
芥蓝SPL基因家族鉴定及表达分析
2022-04-18 08:23曾东琳唐为玲曾家晶GefuWANGPRUSKI赖钟雄郭容芳
西北植物学报 2022年3期
曾东琳,唐为玲,曾家晶,Gefu WANG-PRUSKI,赖钟雄,郭容芳,*
(1 福建农林大学园艺植物生物工程研究所,福州 350002;2 福建农林大学戴尔豪西大学联合实验室,福州 350002)
Squamosa-promoter binding protein-like(SPL)蛋白构成了一个转录因子的多样化家族,在植物生长和发育中起着重要作用。该蛋白存在由76个氨基酸组成的高度保守区域即SBP结构域[1]。SBP结构域存在2个锌指结构,由8个半胱氨酸(Cys)或组氨酸(His)残基组成,其中前4个氨基酸残基结合一个锌离子,后4个氨基酸残基结合另外一个锌离子,锌离子对于维持其构象稳定具有重要作用[2]。此外,SBP结构域还参与核导入和序列特异性DNA结合,该序列特异性DNA可以结合到含有GTAC核心基序和基因特异性侧翼区的共有结合位点[3]。研究表明SPL基因在调控植物根系发育[4]、叶片形成[5]、花器官发育[6]、果实成熟[7]、胚胎发育[8]和发育阶段转变[9]等方面发挥着重要作用。
在植物中,转录因子能与某些特定基因启动子区域的顺式作用元件特异性结合,激活或抑制基因转录,进而调控下游基因的表达[10]。前人发现SPL转录因子在植物生长发育、信号转导及响应生物和非生物胁迫等方面具有重要作用[11-12]。在拟南芥中,由于miR156结合位点的突变而阻碍了其对AtSPL13的负调控而显著延迟了叶原基的出现,从而AtSPL13在萌发期间延迟了真叶的生长[3]。过量表达SPL3会导致拟南芥叶片起始速率轻微下降[13]。过量表达SPL10会降低拟南芥叶片生长速率,并且过早地经历营养期变化[14-16]。SPL2可与AS2结合进而调控拟南芥花器官发育[6]。SPL9通过直接激活2个MYB基因来调节拟南芥毛发分布[15]。此外,SPL转录因子还参与植物信号传导,例如光信号传导[16]、赤霉素信号传导[17-18]和温度信号[19-20]等。综上所述,SPL在调节植物叶片发育、植株生长和信号传导等方面发挥着重要作用。
芥蓝(Brassicaoleraceavar.alboglabra) 是一种十字花科蔬菜,其类型繁多,形态各异。芥蓝含有较多的水分、抗坏血酸、矿物质和芥子油苷[21],在甘蓝类蔬菜中有着较高的营养价值,其叶片、花茎到花薹均可食用[22]。其中‘改良香菇’芥蓝生长势强,叶面有花状变形叶,产量高,商品性状好,风味口感酥脆[23]。因此‘改良香菇’芥蓝的叶片是其重要的植物学性状,可以作为杂交种试配的依据,也可以作为指示性状。本研究以‘改良香菇’芥蓝为试验材料,对芥蓝SPL基因(BoSPL)进行全基因组鉴定及分析,并对芥蓝不同生长时期及不同组织部位进行定量分析,克隆BoSPL3-1、BoSPL10-2和BoSPL11-2基因,为进一步揭示‘改良香菇’芥蓝叶片发育过程的分子机制,利用基因编辑技术调控芥蓝叶片发育,丰富芥蓝种质资源,培育芥蓝新品种奠定了基础。
1 材料和方法
1.1 实验材料
‘改良香菇’芥蓝由广东揭阳农友种子有限公司提供,种植于福建农林大学温室大棚前地块。挑选长势良好的‘改良香菇’植株,将其分为抽薹未开花时期(Ⅰ)和全部结荚期(Ⅱ)2个时期,在Ⅰ时期对植株的根、真叶、薹叶和花茎进行取样;在Ⅱ时期对植株的根、真叶、薹叶、花茎、处于子叶形胚时期的种子和种荚进行取样。为了进一步鉴定调控芥蓝花状变形叶发育过程中的基因,种植‘改良香菇’芥蓝于福建农林大学园艺楼培养室内。培养至30 d时,挑选长势良好的‘改良香菇’植株,取其叶片的4个部位:叶缘、叶肉、叶脉和花状变形叶,每份样品0.1 g,保存于-80 ℃冰箱备用。
1.2 方 法
1.2.1 总RNA提取以及cDNA合成选用TaKaRa试剂盒RNAiso Plus(Code No.9109)提取芥蓝样品RNA;使用超微量分光光度计测定RNA浓度及吸光值(OD值),测定结果OD500值在1.8~2.0 之间为最佳。使用PrimeScriptRTreagent Kit with gDNA Eraser (Code No. RR047A)试剂盒进行反转录成cDNA。
1.2.2 芥蓝SPL基因家族全基因组鉴定通过网站(https://plants.ensembl.org/)下载甘蓝基因组序列,从拟南芥基因组数据库(http://arabidopsis.org/)获得SPL基因家族ID号,利用拟南芥基因组中已公布的SPL氨基酸序列作为种子序列,与芥蓝参考基因组数据库进行同源比对获得同源基因。再将同源基因与NCBI数据库中的SWISS-MODEL进行蛋白质序列比对,采用SMART(http://smart.embl-heidelberg.de/)在线分析蛋白结构域,进一步分析以确定该家族中的成员个数。运用在线网站对BoSPL进行蛋白理化性质(https//web.Expasy.org/protparam/)、信号肽(http//www.cbs.dtu.dk/services/SignalP-4.0/)和亚细胞定位(http//cello.life.nctu.edu.tw/)分析。运用NCBI(https://www.ncbi.nlm.nih.gov/Structure/bwrpb/bwrpsb.cgi)在线网站分析BoSPL基因结构。使用MEME在线软件(http://meme-suite.org/tools/meme)分析该家族成员编码蛋白的保守基序。使用TBtools绘制基因结构和保守基序图。将预测出来的BoSPL家族的蛋白质序列,结合由TAIR官网(https://www.arabidopsis.org/)上下载的拟南芥SPL蛋白质序列,运用MAGA-X进行多重序列比对,使用最大似然法(maximum likelihood)构建系统进化树,自展法系数(Bootstrap)设置1 000次重复来检验分支点的稳定性。
1.2.3 基因克隆根据‘改良香菇’不同时期组织部位表达模式结合文献中与叶片发育相关的SPL基因,选择BoSPL3-1[13]、BoSPL10-2[13-14, 24]和BoSPL11-2[24]进行基因克隆,GeneBnak上获得的登录号分别为OK050589、OK050588和OK050590。并设计PCR扩增引物(表1)。以‘改良香菇’叶片cDNA为模板,进行PCR扩增。PCR反应体系如下:98 ℃预变性3 min;98 ℃变性10 s,60 ℃退火20 s,72 ℃延伸30 s,共35个循环;72 ℃延伸5 min。PCR扩增产物使用Thermo science Gene JET Plasmid Extraction Kit(Code No.00967821)试剂盒进行胶产物回收,回收产物-20 ℃冰箱保存。
1.2.4BoSPL3-1、BoSPL10-2和BoSPL11-2亚细胞定位使用Snapgene3.2.1设计引物(表1),去除终止密码子并分别在5′端加上NcoⅠ 和SpeⅠ酶切位点,将上述目的片段与pCAMBIA1302载体连接后转DH5α大肠杆菌感受态细胞,平板培养后挑单克隆菌落,以pCAMBIA1302-35S-GFP质粒上功能元件35S、GFP设计上下游引物pCAMBIA1302(表1)进行PCR筛选鉴定,鉴定成功的重组质粒通过冻融法转至农杆菌GV3101。最后通过农杆菌侵染法对烟草下表皮细胞进行瞬时表达,以空载pCAMBIA1302-35S-GFP为对照,培养48 h后在激光共聚焦显微镜下观察BoSPL3-1、BoSPL10-2和BoSPL11-2的定位情况。

表1 引物序列
1.2.5BoSPL基因荧光定量表达使用TB Green®Premix Ex TaqTM Ⅱ (Code No. RR820A)进行qRT-PCR反应,以BoActin为内参基因,使用CFX96 Real-Time PCR Detection System进行定量检测;利用Microsoft 2007软件整理试验数据,结果为3次生物学重复的平均值±SD,采用SPSS 24软件中的Duncan法进行显著性分析(P<0.05)。利用Primer Preminer5软件进行qRT-PCR引物设计(表1)。
2 结果与分析
2.1 BoSPL基因家族全基因组鉴定
利用生物信息学方法共获得芥蓝31个BoSPL家族成员,根据拟南芥已有的命名方式,依次命名为BoSPL1-1~BoSPL16-2(表2)。蛋白质特性分析发现,该家族成员编码蛋白质的氨基酸长度从157~1 028 aa不等;分子量介于18 041.04~113 747.21 Da之间,其中BoSPL1-1、BoSPL7-2、BoSPL10-1等电点小于6.55,属于酸性蛋白;其余BoSPL家族成员等电点均大于6.66,属于碱性蛋白;BoSPL蛋白质的氨基酸均没有信号肽,属于非分泌蛋白;亚细胞定位预测结果显示,所有的BoSPL家族成员均有定位在细胞核上,少数成员有定位在液泡、叶绿体和细胞质中。

表2 芥蓝SPL基因家族成员信息
使用MEME对BoSPL家族蛋白序列进行保守基序分析(图1),设定搜索出10个motif,分别命名为motif 1—10,motif 1、2、3高度保守,31个BoSPL成员中均含有该基序,推测其为SBP保守结构域。结合进化关系与保守基序图可以看出进化关系相似的BoSPL家族成员其保守基序分布相似。其中有4个成员包含motif 9,6个成员包含有motif 4、7、10,7个成员包含motif 8,13个成员中包含motif 6,20个成员包motif 5。
基因结构分析发现(图1),鉴定到的31个BoSPL均有内含子,其数量1~9个不等;外显子数量2~10个不等。其中,BoSPL3-1、BoSPL3-2、BoSPL5-1、BoSPL5-2、BoSPL5-3、BoSPL5-4、BoSPL5-5和BoSPL6-1均包含2个外显子,占总数的25.8%;BoSPL6-2、BoSPL8-1、BoSPL8-2、BoSPL9-1、BoSPL9-2、BoSPL13A-1、BoSPL13A-2、BoSPL13A-3、BoSPL15-1和BoSPL15-2均包含3个外显子,占总数32.3%;BoSPL2-2、BoSPL10-1、BoSPL10-2、BoSPL10-3、BoSPL11-1和BoSPL11-2均包含4个外显子,占总数的19.4%;BoSPL2-1拥有5个外显子占总数的3%;BoSPL1-1、BoSPL7-1、BoSPL7-2、BoSPL14-1、BoSPL16-1和BoSPL16-2均包含10个外显子占总数19.4%。结合基因结构和系统进化关系得出结构相似基因的内含子数量大多一致,亲缘关系也越近。
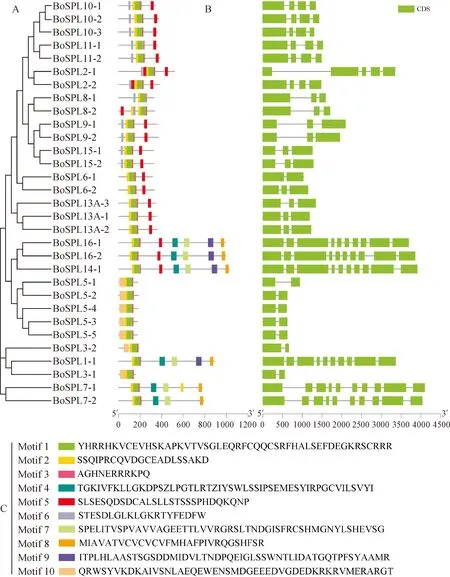
图1 芥蓝SPL基因家族成员蛋白保守基序和基因结构分布图
使用芥蓝和拟南芥SPL蛋白序列构建一个极大似然系统发生树(图2)。拟南芥中有16个SPL转录因子,根据其SBP保守结构域的氨基酸序列可分为8个进化支:AtSPL1/12/14/16、AtSPL2/10/11、AtSPL3/4/5、AtSPL6、AtSPL7、AtSPL8、AtSPL9/15和AtSPL13[25]。根据拟南芥中SPL分类结合进化关系,将芥蓝SPL分为Ⅰ~Ⅷ共8个亚族,各族成员分别包含4、2、3、2、7、4、2和7个成员。从系统进化树中可以看出BoSPL的进化关系与AtSPL相似。
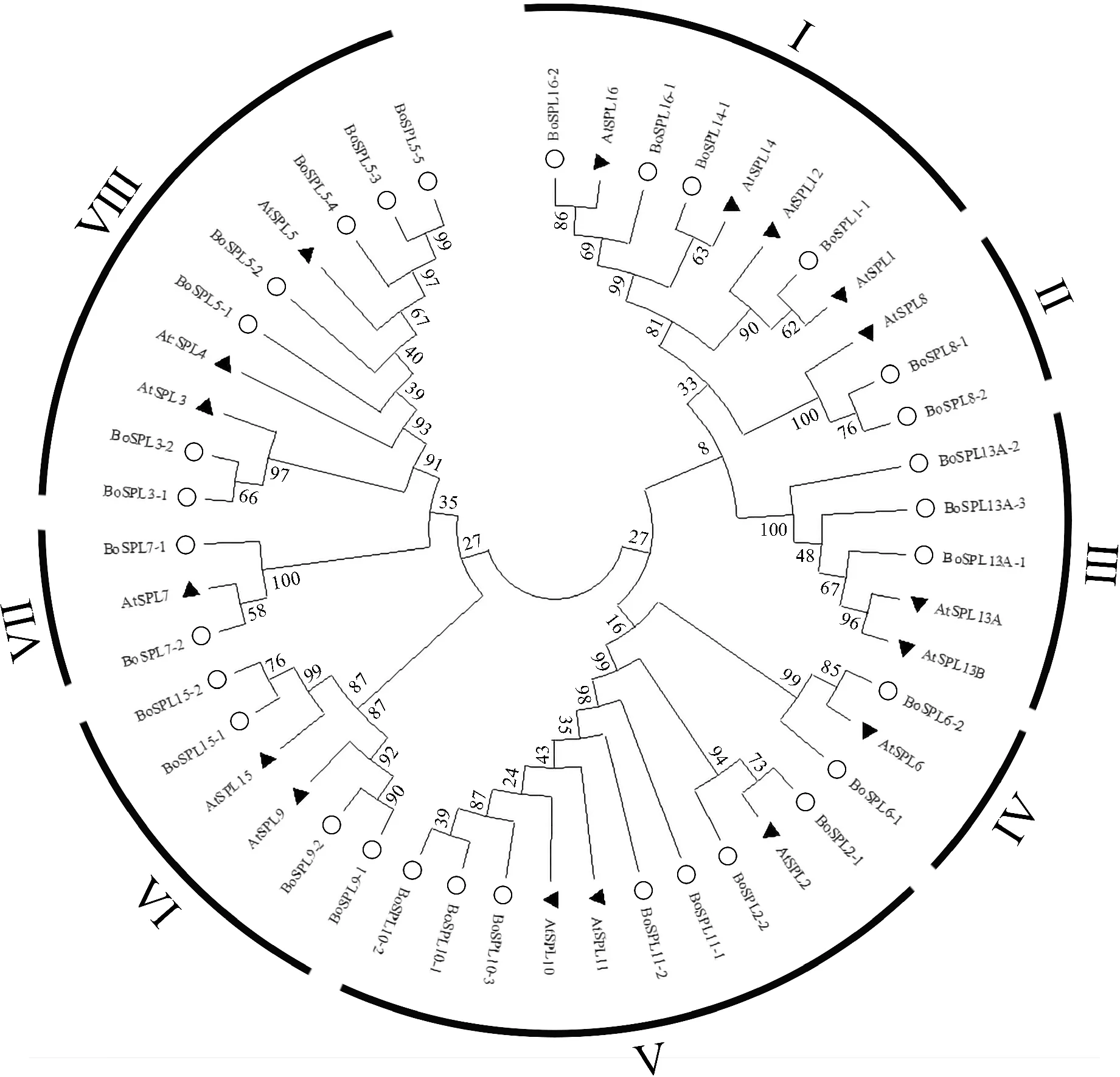
○.芥蓝;▲.拟南芥
2.2 BoSPL3-1、BoSPL10-2和BoSPL11-2基因克隆及重组质粒构建
PCR扩增凝胶电泳结果显示,图3,A-C分别在500 bp、1 200 bp和1 200 bp左右有一条清晰的条带,其大小均与预期一致,表明BoSPL3-1、BoSPL10-2和BoSPL11-2基因克隆成功;图3,C-E分别在700 bp、1 400 bp和1 400 bp左右有一条清晰的条带,表明pCAMBIA1302-35S-BoSPL3-1-GFP、pCAMBIA1302-35S-BoSPL10-2-GFP和pCAMBIA1302-35S-BoSPL11-2-GFP重组质粒构建成功,可用于亚细胞定位。
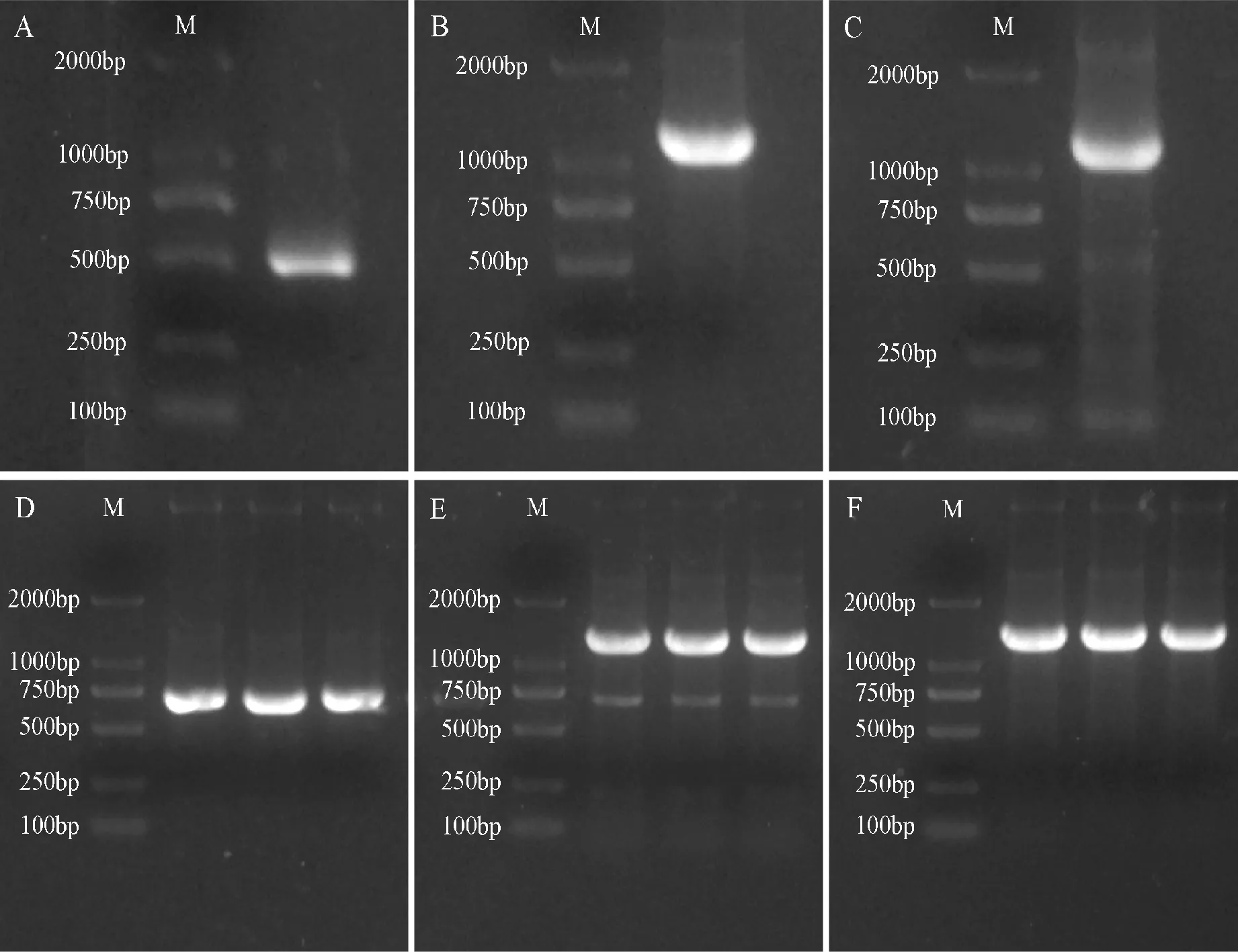
M. DL2000;A-C.BoSPL3-1、BoSPL10-2和BoSPL11-2基因的克隆; D-F.pCAMBIA1302-35S-BoSPL3-1-GFP、pCAMBIA1302-35S-BoSPL10-2-GFP和pCAMBIA1302-35S-BoSPL11-2-GFP重组质粒
2.3 BoSPL3-1、BoSPL10-2和BoSPL11-2亚细胞定位分析
经鉴定正确的重组质粒pCAMBIA1302-35S-BoSPL3-1-GFP、pCAMBIA1302-35S-BoSPL10-2-GFP和pCAMBIA1302-35S-BoSPL11-2-GFP转化到根癌农杆菌GV3101,将转化成功的含有亚细胞定位载体的农杆菌用于瞬时转化烟草下表皮细胞,在激光共聚焦显微镜下检测荧光信号。实验结果如图4所示,BoSPL3-1、BoSPL10-2和BoSPL11-2亚细胞定位均在细胞核中,该结果与预测结果一致。

图4 BoSPL3-1、BoSPL10-2和BoSPL11-2蛋白在烟草叶片的亚细胞定位
2.4 BoSPL家族基因在不同生长时期不同组织部位的qRT-PCR分析
根据系统进化关系结合陈文文等[26]不同亚族SPL基因功能不同,综合选取BoSPL1-1、BoSPL3-1、BoSPL10-2、BoSPL11-2、BoSPL14-1和BoSPL16-1设计引物做qRT-PCR分析,探讨其在‘改良香菇’芥蓝中不同生长时期不同组织部位表达模式(图5)。
结果显示,BoSPL3-1在种子中表达量较低,BoSPL1-1、BoSPL10-2、BoSPL14-1和BoSPL16-1在种子中的表达量较其他部位相比均较高,推测BoSPL1-1、BoSPL10-2、BoSPL14-1和BoSPL16-1可能与胚胎发育相关;其中在Ⅰ时期中叶片的表达量普遍比Ⅱ时期高;推测BoSPL可能与叶片发育早期的关系更为密切。BoSPL10-2在Ⅰ时期中的真叶和薹叶中相对表达量较高,推测其可能与叶片发育具有相关性,这与前人研究一致[11];BoSPL11-2在Ⅰ时期中根与真叶的表达量最高,推测其可能与植物的根、真叶发育相关性较大。
2.5 BoSPL3-1、BoSPL10-2和BoSPL11-2在叶片不同部位的qRT-PCR分析
根据‘改良香菇’芥蓝不同时期不同组织部位的表达模式(图5),结合Shikata等[24]过表达的SPL10/11具有宽的茎生叶,并且茎生叶和茎上有许多毛状体,且SPL10转基因植物的形状还会受到影响。综合分析选取BoSPL3-1、BoSPL10-2和BoSPL11-2设计引物做qRT-PCR分析其在‘改良香菇’叶片不同部位的表达模式(图6)。

R. 根;E. 真叶;SL. 薹叶;FM. 花茎;PD和SD为子叶胚时期的种荚、种子;Ⅰ. 抽薹未开花时期;Ⅱ. 完全结荚期;小写字母表示同一基因间0.05水平显著性差异,下同

A. ‘改良香菇’芥蓝叶片模式图;B. BoSPL3-1、BoSPL10-2和BoSPL11-2在‘改良香菇’芥蓝叶片表达模式:ME. 叶肉;LM. 叶缘;LV. 主叶脉;FL.花状变形叶
结果显示,BoSPL3-1、BoSPL10-2和BoSPL11-2在叶肉中相对表达量无显著差异。3个BoSPL基因中BoSPL10-2在花状变形叶中的表达量最高,表明BoSPL10-2可能与花状变形叶形成相关机制有关联,其作用机制如何有待进一步研究。
3 讨 论
SPL基因存在于所有绿色植物中,包括苔藓、裸子植物和被子植物[27-28]。本研究共鉴定到31个BoSPL成员,成员数量多于拟南芥(16个)[11]、水稻(19个)[29]和番茄(15个)[30]。鉴定得到BoSPL家族成员内含子数量在1~9个之间,进化关系相似,其基因结构相似性高。所有家族成员中均包含motif 1、motif 2、motif 3推测其构成SBP保守结构域,结构域的存在可能涉及到其进化关系以及行使基因功能的重要性。SBP结构域参与核导入和序列特异性的脱氧核糖核酸结合,该脱氧核糖核酸结合到含有GTAC核心基序和基因特异性侧翼区的共有结合位点[3]。依据进化关系结合拟南芥中的分类可以将BoSPL划分为8个亚族[25],且系统进化树表明BoSPL进化关系与拟南芥相似。值得注意的是,本研究中未鉴定到BoSPL4和BoSPL12,这可能是由于甘蓝中不存在这两个基因,其结果有待进一步验证。
亚细胞定位预测发现BoSPL3-1、BoSPL10-2和BoSPL11-2定位于细胞核中,与预测结果相一致(表2),表明其可能在细胞核内发挥调控作用。为进一步确定与叶片发育相关的BoSPL基因,本研究对6个BoSPL家族成员,检测不同时期不同组织部位的表达模式,结果表明各成员在检测组织中均有表达,但表达量存在明显差异,BoSPL1-1、BoSPL10-2、BoSPL16-1在种子中表达量较高,这与先前文章报道SPL与胚胎发育相关具有一致性[8]。此外,BoSPL3-1在Ⅰ时期的花茎和薹叶中的相对表达量较高,表明其可能与成花诱导具有相关性[31-32];BoSPL10-2在Ⅰ时期中薹叶与真叶中的相对表达量较高,推测其与叶片发育相关[11];BoSPL11-2和BoSPL14-1在Ⅰ时期根中的相对表达量较高,说明BoSPL不同成员发挥作用的主要部位不同[4, 33]。在对‘改良香菇’叶片不同部位检测BoSPL3-1、BoSPL10-2和BoSPL11-2的相对表达量,各基因在叶肉中的相对表达量无明显差异,值得注意的是BoSPL10-2在花状变形叶中显著表达,推测其可能与花状变形叶形成相关,其具体机制如何有待进一步验证。
猜你喜欢
南方医科大学学报(2022年3期)2022-04-13
山西师范大学学报(自然科学版)(2021年3期)2021-11-02
浙江大学学报(农业与生命科学版)(2021年3期)2021-07-10
中国预防兽医学报(2020年8期)2020-11-05
三农资讯半月报(2020年15期)2020-08-25
饮食保健(2019年21期)2019-11-22
食品与健康(2016年6期)2016-06-08
中国蔬菜(2015年11期)2015-12-21
红领巾·探索(2015年9期)2015-09-10
农产品市场周刊(2014年8期)2014-05-15
