2细胞的黏附能力减少约0.28%,差异不显著(P>0.05)。
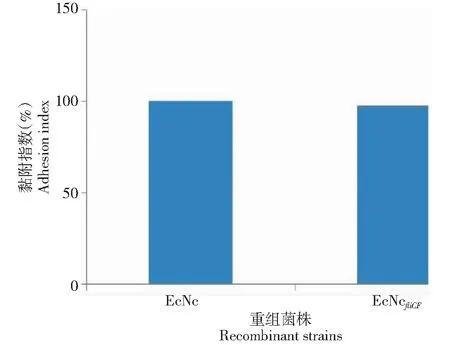
图5 重组菌株对IPEC-J2的黏附指数(EcNc的黏附指数假设为100%)Fig.5 Bacterial adherence to IPEC-J2 cells (EcNc strain adhesion index is assumed as 100%)
3 讨 论
益生菌通过不同机制预防和控制胃肠道致病菌感染,改善动物生产性能和产量,作为抗生素促进剂的替代物越来越受到关注。自Alfred Nissle发现摄入特定的大肠杆菌菌株可以治疗感染性疾病患者起,EcN菌株作为益生菌被应用于防治人类疾病。杨颖等[19-22]从猪粪便中分离到EcN,其能持续在猪体内定殖,该菌株可以保护猪宿主免受产毒素大肠杆菌的侵害,EcN对机体的保护作用有赖于其F1C菌毛和鞭毛介导的对有害菌株的黏附抑制[22]。
EcN鞭毛丝含有大量鞭毛蛋白单体,在前期研究中发现可利用鞭毛蛋白展呈技术将外源抗原展呈于EcN表面,重组EcN菌株定殖于机体发挥益生作用的同时,还可诱导机体产生针对外源抗原的体液及黏膜免疫应答[23]。但是不同血清型的细菌,其鞭毛蛋白尤其是高变区氨基酸差异极大,使用鞭毛蛋白展呈技术需要确定较好的外源抗原插入位点,进而优化外源抗原的展呈效果。故本研究基于鞭毛蛋白构建重组质粒,产生高变区随机缺失的质粒库,筛选鞭毛蛋白可缺失的区域(fliC基因缺失910~927 bp处碱基序列),该区域缺失后,并不影响菌株鞭毛丝的形成和运动能力。EcN含有2种隐性质粒,分别命名为pMUT1和pMUT2,前期研究中,已构建无质粒的EcN变体(EcNc),其生物学特征与野生型EcN菌株没有区别,可以更好地用作基因工程活载体菌[24]。
EcNc已被证明可以通过其鞭毛系统展呈6His标签[16]。EcNc鞭毛蛋白高变区较长,位于195~320 aa处。本研究基于筛选到的EcNc鞭毛蛋白可缺失区域(910~927 bp处碱基序列),在该缺失区域处插入F18大肠杆菌黏附素FedF亚单位与仔猪小肠上皮细胞相互作用的受体结合域(60~109位氨基酸残基区段),成功将FedF亚单位受体结合域的50个氨基酸展呈于EcNc表面。构建的重组菌株EcNcfliCF能够形成鞭毛丝,虽然不具备运动能力,但不影响其对IPEC-J2细胞的黏附能力;且在EcNc鞭毛蛋白高变区插入50个氨基酸,与6His标签的长度比较有了显著的容纳性,大大提升插入容量。
4 结 论
研究成功将FedF亚单位受体结合域的50个氨基酸展呈于EcNc表面,这为后期进一步筛选鞭毛蛋白高变区其他缺失区域,和基于已筛选位点插入展呈其他外源抗原,优化构建遗传稳定、定植力强的功能性益生菌打下基础。
猜你喜欢
舰船科学技术(2022年11期)2022-07-15 07:51:56
西藏农业科技(2019年3期)2019-11-04 00:35:10
实用检验医师杂志(2019年3期)2019-11-01 04:21:48
人人健康(2019年10期)2019-10-14 03:25:12
现代园艺(2018年3期)2018-02-10 05:18:12
科学家(2017年20期)2017-11-10 23:10:19
上海农业学报(2017年3期)2017-04-10 12:39:12
科技资讯(2016年19期)2016-11-15 10:39:37
西南医科大学学报(2015年1期)2015-08-22 13:01:46
医学研究杂志(2015年6期)2015-07-01 17:41:11
