
晋阳湖丝状蓝藻的形态学分析与分子鉴定
2022-04-18 01:40:34王清华李艳晖谭梅娟
西北植物学报 2022年3期
肖 喆,王 捷*,石 瑛, 2,王清华,李艳晖,谭梅娟
(1 太原师范学院 生物系,山西晋中 030619,2 山西工程科技职业大学,山西晋中 030619)
晋阳湖位于山西省太原市晋源区,是华北地区最大的人工湖,享有“中国北湖”的美称。晋阳湖水体由汾河西干渠引入,与平原水体的性质相似,最初用途是为太原市第一热电厂提供冷却用水,现主要以其为主体构建城市公园[1]。蓝藻生存环境广泛,种类繁多,具有较高的研究价值和利用潜力。有研究表明,颤藻目(Oscillatoriales)和念珠藻目(Nostocales)等分类单元下的丝状蓝藻可固定大气中的氮转化为含氮化合物[2-3]。在研究结皮微藻多样性时,发现细鞘丝藻亚科和颤藻科(Oscillatoriaceae)等中的丝状蓝藻可与沙土黏附以便形成生物结皮[4-6],改善土壤环境。颤藻目和聚球藻目(Synechococcales)等中的蓝藻在生产生物燃料等工业方面也具有良好的前景[7]。但某些种类在富营养化水体中产生水华,并成为藻源毒素主要的制造者。晋阳湖水体中生活着大量的藻类,目前对于晋阳湖的蓝藻甚至是其他藻类研究较少。因此,对于晋阳湖丝状蓝藻的研究可为今后该湖藻类资源利用提供一定的理论基础。
蓝藻为富氧光合自养藻类,形态简单、独特且多样,在多种环境中存活[8],甚至可生活在温度极高或极低的恶劣环境中[9]。中国对于蓝藻分类学研究起步较晚,在解放后才正式开始[10],胡鸿钧和魏印心等科学家为了中国藻类学的科研工作、教学及生产所需,将中国淡水藻类大多数属重新整理,并编写成《中国淡水藻类——系统、分类及生态》一书,该书已成为国内藻类分类学研究的重要依据之一[11]。藻类分类学的主要工作包含命名、形态特征描述、分群归类,并利用系统发育分析其亲缘关系。近年来,蓝藻分类不再只依靠形态学,而是基于多相分类方法(生态学、形态学、生物化学和分子数据等相结合)定义新类群和经典藻类的分子表征[12-13]。颤藻目和聚球藻目中包含许多鉴定不明确的物种,分子分析证明它们是多系的,给分类工作增加了极大的阻力。
早期的分类系统中,颤藻目包括不含具异形胞的丝状蓝藻。2014年,Komrek等根据类囊体的超微结构及系统发育分析将颤藻目分为两个目:颤藻目与聚球藻目[14]。目前,聚球藻目下有12个科:Heteroleibleiniaceae,细鞘丝藻亚科(Leptolyngbyaceae),Oculatellaceae,伪鱼腥藻科(Pseudanabaenaceae),Romeriaceae,裂须藻科(Schizotri-chaceae),Acaryochloridaceae, 管胞藻科(Chamaesiphonaceae), Coelosphaeriaceae, 平裂藻科(Merismopediaceae), Prochloraceae和聚球藻科(Synechococcaceae)。其中前6科包含丝状蓝藻形态的藻种[15]。颤藻目的成员其形态差异较小,但遗传多样性和变异性强,且均不含异形胞[16-17]。当环境中的水温和营养物(尤其是磷)浓度高,水的流动性不强时,颤藻的生长趋于旺盛。因此,水库、湖泊和水坝等地的颤藻会频繁出现[18]。颤藻在生长过程中可固定CO2,并生成生物燃料等物质,但有些颤藻可产生藻毒素,过度繁殖可污染水资源,对水生动物和人体健康造成损害[19]。对晋阳湖水体中丝状蓝藻种类及数量调查是亟待研究解决的问题。
本研究对晋阳湖及周边地区的藻类进行采集与分类,观察并记录了藻类的生存环境及生态分布。分离纯化其中的丝状蓝藻,观察藻丝长度、细胞长宽、顶端细胞形态等,同时运用16S rRNA基因序列与NCBI数据库中同源性较高的序列构建系统发育树,确定丝状蓝藻的遗传属性,以期对晋阳湖蓝藻种类有进一步的了解,丰富该地区蓝藻的多样性,研究结果对预防湖泊蓝藻水华的发生起到预警效果,对维护水资源环境稳定与生态平衡提供科学数据,并为后续晋阳湖蓝藻资源利用与开发提供理论依据。
1 材料和方法
1.1 藻种采集和分离
样品采集地为山西省太原市晋阳湖,采样时间为2019和2020年3月、6月、9月和11月4个月份,每月2次采集水体中的藻类植物,使用25号浮游生物网和镊子、刷子等工具分别采集浮游和石头等附着的藻类,采集地的详细经纬度信息见表1,采集点位置在图1标出。将采集到的新鲜藻样带回实验室镜检,并利用经典的毛细管分离法[15]分离纯化藻类,首先在倒置显微镜下挑取单个藻丝体,使用无菌水多次清洗,然后放入含有BG-11培养基[20]的24孔细胞培养板中培养,确定为单克隆培养后可转入到三角瓶中扩大培养。培养温度为(25±1)℃,光周期为12 h∶12 h。纯化后的藻种保存在太原师范学院淡水藻种库中,其编号分别为JYH005、JYH008、JYH010、JYH012和JYH022。
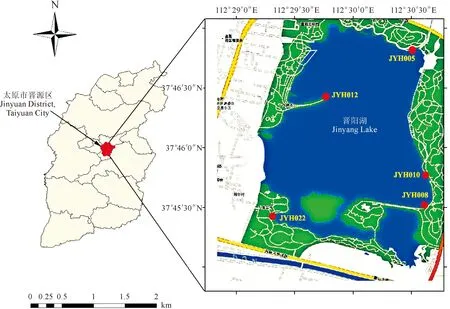
图1 红色点显示晋阳湖的采样地点
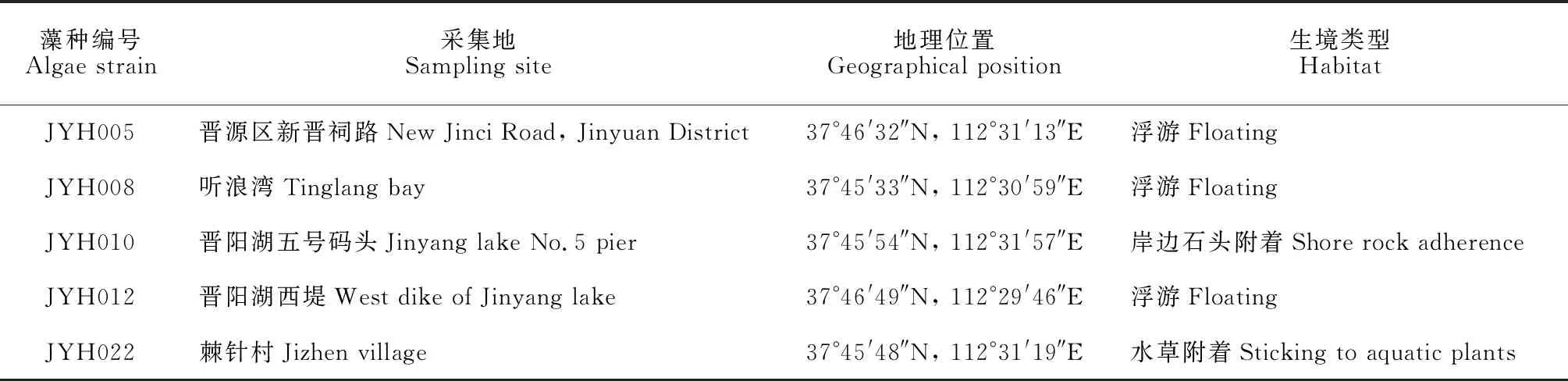
表1 山西省晋阳湖采样信息
1.2 显微观察和测量
将蓝藻藻丝体制作成显微玻片标本,在Nikon ECLIPSE NI型光学显微镜下观察,使用外接电脑的NIS-Elements D5.20软件拍摄藻种的形态特征,并测量与分析藻丝、细胞长宽等数据。每个样本至少测定30个细胞的大小、藻体形态、鞘的形态、末端细胞的形状等数据,并对其形态进行观察和描述。
1.3 基因组DNA提取和PCR扩增
DNA提取的操作步骤:取适量藻液离心,去除上清培养基,大约浓缩1.5 mL新鲜的藻液来提取DNA。加入无菌蒸馏水,使用冻融法破除细胞。为使细胞裂解充分,加入PlantZol裂解液并金属浴。然后利用DNA提取酚∶氯仿∶异戊醇(体积比为25∶24∶1)多次抽提上清液,使蛋白质及多糖等杂质变性沉淀,离心后去除沉淀。最后利用DNA不溶于乙醇的特点沉淀出DNA。室温挥发乙醇,加入无菌蒸馏水悬浮DNA,保存在冰箱中。
PCR扩增:扩增16S rRNA序列所用引物为F1和1 492[21-22]。PCR反应体系为20 μL,包含2.5 mmol·L-1dNTP 混合液2 μL,5 U·μL-1taq DNA 聚合酶 0.2 μL,10 μmol·L-1正反引物各2 μL,10× Easy Taq Buffer 2 μL,基因组DNA 1 μL,ddH2O为10.8 μL。PCR的条件如下:94 ℃预变性3 min;94 ℃变性30 s,55 ℃退火30 s,72 ℃延伸50 s,此阶段循环35次;最后72 ℃延伸10 min。PCR产物点样于1%琼脂糖凝胶并电泳,检验产物,以确保获得预期扩增的序列。将纯化的PCR产物测序,测序工作交由北京华大基因公司完成。
1.4 系统发育树分析
测序获得的序列为ABI和SEQ两种文件类型,可用BioEdit 7.0软件处理,并上传在NCBI数据库中BLAST进行对比,下载不同相似度的同源序列,基于16S rRNA基因的序列在BioEdit7.0软件中进行多重序列比对,然后手工编辑序列。系统发育树是在MEGA7.0软件中构建,邻接法(neighbor-joining method, NJ)和最大似然法(maximum likelihood, ML)使用的模型为Kimura 2-parameter model[23]。所有的系统发育评估均计算1 000次重复。同样利用MEGA7.0软件通过最大似然分析(ML)验证了树状拓扑结构的可靠性。
2 结果与分析
2.1 形态学分析
根据光学显微镜下观察到的形态特征,鉴定出4株丝状蓝藻属于聚球藻目,其中2株属于细鞘丝藻亚科,2株属于伪鱼腥藻科,1株丝状蓝藻属于颤藻目分类单元下的沙丝藻科。表2和图2详细描述了分离到的5株丝状蓝藻的形态特征。

A、B.JYH005,具明显的鞘;C、D.JYH008, 含死细胞,偶尔具鞘;E、F.JYH010,具鞘,末端圆柱状或渐细;G、H.JYH012, 藻丝体偶尔缠绕; I、J.JYH022,具鞘

表2 本研究中丝状蓝藻的形态学描述
2.2 分子系统分析
分离纯化得到的5株丝状蓝藻的系统发育关系如图3所示,以GenBank数据库中下载的紫色粘菌藻(Gloeobacterviolaceus)序列为外类群构建系统发育树。使用NJ、ML和MP方法构建的系统发育树拓扑结构基本一致,本研究以MP系统发育树为基准,标注了3种方法构建系统树所计算出的支持率。系统发育树中显示了所研究的5株丝状蓝藻聚为3个不同的大支系,JYH005与JYH012为细鞘丝藻亚科,JYH008与JYH022为伪鱼腥藻科,JYH010为沙丝藻科。

节点处数字代表NJ,ML和MP构建系统发育树的支持率,低于50%的未显示
对JYH005、JYH008、JYH010、JYH012和JYH022五株丝状蓝藻进行16S rRNA基因序列分析,将扩增出的序列片段与BLAST数据库中的序列对比表明,藻种JYH005与GenBank数据库中的结节结丝藻(NodosilineanodulosaMH179051)相似度为98.9%;藻种JYH012与细鞘丝藻属Lep-tolyngbyamargaretheana(FR798934)的相似度为99.1%;这表明藻种JYH012和藻种JYH005与细鞘丝藻科的亲缘关系较近。藻种JYH008与Arthronemaafricanum(KM019913)的相似度为97%;藻种JYH022与伪鱼腥藻科蓝藻(KF246480)的相似度为97.2%,与其他蓝藻的相似度低于90%,无法确定其属的位置;所以藻种JYH008和藻种JYH022的亲缘关系与伪鱼腥藻科相近。藻种JYH010与塔尔沙丝藻(DesertifilumtharenseMW411006)的相似度为99.6%;与沙丝藻属的藻种聚为一支,说明两者的亲缘关系较近。
3 讨 论
蓝藻又被称为蓝细菌,因此蓝藻的分类受控于植物学和细菌学两个学科。传统分类主要依据藻类的形态特征,但某些蓝藻会表现出可塑性,藻种之间的进化关系难以准确构建[28]。现代的蓝藻分类修订和重新构建分类基于多相方法,蓝藻分类的首要标准即为确定其分类单元的系统发育位置[29]。分子生物学技术的发展对某些在形态方面难以分辨的蓝藻(如颤藻目)分类起到了巨大的作用。
基因16S rRNA高度保守,具有“生物进化分子钟”的称号。原核生物属及属以下水平的分类均可采用该基因序列,许多学者对这一基因序列的可靠性进行了检测,结果得到了肯定[30]。通常情况下,根据分离株的形态特征都可将其鉴定到属水平,某些物种具有非常特别的形态特征的话,也可鉴定到种水平[31]。蓝藻分子生物学研究对弥补传统形态学分类体系的缺陷起到了关键的作用,提供了更完善的基础基因数据,蓝藻的分类学研究日益趋于完善,更亲近于物种自然亲缘关系的图谱建立[32]。
本研究采用基于传统形态学和分子16S rRNA序列结合的方法,对5株晋阳湖丝状蓝藻进行了表征。在这5株丝状蓝藻中,JYH012属于细鞘丝藻属,该属由于缺乏典型的判断特征或与相关类群有重叠特征,仅依靠形态特征无法准确鉴定。事实上细鞘丝藻属的形态特征极其简单,主要表现在藻体薄,蓝绿色,无异形胞,藻丝体宽度小于3.5 μm,具鞘且每鞘只包裹着1个藻丝体,类囊体分布在细胞周围[33-34]。而JYH012藻株藻丝体极细,藻丝单生或缠绕,横壁收缢明显,细胞内偶尔可见颗粒状,这与该属的形态特征不同。JYH005属于结丝藻属,该属的模式种为结节结丝藻(Nodosilineanodulosa)[24],JYH005与该属的典型特征基本吻合,但是在培养过程中未观察到膨胀的鞘。JYH022为伪鱼腥藻科,由于该藻株序列信息与该科其他属的相似度低于90%,因此只能确定到科水平,物种之间的相似度低于94.6%即为不同属[35]。JYH008与Komrek等修订的属Arthronema[25]形态特征基本一致,但是已报道的Arthronema形态未见死细胞,而对JYH008形态观察时发现藻丝中有死细胞,可通过死细胞进行繁殖。JYH010形态基本符合沙丝藻属[26]所描述的形态特征,但是未在JYH010藻丝中观察到气囊,原因可能是实验室培养条件优越,造成了表型的改变。
本研究以从山西省太原市晋阳湖分离纯化出的5株丝状蓝藻为实验样品,利用光学显微镜观察其形态结构特征,并采用16S rRNA序列分析其系统发育关系,确定其分类单元,对山西淡水湖泊丝状蓝藻种类有了进一步的了解,为可能发生的水华的防控提供理论基础,也为深度利用晋阳湖丝状蓝藻生物资源提供有效信息。
猜你喜欢
生物化工(2022年4期)2022-09-20 09:18:08
当代水产(2019年9期)2019-10-08 08:02:42
当代水产(2018年8期)2018-11-02 05:30:42
农业工程学报(2017年3期)2017-03-04 02:48:56
中外医疗(2015年5期)2016-01-04 03:57:57
幼儿智力世界(2015年5期)2015-08-20 09:41:39
山西建筑(2015年17期)2015-06-05 09:37:07
恋爱婚姻家庭·养生版(2015年10期)2015-05-14 21:46:23
化工管理(2015年6期)2015-03-23 06:03:38
生态环境学报(2014年4期)2014-07-16 05:40:44
