
TGF-β1基因g.8666_8667delAC位点突变与大白猪繁殖性能的关系
2022-04-06 02:06王玲芳李尚来李琦琦杜星吴望军宋奔驰李齐发
南京农业大学学报 2022年2期
王玲芳,李尚来,李琦琦,杜星,吴望军,宋奔驰,李齐发*
(1.南京农业大学动物科技学院,江苏 南京 210095;2.宿豫区正博生猪养殖场,江苏 宿迁 223800)
转化生长因子β1(transforming growth factor β1,TGF-β1)是一种多功能生长因子,是TGF-β超家族中被研究最为广泛的一员,以细胞类型依赖性方式调节多种细胞活性,例如特异性地影响细胞活力、凋亡、增殖、分化、黏附、迁移和上皮-间充质化等[1-4]。研究发现,TGF-β1在各种正常的组织、细胞和转化细胞中均存在,且在卵巢组织中表达量较高[5]。TGF-β1在哺乳动物卵巢组织的多种细胞中均有表达,如卵母细胞、颗粒细胞和各级卵泡的卵泡膜细胞,并在卵泡液中检测到TGF-β1蛋白的存在[2,6]。TGF-β1可调节各种卵巢功能,包括类固醇激素合成、卵泡发育和黄体功能[7]。例如,TGF-β1可抑制人黄体化颗粒细胞凋亡[8]。TGF-β1还参与颗粒细胞与膜细胞、颗粒细胞与卵母细胞间相互交流,在促进卵母细胞成熟中发挥重要作用[9]。卵泡液中TGF-β1可以促进排卵、受精和早期胚胎发育,并能抑制颗粒细胞生长[6]。TGF-β1在猪黄体化颗粒细胞中可显著抑制促黄体素诱导的孕酮升高[10]。TGF-β1与母猪繁殖性能关系密切,研究发现妊娠母猪血浆中TGF-β1浓度与繁殖性能有关[11]。另外,TGF-β1基因突变位点多态性与母猪的产仔数性状显著关联[11-12]。吴艳萍[11]利用SSCP技术在大白猪、长白猪、皮特兰猪、圣特西猪和二花脸猪等猪种TGF-β1基因中发现5个突变位点[第4内含子32核苷酸(nt)处C>T突变,第6内含子179 nt处 G>A 突变和1 043 nt处C>T突变,3′-UTR 2 490 nt处G>A突变和2 494 nt处G>A突变],共形成2种单倍型CGCAA(记为A)和TATGG(记为K),关联分析发现大白猪群体中KK型母猪的初产总产仔数显著高于AA型,两者平均每胎相差1.02头。为了进一步研究TGF-β1基因多态性与母猪繁殖性能的关系,本文采用PCR和测序技术鉴定大白猪TGF-β1基因第4内含子全序列和突变位点,分析突变位点多态性与母猪繁殖性能的关系,以及突变影响母猪繁殖性能的机制,以筛选可用于大白猪繁殖性状分子育种的有效标记。
1 材料与方法
1.1 试验动物
325头大白母猪耳组织样来自江苏省康乐阳光猪场,记录各胎繁殖性能:总产仔数(TNB)、产活仔数(NBA)、死胎数(NSW)和出生窝重(LW),共1 145条记录。猪新鲜卵巢来自江苏竹顺生物科技有限公司。
1.2 引物设计
根据GenBank数据库(https://www.ncbi.nlm.nih.gov/)中杜洛克猪TGF-β1基因组序列(GenBank ID:397078)和mRNA序列(GenBank ID:NM_214015.2),利用Primer Premier 5.0软件设计引物(表1)。引物由北京擎科生物有限公司合成。

表1 本试验所用PCR引物
1.3 PCR扩增和测序
采用苯酚/氯仿抽提法提取猪耳组织样或卵巢组织样基因组DNA,使用分光光度计(Thermo公司)测浓度后稀释至50 ng·μL-1,置于-20 ℃冰箱保存,以用于后续试验。PCR反应体系(20 μL):上游引物 1 μL、下游引物 1 μL、DNA模板1 μL、2 × VazymeLAmp®Master Mix(南京诺唯赞公司)10 μL、ddH2O 7 μL。PCR反应程序:95 ℃ 2 min;95 ℃ 10 s,退火15 s,72 ℃ 40 s,30个循环;72 ℃ 5 min;4 ℃保存。PCR产物用15 g·L-1琼脂糖凝胶电泳分离,切胶、回收,由上海生工生物有限公司测序。
1.4 突变位点筛选、分型和关联性分析
以大白猪DNA混池(n=20)为模板,利用引物对P1(表1)扩增和测定TGF-β1基因第4内含子序列,根据峰图寻找潜在的突变位点。利用引物对P2(表1)对单个DNA样品进行PCR扩增和测序,确定突变位点,再对大白猪群体(n=325)进行分型。根据文献[13]中的方法统计大白猪群体中突变位点的基因型频率和等位基因频率,利用SAS 9.2软件(SAS Institute Inc)进行突变位点多态性与繁殖性状的关联性分析。
1.5 克隆测序和序列分析
从猪新鲜卵巢上直径为3~5 mm卵泡中抽取卵泡液,分离颗粒细胞,加入1 mL TRizol试剂(Invitrogen司),用TRizol-氯仿法提取颗粒细胞总RNA。测定RNA浓度后,用反转录试剂盒(诺唯赞公司)将RNA反转录成cDNA,保存在-20 ℃备用。具体方法参考文献[14]。利用引物对P3(表1)扩增猪卵巢颗粒细胞中TGF-β1基因编码区序列,扩增产物切胶、回收后,按文献[13]中的方法进行克隆并测序。利用在线软件miRBase(http://www.mirbase.org/)预测潜在的miRNA结合位点;通过Jaspar数据库(http://jaspar.binf.ku.dk/cgi-bin/jaspar_db.pl)预测转录因子结合位点;利用在线软件MethPrimer(http://www.urogene.org/methprimer2/)预测CpG岛(CGI),参数:序列长度>100 bp,CG含量>60%,观察CpG数/预期CpG数>0.7。
2 结果与分析
2.1 大白猪TGF-β1基因第4内含子序列扩增
利用引物对P1(表1)对大白猪耳组织样基因组DNA进行PCR扩增,扩增产物用15 g·L-1琼脂糖凝胶进行电泳检测,结果得到单一清晰的条带(图1)。测序发现扩增片段与GenBank中杜洛克猪TGF-β1基因相应序列的一致性为99.69%。
图1 大白猪TGF-β1基因第4内含子PCR扩增产物电泳图
2.2 大白猪TGF-β1基因第4内含子序列分析
大白猪TGF-β1基因第4内含子序列全长189 bp(图2-A),序列中A、T、C、G碱基含量分别为13.23%、21.69%、39.15%和25.92%,其中A+T含量34.92%,C+G含量65.07%,可见C+G含量明显大于A+T含量。序列比对发现大白猪TGF-β1基因第4内含子序列与杜洛克猪的一致性为98.94%,比后者多2个碱基。在大白猪TGF-β1基因第4内含子序列的131~142 nt处发现一个由6个GT二核苷酸重复组成的微卫星序列。利用生物信息学软件对大白猪TGF-β1基因第4内含子上潜在的顺式作用元件(cis-acting elements)进行了预测,结果在这个区域预测到1个猪miRNA(ssc-miR-574-3p)结合位点和芳香烃受体(AHR)、Nrf2相关因子1(NRF1)和缺氧诱导因子1α(HIF1A)等多个与雌性生殖相关转录因子的结合位点。另外,利用在线软件MethPrimer在大白猪TGF-β1基因第4内含子32~132 nt处预测到1个CGI,长度101 bp,含有14个CpG位点(图2-B)。

图2 大白猪TGF-β1基因第4内含子核苷酸序列分析(A)和CpG岛预测(B)
2.3 大白猪TGF-β1基因第4内含子突变位点筛选
以大白猪池DNA为模板,利用引物对P1(表1)进行PCR扩增和测序,在大白猪TGF-β1第4内含子63~64核苷酸(nt)处发现1个2 bp序列(AC)的插入/缺失突变(图3)。由于这个插入/缺失突变位于猪TGF-β1基因的8 666~8 667 nt处,因此命名为g.8666_8667delAC。但本试验并没有检测到武艳萍[11]在大白猪TGF-β1基因第4内含子中发现的32 nt处C>T突变。

图3 大白猪TGF-β1基因g.8666_8667delAC突变位点鉴定
2.4 大白猪TGF-β1基因g.8666_8667delAC位点多态性分析
大白猪群体(n=325)TGF-β1基因g.8666_8667delAC位点分型显示大白猪群体中含有3种基因型,分别定义为ins/ins型、ins/del型和del/del型(图4-A)。基因型频率计算发现ins/ins基因型频率最高(57.8%),在大白猪群体中为优势基因型(图4-B)。等位基因型频率计算结果显示等位基因ins的频率最高(76.2%),为优势等位基因(图4-C)。遗传多样性分析发现大白猪群体中TGF-β1基因g.8666_8667delAC位点的多态信息含量(PIC)为0.36,为中度多态性位点(0.25
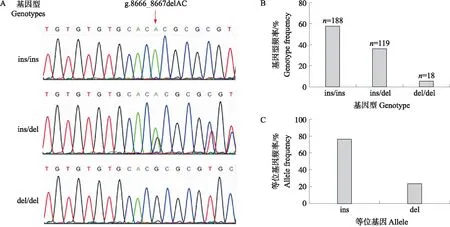
图4 TGF-β1基因g.8666_8667delAC位点多态性分析
2.5 g.8666_8667delAC位点多态性与大白猪繁殖性能关联性分析
关联性分析结果显示:ins/del基因型母猪的初产死胎数(NSB)显著高于ins/ins基因型和del/del基因型母猪(P<0.05)(表2)。在所有胎次中,del/del基因型母猪的总产仔数(TNB)比ins/ins基因型和ins/del基因型母猪每胎分别高0.47和0.46头,虽未达到显著水平(P>0.05),但每胎相差近0.50头,在养猪实践中也有一定的意义。

表2 大白猪TGF-β1基因g.8666_8667delAC位点多态性与繁殖性状的关联分析
2.6 g.8666_8667delAC突变对TGF-β1基因mRNA剪接的影响
内含子突变往往通过改变顺式作用元件影响基因转录或mRNA稳定性,或通过选择性剪接影响基因的功能[15-16]。分析发现g.8666_8667delAC突变未引起大白猪TGF-β1基因第4内含子顺式作用元件的改变(图2-A)。为了分析g.8666_8667delAC突变对猪卵巢颗粒细胞中TGF-β1基因选择性剪接的影响,本试验分离了g.8666_8667delAC位点ins/ins基因型和ins/del基因型猪卵巢颗粒细胞,克隆测序发现同一基因型内和不同基因型间卵巢颗粒细胞中TGF-β1基因编码区序列长度完全一致,说明g.8666_8667delAC突变对猪卵巢颗粒细胞中TGF-β1基因选择性剪接没有影响。大白猪TGF-β1基因编码区序列长度1 173 bp,与人、猕猴、牛、猪、绵羊和小鼠等哺乳动物一样,也由390个氨基酸残基组成,同样含有经典的N-端疏水信号肽、潜伏相关肽和TGF-β样结构域(图5)。大白猪TGF-β1蛋白氨基酸序列与杜洛克猪的同源性为100%,与人、猕猴、牛和小鼠等的同源性分别为94.36%、94.62%、94.62%和89.23%,说明哺乳动物TGF-β1蛋白氨基酸序列比较保守。
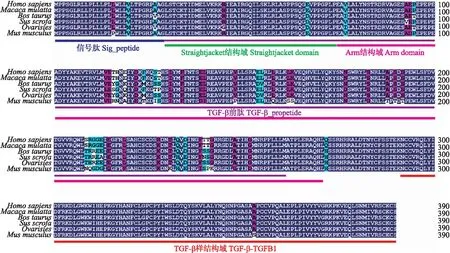
图5 大白猪和哺乳动物其他物种TGF-β1基因编码蛋白氨基酸序列比对分析
3 讨论
TGF-β1是一个重要的雌性动物卵泡发育命运(成熟排卵或闭锁退化)决定因子,影响雌性繁殖力[17-18]。在猪卵巢组织中,TGF-β1在卵母细胞和颗粒细胞中高表达,在体外培养的颗粒细胞中添加 TGF-β1 可促进颗粒细胞增殖,抑制颗粒细胞凋亡,调节雌激素(E2)和孕酮(P4)等类固醇激素的分泌[13,19-20]。进一步分析发现,TGF-β1可调节猪卵巢颗粒细胞状态(增殖或凋亡)相关信号通路如FoxO、TGF-β、Wnt、PIK3-Akt、p53和Ras等通路信号分子的表达;同时还发现多个已知的影响母猪繁殖性状的候选基因,如胰岛素样生长因子结合蛋白3基因(IGFBP3)[21]和WNT家族成员10B基因(WNT10B)[22-23]等也是TGF-β1的下游靶基因。本文在TGF-β1基因第4内含子上发现1个新的与大白猪繁殖性能显著关联的突变位点g.8666_8667delAC,进一步证实TGF-β1基因是大白猪繁殖性状的关键基因。但我们并未在研究的大白猪群体中发现前人鉴定的第4内含子上的另一个突变位点。武艳萍[11]研究发现第4内含子的32 nt处C>T突变位点与第6内含子的179 nt处G>A突变和1 043 nt处C>T突变、3′-UTR的2 490 nt处G>A突变和2 494 nt处G>A突变的合并基因型与大白猪初产TNB显著关联。Du等[13]研究发现 miR-130a 启动子区-573G>A、-540T>C位点与TGF-β1基因c.1583A>G位点合并基因型与大白猪NBA、NSB和LW显著关联。
目前一些功能和作用机制被注释的与家畜重要经济性状显著关联的突变位点主要为启动子区、编码区和3′-UTR上的突变位点[13,24],而大量的内含子区突变位点则很少被关注[25]。实际上,内含子突变与宿主基因的转录、mRNA稳定性和功能关系密切。研究发现内含子可作为基因内部启动子或增强子,这些内含子突变可引起与转录因子结合力的变化,从而促进或抑制基因的转录[26-27]。在海肾萤光素酶的开放阅读框(ORF)中,插入人磷酸丙糖异构酶(TPI)基因的第6内含子序列,可以增强海肾萤光素酶基因的表达[28]。Li等[25]在骨形态发生蛋白受体IB(BMPR1B)基因第1内含子中ESR1反应元件(ERE)上发现 4个突变位点,高繁殖力猪种特异的单倍型ERE与转录因子ESR1结合力强,子宫中BMPR1B基因mRNA和蛋白表达水平高。另外,内含子突变还通过影响基因的选择性剪接来影响基因的功能[15-16],研究发现A型血友病患者F8基因中的内含子突变c.6430_3C>G导致F8基因剪接效率低或剪接错误,影响F8蛋白的正常功能[16]。本文研究发现g.8666_8667delAC突变未引起大白猪TGF-β1基因第4内含子上顺式作用元件的改变,也不影响猪卵巢颗粒细胞中TGF-β1基因的选择性剪接。因此,g.8666_8667delAC位点突变影响大白猪繁殖性能的机制尚需进一步研究。
综上,本文获得了大白猪TGF-β1基因第4内含子全序列,发现了大量的顺式作用元件,鉴定到1个新的小插入/缺失突变(g.8666_8667delAC),其多态性与大白猪繁殖性状显著关联,证实TGF-β1基因是影响大白猪繁殖性状的关键基因,其中del/del基因型是TNB的有利基因型,ins/del基因型母猪的NSB较高。另外,本文还证实第4内含子的g.8666_8667delAC位点突变不是通过影响猪卵巢颗粒细胞中TGF-β1基因的选择性剪接来影响繁殖性状的。
猜你喜欢
中国畜牧杂志(2022年11期)2022-11-17
中国畜牧兽医(2022年8期)2022-08-23
当代畜禽养殖业(2022年3期)2022-07-13
家畜生态学报(2021年7期)2021-07-28
内蒙古师范大学学报(自然科学汉文版)(2021年3期)2021-06-01
湖北农业科学(2020年22期)2020-12-18
今日农业(2020年17期)2020-12-15
山地农业生物学报(2020年2期)2020-11-09
现代农业科技(2020年15期)2020-08-16
视野(2020年9期)2020-05-11
