
5-Aza-cdR通过DNA甲基化调节BDNF参与学习记忆的机制*
2022-03-01 01:26张世纪贾小娥巴德仁贵姜树原谢雅彬霍东升刘晓蕾
包头医学院学报 2022年1期
张世纪,付 翁,贾小娥,巴德仁贵,姜树原,谢雅彬,霍东升,谢 伟,刘晓蕾,邵 国
(1.包头医学院内蒙古自治区低氧转化医学重点实验室,内蒙古 包头 014040;2.包头医学院基础医学与法医学院;3 包头医学院医学技术与麻醉学院神经科学研究所)
5-氮-2’脱氧胞苷( 5-Aza-2'-deoxycy- tidine, 5-Aza-cdR)是一种有效的 DNA 甲基化抑制剂。5-Aza-cdR 可以与DNA甲基转移酶活性位点的半胱氨酸之间形成共价键,从而抑制DNA甲基转移酶的作用。有研究发现5-Aza-cdR 对小鼠学习记忆的影响可能与小鼠海马体中p-Y1472 NR2B磷酸化的增加有关。尽管研究表明5-Aza-cdR 可以影响学习记忆,但相关分子机制有待深入研究[1]。
脑源性神经营养因子(brain-derived neurotrophic factor, BDNF)是一种碱性蛋白,是神经营养因子之一。BDNF在下丘脑、杏仁核、海马、新皮质等脑区高表达,参与各种神经活动[2]。BDNF的表达和分泌,提升海马锥体神经元的树突状棘密度,并诱导海马长时程增强产生。有研究证实,提高BDNF的水平可以改善衰老导致的记忆障碍。还有研究发现BDNF可以通过参与神经细胞的生长发育及调节突触可塑性等方面,调节大脑海马区的学习记忆功能[3]。
BDNF在突触可塑性和学习记忆中的作用已经被证实是不可或缺的,而最新研究发现 DNA 甲基化可以通过调控BDNF表达来影响学习记忆[4]。然而DNA甲基化参与BDNF表达和调节学习记忆能力的机制尚不清楚。因此,本文通过研究5-Aza-cdR影响DNMT功能后,对BDNF表达以及学习记忆的影响,从而为后续DNA甲基化研究特别是在学习记忆方面提供理论依据。
1 材料与方法
1.1主要试剂与仪器 离心机、超声波破碎仪、恒温摇床、酶标仪、金属浴、电泳仪、转膜仪、化学发光仪、实时荧光定量PCR仪、激光共聚焦显微镜、BCA浓度定量试剂盒,抗氧化剂、PVDF膜、10XTBS、吐温20、Trizol裂解液、反转录试剂盒、SYBR Green mix、引物、一抗、96孔板、二抗等。
1.2动物及分组 6~8周龄的SPF级雄性ICR小鼠70只,体质量18~22 g,购自于斯贝福 (北京) [SCXK ( 京) 2016-0002]生物技术有限公司,所有小鼠在自然通风饲养室中自由摄入水和食物,适应性饲养24 h。注射PBS为C组,25只;注射5-Aza-cdR 为注射组,30只;注射K252a为K252a注射组,15只。本研究经包头医学院实验动物伦理委员会批准。
1.3动物处理 小鼠用1.2 %的戊巴比妥钠溶液麻醉,以俯卧位固定于脑立体定位仪上。消毒,在头顶切开一长约1.5 cm的切口,按照标准小鼠脑立体定位图谱,以十字缝交叉点为原点,按照十字缝原点右1 mm、十字缝前0.5 mm的位置上为点做标记,使用颅骨钻孔器钻孔;钻孔后以孔位为原点,使注射器针头向下进入2.5 mm,缓慢注射10 μL 10 μ/M 5-Aza-cdR 或10 μL10 μ/M K252a工作液。注射后停针5 min,使药品充分进入脑内,然后缓慢退针。缝合切口后放入单独鼠笼保温等待苏醒。C组注射等量0.01M PBS缓冲液。 注射5-Aza-cdR 或K252a 24h后,将小鼠断头处死,在冰上剥离海马,置于1.5 mL EP管中,立即放入-80 ℃冰箱中冻存。
1.4Real-time PCR检测 使用Trizol法提取总RNA,使用Thermo公司反转录试剂盒。将RNA反转录为cDNA,-20 ℃ 保存。引物由生物工程( 上海) 有限公司合成,DNMT3B基因、BDNF 基因和 β-Actin 基因引物的设计见表 1。Real-time PCR实验重复3次。

表1 Real-time 基因引物序列
1.5Western blot 电泳 将冻存于-80 ℃冰箱中的小鼠海马于冰上融化。每个海马样品中加入300 μLRIPA裂解液,蛋白酶抑制剂3 μL。使用超声波破碎仪破碎;4 ℃ 、12 000 rcf、离心15 min;吸取上清液到新的EP管中。使用BCA蛋白盒测定蛋白浓度。蛋白电泳时上样质量为20 μg,条件为先恒流19 mA,然后29 mA,转膜4 h,将蛋白转移到PVDF膜上。室温下使用5 %的脱脂牛奶35 r/min孵育1.5 h,使用1XTBST洗膜3次,每次15 min。一抗(1∶1000) 4°C过夜孵育,1XTBST洗膜三次,每次15 min。辣根过氧化物酶驴抗兔和山羊抗小鼠二抗(1∶1000) 35 r/min孵育1.5 h,1XTBST洗膜3次,每次15 min。ECL超敏发光液发光检测蛋白表达情况,实验重复3次。
1.6免疫荧光染色 小鼠使用1.2 %戊巴比妥钠麻醉后,心脏先后灌注30 mL生理盐水和30 mL 4 %多聚甲醛,取出全脑放入4 %多聚甲醛中后固定过夜,后更换30 %PB蔗糖脱水72 h,OTG包埋后进行冠状位冰冻切片,厚度20 μm。10 %NGS室温封闭1 h后分别使用BDNF和CaV1.2的特异性一抗4 ℃孵育过夜,0.3 %PBST清洗3次后室温使用Alexa Fluor 488、594避光孵育4 h,0.3 %PBST清洗3次后Hoechst 33342 室温避光染核后封片。每组实验重复 3 次以上。
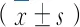
2 结果
2.1小鼠海马中DNMT3B表达变化 与C组相比,5-Aza-cdR 注射组小鼠海马中DNMT3B蛋白表达水平降低(P<0.05)(图1A);与C组相比,5-Aza-cdR 注射组小鼠海马中DNMT3B mRNA表达水平也降低(P<0.05)(图1B)。
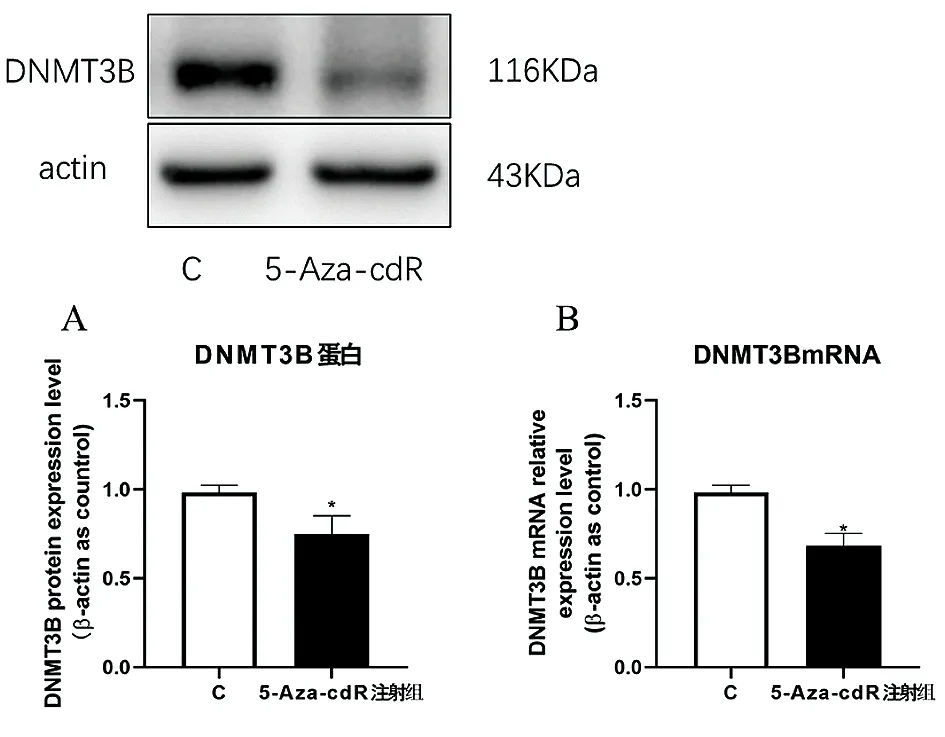
图1 DNMT3B蛋白和RNA表达
2.2小鼠海马中BDNF表达变化 与C组相比,5-Aza-cdR 注射组小鼠海马中BDNF蛋白表达升高(P<0.05)(图2A);与C组相比,5-Aza-cdR 注射组小鼠海马中BDNF RNA表达也升高(P<0.05)(图2B)。
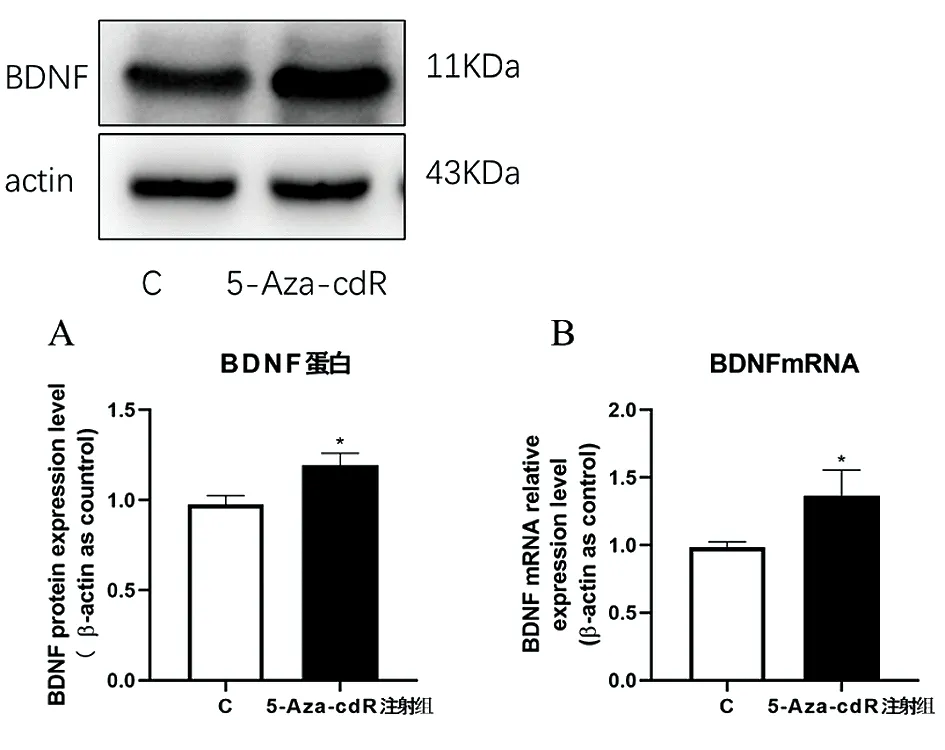
图2 BDNF蛋白和RNA表达
2.3小鼠海马区BDNF的表达变化 采用免疫荧光检测法,蓝色荧光为细胞核,绿色荧光为BDNF蛋白,免疫荧光显示BDNF定位于小鼠海马脑区(图3A)。注射5-Aza-cdR 后,与C组相比小鼠海马脑区的BDNF荧光强度显著增强(P<0.05)(图3B)。
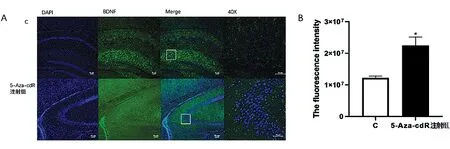
图3 不同处理组小鼠海通过免疫荧光检测BDNF变化的结果
2.45-Aza-cdR处理增强小鼠的空间学习记忆能力 与C组相比,5-Aza-cdR 注射组小鼠的潜伏期时间趋势在第1~6天明显变短(P<0.05);K252a注射组小鼠的潜伏期时间与C组相比明显变长(P<0.05)(图4D)。与C组相比,5-Aza-cdR 注射组小鼠在目标象限时间的百分比增加明显(P<0.05);K252a注射组小鼠的目标象限时间的百分比明显减少(P<0.05)(图4E)。与C组相比,5-Aza-cdR 注射组小鼠的穿梭平台次数明显增加(P<0.05);K252a注射组小鼠的穿梭平台次数明显减少(P<0.05)(图4F)。
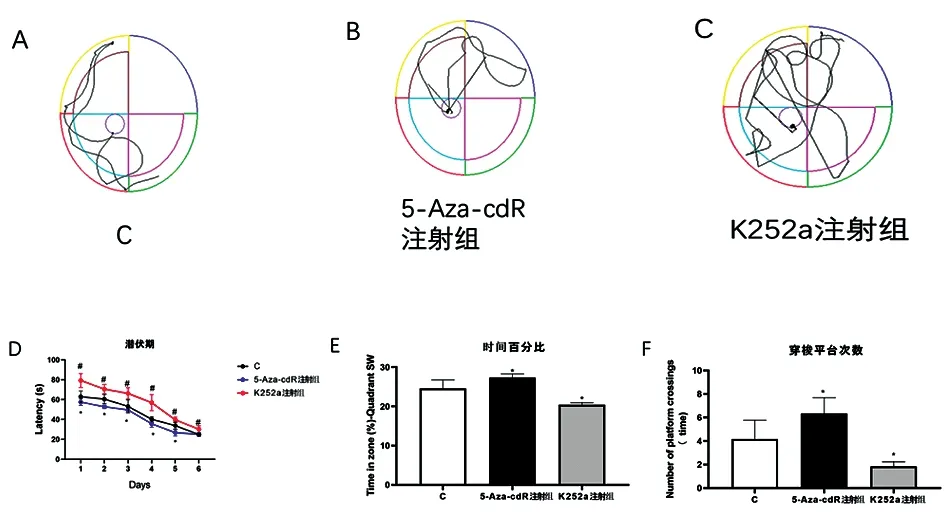
图4 水迷宫结果
3 讨论
5-Aza-cdR 是一种DNA甲基转移酶的抑制剂,已被批准用于各种条件的临床治疗。5-Aza-cdR可以提高水迷宫中小鼠的学习和记忆能力,并抑制PP1γ的表达来提高小鼠的学习记忆能力。5-Aza-cdR 研究了DNA甲基化在神经发育、分化、突触学习记忆可塑性以及主要神经元生存中的作用。研究发现,海马CA1区域的p-Y1472 NR2B受到5-Aza-cdR 的影响,对成人小鼠大脑的记忆形成和突触可塑性具有重要意义[5]。本研究得出在大鼠海马神经元中注射5-Aza-cdR后,DNA 甲基化水平显著降低,伴随BDNF基因表达上调,同时大鼠的学习能力也明显提升。
BDNF是德国神经生物学家 Barde在 1982年首次从猪脑中分离纯化出的一种碱性蛋白,是神经营养因子之一[6]。研究表明,增加 BDNF 的表达和分泌,可以提升海马锥体神经元的树突状棘密度,并诱导海马长时程增强(long-term potentiation,LTP)的产生[7];通过调节神经元分化,影响血清素能和多巴胺能神经传递。BDNF能在突触前和突触后靶位点充当旁分泌和自分泌因子,从而对突触活动转化为长期突触记忆至关重要。 BDNF 被认为是中枢神经系统中功能和结构可塑性的指导性介质。在海马体中BDNF还影响成人神经发生,进而对多种形式的学习和记忆产生作用[8]。
DNA 甲基化可以通过调控 BDNF 表达来影响学习记忆[4]。在大鼠海马神经元中,DNA 甲基化水平显著降低,伴随BDNF基因表达上调,同时大鼠的学习能力也明显提升[9]。还有研究指出,DNMT 抑制剂 RG108 可以在短时间内增加 BDNF 表达量,而且伴随着基因启动子中特定 CPG 岛甲基化的增加,可以增强神经元的突触可塑性,提高认知能力[10]。一部分脑卒中患者会同时患有焦虑障碍,而脑卒中患者 BDNF 基因的高甲基化则与焦虑有关;BDNF作为卒中后焦虑障碍的易感性标靶,应受到密切监测,以明确焦虑是否发生,特别是在卒中恢复期[11]。K252a是一种有效的丝氨酸/苏氨酸蛋白激酶抑制剂,抑制蛋白激酶C(PKC),PKA,钙/钙调蛋白依赖性激酶II型与磷酸酶激酶,可以阻断注射5-Aza-cdR对BDNF下游信号通路的影响。
先前的研究表明,5-Aza-cdR 可以降低DNMTs的表达,本研究提取小鼠海马进行Real-time PCR实验和Western blot 电泳实验发现5-Aza-cdR 可以抑制小鼠海马中DNMT3B的表达, DNA甲基化水平很可能因此降低,从而可以提高小鼠海马中BDNF的蛋白表达。通过水迷宫实验发现,BDNF参与了小鼠学习记忆的过程,也证实5-Aza-cdR在上调BDNF表达后,可能提升小鼠的学习记忆能力。由于在BDNF 启动子区域有大量CpG 岛,注射5-Aza-cdR 后,DNMT3B的水平降低,而BDNF的蛋白和RNA表达水平均升高。因此,5-Aza-cdR 可以抑制DNMT3B的表达,降低 BDNF 启动子区域的甲基化水平上调 BDNF 表达,从而提升小鼠的学习记忆能力。
有研究发现,在恐惧学习的环境中,可以快速增加记忆抑制基因蛋白磷酸酶1的甲基化,同时对可塑性相关基因的启动子区域去甲基化。恐惧学习的经验提高了大脑海马体中DNMT3A和DNMT3B的表达,对DNMT1 没有影响[12]。因此,DNA甲基化虽然对记忆的形成至关重要,但不太可能是一种长期储存的机制。 进一步的研究发现,在恐惧学习环境中,海马体中的BDNF外显子III和外显子IV启动子去甲基化,这表明DNA甲基化可以通过BDNF影响学习记忆[13]。
猜你喜欢
作文周刊·小学二年级版(2022年20期)2022-05-05
昆明医科大学学报(2022年1期)2022-02-28
昆明医科大学学报(2021年8期)2021-08-13
昆明医科大学学报(2021年6期)2021-07-31
中学生物学(2020年10期)2020-12-25
东方教育(2016年4期)2016-12-14
科技知识动漫(2016年6期)2016-06-24
医学研究杂志(2015年12期)2015-06-10
癌变·畸变·突变(2015年3期)2015-02-27
汽车观察(2009年1期)2009-02-18
