
自噬在结核菌素诱导破骨细胞形成中的作用及机制
2022-02-21 05:36王增顺索南昂秀刘立民周京元
脊柱外科杂志 2022年1期
王增顺,索南昂秀*,刘立民,周京元,伍 骥
1.青海省人民医院骨科,西宁 810000
2. 中国人民解放军空军特色医学中心骨科,北京 100142
结核病是结核分枝杆菌感染引起的呼吸道传染病,结核分枝杆菌经血液循环侵入骨关节后引起骨关节结核。脊柱结核是最常见的骨关节结核,以进行性骨质破坏为主要特征[1-2]。破骨细胞由前体细胞单核巨噬细胞分化而来,并介导骨质吸收,破骨细胞的异常增殖及分化在骨质破坏中起关键作用。多项骨关节结核的细胞实验研究[3-4]发现,结核菌素对单核巨噬细胞向破骨细胞的分化过程具有促进作用,但与这一作用相关的分子机制尚不十分清楚。
自噬是近年发现的与结核分枝杆菌感染、骨质疏松发生密切相关的生物学过程。在骨质疏松的发生过程中,自噬对破骨细胞的分化具有促进作用,使用自噬激动剂雷帕霉素干预后,单核巨噬细胞分化为破骨细胞的数量明显增多[5]。在结核分枝杆菌感染单核巨噬细胞的过程中,自噬激活明显,多种自噬基因(Atg5、LC3-Ⅱ/LC3-Ⅰ)的表达增加[6-7]。但结核分枝杆菌激活单核巨噬细胞自噬的作用是否与其促进破骨细胞形成有关并未明确。因此,本研究通过细胞实验来观察自噬在结核菌素诱导破骨细胞形成中的作用及机制,旨在为阐明脊柱结核发生过程中骨质破坏的分子机制提供参考。
1 材料与方法
1.1 材料
小鼠单核巨噬细胞(RAW264.7细胞)购自中国科学院上海细胞资源中心,结核菌素纯化蛋白衍生物(PPD)注射液(规格50 IU/mL、1 mL/支)购自北京祥瑞生物制品有限公司,自噬抑制剂3-甲基腺苷(3-MA)、自噬激动剂雷帕霉素均购自美国Sigma公司,MTS细胞活力检测试剂盒购自美国Promega公司,抗酒石酸酸性磷酸酶(TRAP)染色试剂盒、TRAP活力检测试剂盒购自南京建成研究院,Beclin-1、LC3抗体购自英国Abcam公司。
1.2 细胞培养及分组
RAW264.7细胞用含有10%胎牛血清的DMEM培养基进行贴壁培养,细胞密度达80% ~ 90%后用0.25%胰蛋白酶消化,传代后的细胞接种在培养板内并进行分组给药。对照组用不含药物的培养基处理,PPD组分别用含有1.0、2.5、5.0、10.0 IU/mL PPD的培养基处理,5.0 IU/mL PPD+激动剂组用含有 5.0 IU/mL PPD+100 nmol/L 雷帕霉素的培养基处理,5.0 IU/mL PPD+ 抑制剂组用含有5.0 IU/mL PPD+2 mmol/L 3-MA的培养基处理。雷帕霉素的剂量参照陆明等[8]的报道,3-MA的剂量参照徐亦文等[9]的报道。每组设置5个复孔。
1.3 细胞存活率的检测
将RAW264.7细胞接种于96孔培养板内,分组给药培养24 h后,采用MTS细胞活力检测试剂盒进行检测。每孔加入200 μL检测液,同时设置不接种细胞的检测液作为空白组,37℃孵育4 h后,在酶标仪上检测490 nm处的光密度值,细胞活力(%)=(实验组光密度值-空白组光密度值)/(对照组光密度值-空白组光密度值)×100%。
1.4 破骨细胞形成的检测
将RAW264.7细胞接种于6孔培养板内,分组给药培养7 d后,用4%多聚甲醛固定细胞,采用TRAP染色试剂盒进行染色,用苏木精复染。在显微镜下观察,随机选择5个视野,对TRAP阳性染色的破骨细胞进行计数。
1.5 TRAP活力检测
将RAW264.7细胞接种于6孔培养板内,分组给药培养7 d后,收集培养基,采用TRAP活性检测试剂盒进行检测。加入反应底物后孵育3 h,然后在酶标仪上检测540 nm波长处的吸光度值。
1.6 自噬小体的检测
将RAW264.7细胞接种于6孔培养板内,分组给药培养7 d后,用2.5%戊二醛溶液固定。用1%锇酸4℃处理1 h,经70%、80%、95%、100%乙醇溶液梯度脱水后,用环氧树脂包埋,-80℃聚合24 h,之后用3%柠檬酸铅染色。在透射电镜下观察,随机选择5个视野,对自噬小体进行计数。
1.7 自噬相关基因表达的检测
采用蛋白质印迹法检测自噬相关基因的蛋白表达。将RAW264.7细胞接种于12孔培养板内,分组给药培养7 d后,用RIPA裂解液提取细胞蛋白,测定蛋白含量。取30 μg蛋白样本加入SDS-聚丙烯酰胺凝胶中,电泳后电转移至PVDF膜,用5%脱脂牛奶在室温下封闭PVDF膜1 h。PVDF膜用1∶1 000稀释的 Beclin-1、LC3抗体或1∶2 500稀释的β-actin抗体4℃孵育过夜,次日洗膜3次后,再用1∶2 000稀释的二抗室温孵育1 h。在凝胶成像仪中曝光得到蛋白条带,测定灰度值,以Beclin-1、LC3-Ⅱ/Ⅰ与β-actin的比值作为蛋白相对表达量。
1.8 统计学处理
采用SPSS 22.0软件对数据进行统计分析,符合正态分布的计量资料以表示,组间比较采用单因素方差分析,差异有统计学意义的指标用LSD-t法进行两两比较;以P< 0.05为差异有统计学意义。
2 结 果
2.1 不同浓度PPD对RAW264.7细胞活力的影响
与对照组相比,1.0、2.5、5.0 IU/mL PPD 组细胞活力无明显变化,差异无统计学意义(P> 0.05,图1);10.0 IU/mL PPD组细胞活力降低,差异有统计学意义(P< 0.05,图1)。
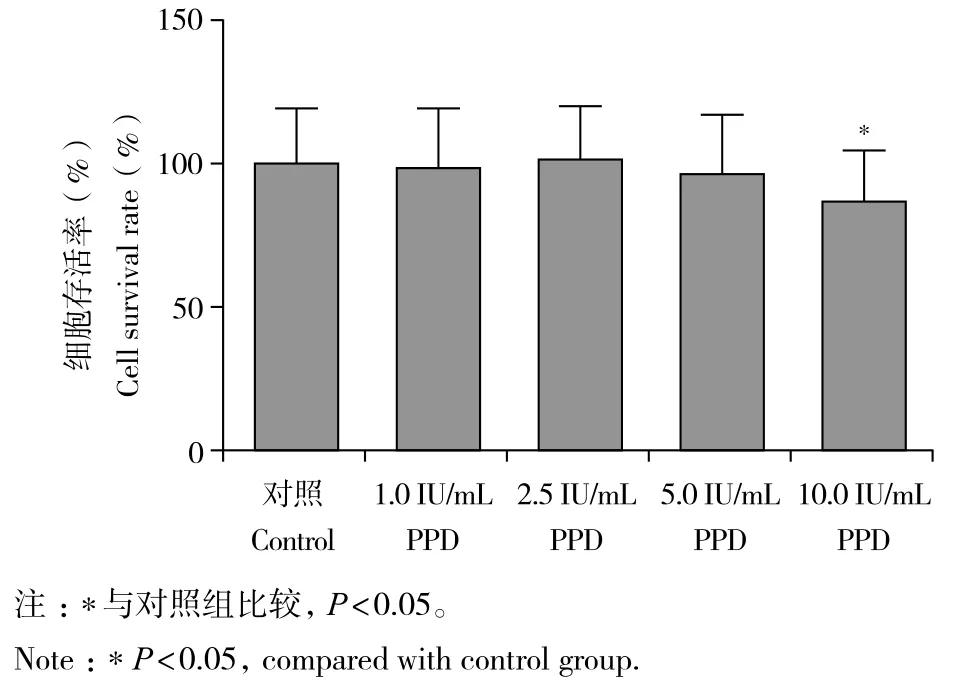
图1 不同浓度PPD对细胞活力的影响Fig. 1 Effect of PPD at different concentrations on cell viability
2.2 不同浓度PPD对RAW264.7细胞分化为破骨细胞的影响
与对照组相比,1.0 IU/mL PPD组破骨细胞数量及培养基中TRAP活力增高,但差异无统计学意义(P> 0.05,图2);2.5、5.0、10.0 IU/mL PPD 组破骨细胞数量及培养基中TRAP活力均显著增高,差异有统计学意义(P< 0.05,图2)。
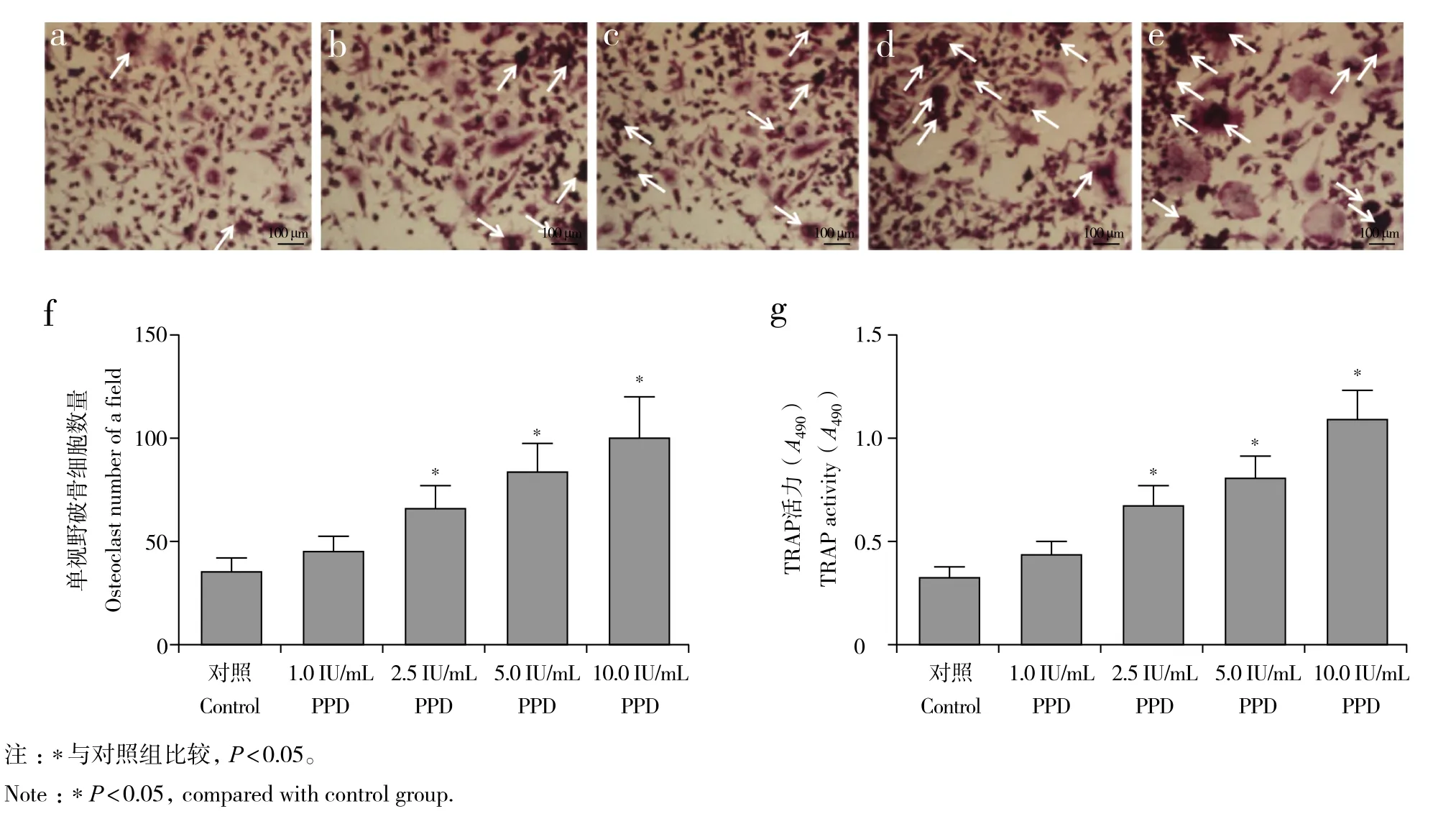
图2 不同浓度PPD对RAW264.7细胞分化为破骨细胞的影响Fig. 2 Effect of PPD at different concentrations on differentiation of RAW264.7 cells into osteoclasts
2.3 不同浓度PPD对RAW264.7细胞自噬水平的影响
与对照组相比,1.0、2.5、5.0、10.0 IU/mL PPD组RAW264.7细胞中自噬小体的数量及Beclin-1、LC3-Ⅱ/LC-Ⅰ的表达水平均明显增加,差异有统计学意义(P< 0.05,图3、4)。
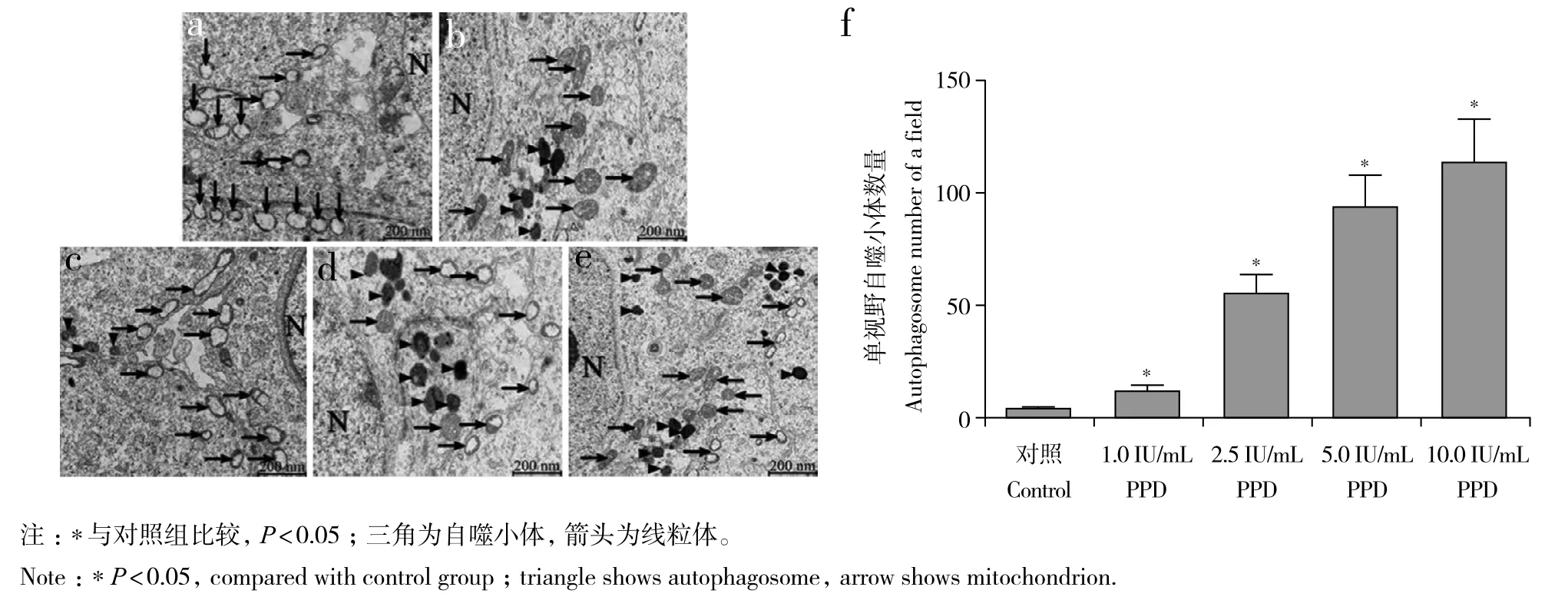
图3 不同浓度PPD对RAW264.7细胞中自噬小体数量的影响Fig. 3 Effect of PPD at different concentrations on number of autophagosomes in RAW264.7 cells

图4 蛋白质印迹法检测不同浓度PPD诱导后RAW264.7细胞中、LC3的表达Fig. 4 Expression of Beclin-1 and LC3 in RAW264.7 cells induced by PPD at different concentrations detected by Western blotting
2.4 自噬激动剂及抑制剂对PPD诱导后RAW264.7细胞自噬水平的影响
自噬激动剂及抑制剂对RAW264.7细胞活力均无明显影响(图5)。与对照组比较,5.0 IU/mL PPD组RAW264.7细胞中自噬小体数量及Beclin-1、LC3-Ⅱ/LC-Ⅰ的表达水平均明显增加,差异有统计学意义(P< 0.05,图6、7)。与5.0 IU/mL PPD 组比较,5.0 IU/mL PPD+激动剂组RAW264.7细胞中自噬小体数量及Beclin-1、LC3-Ⅱ/LC-Ⅰ的表达水平均明显增加,5.0 IU/mL PPD+ 抑制剂组RAW264.7细胞中自噬小体数量及Beclin-1、LC3-Ⅱ/LC-Ⅰ的表达水平均明显减少,差异均有统计学意义(P< 0.05,图6、7)。
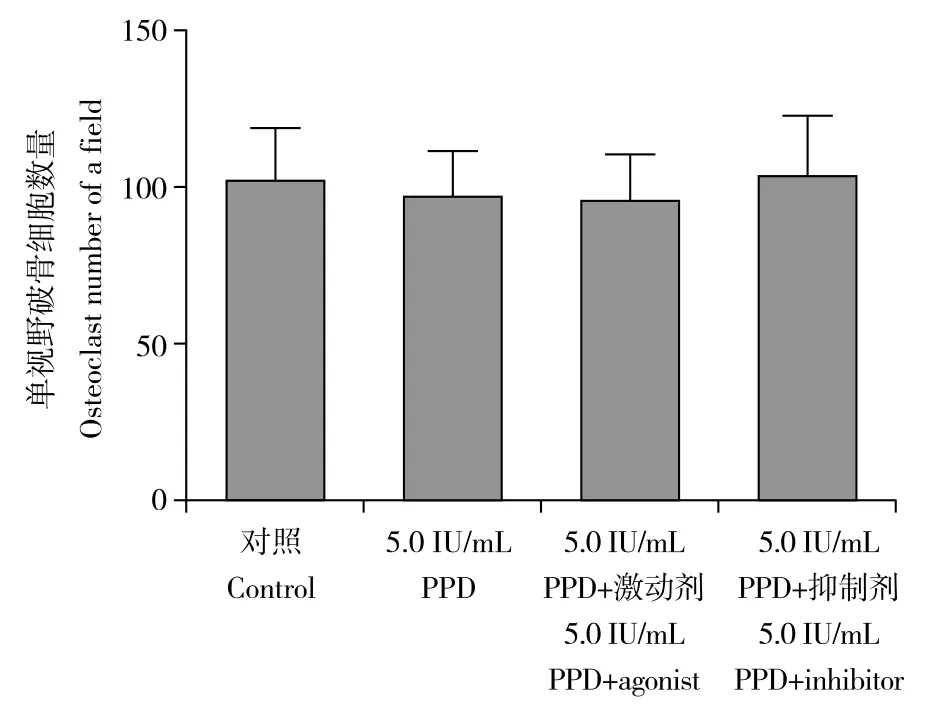
图5 自噬激动剂及抑制剂对5.0 IU/mL PPD诱导后RAW264.7细胞活力的影响Fig. 5 Effects of autophagy agonist and inhibitor on viability of RAW264.7 cells after 5.0 IU/mL PPD induction
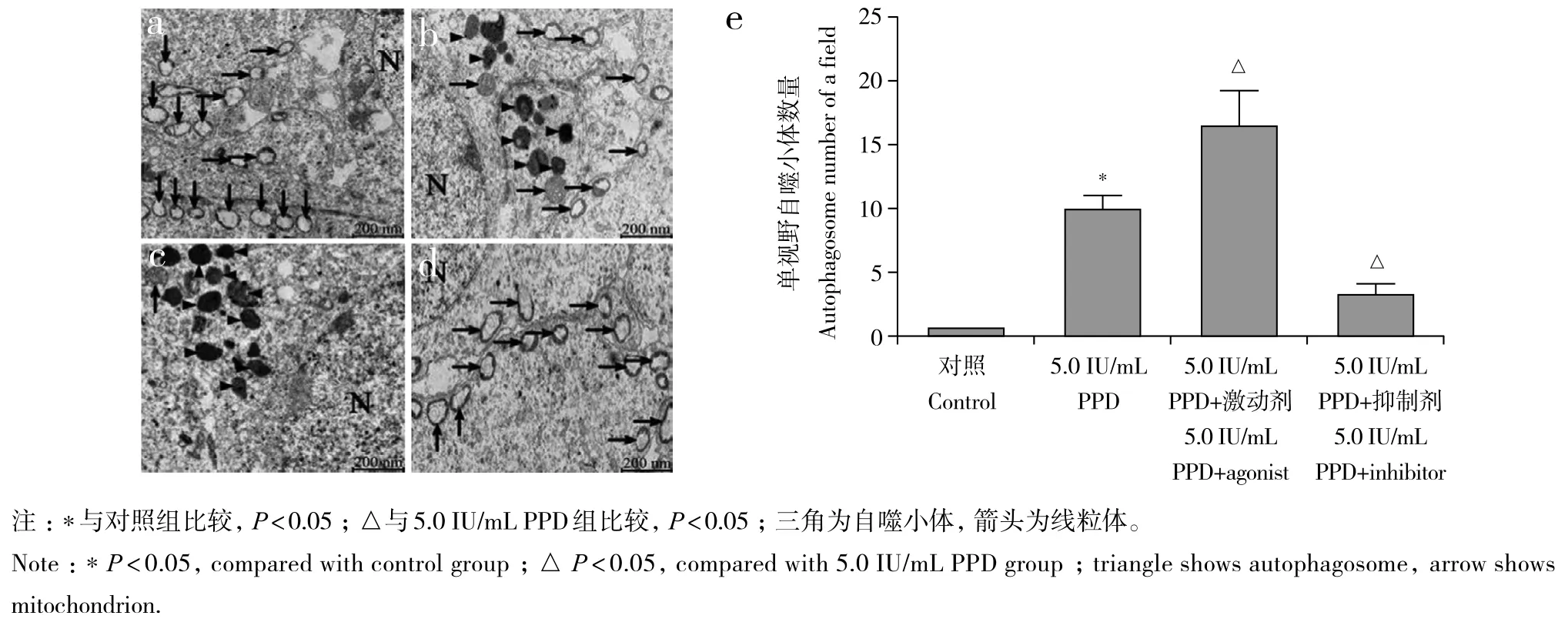
图6 自噬激动剂及抑制剂对5.0 IU/mL PPD诱导后RAW264.7细胞中自噬小体数量的影响Fig. 6 Effects of autophagy agonists and inhibitors on number of autophagosomes in RAW264.7 cells after 5.0 IU/mL PPD induction
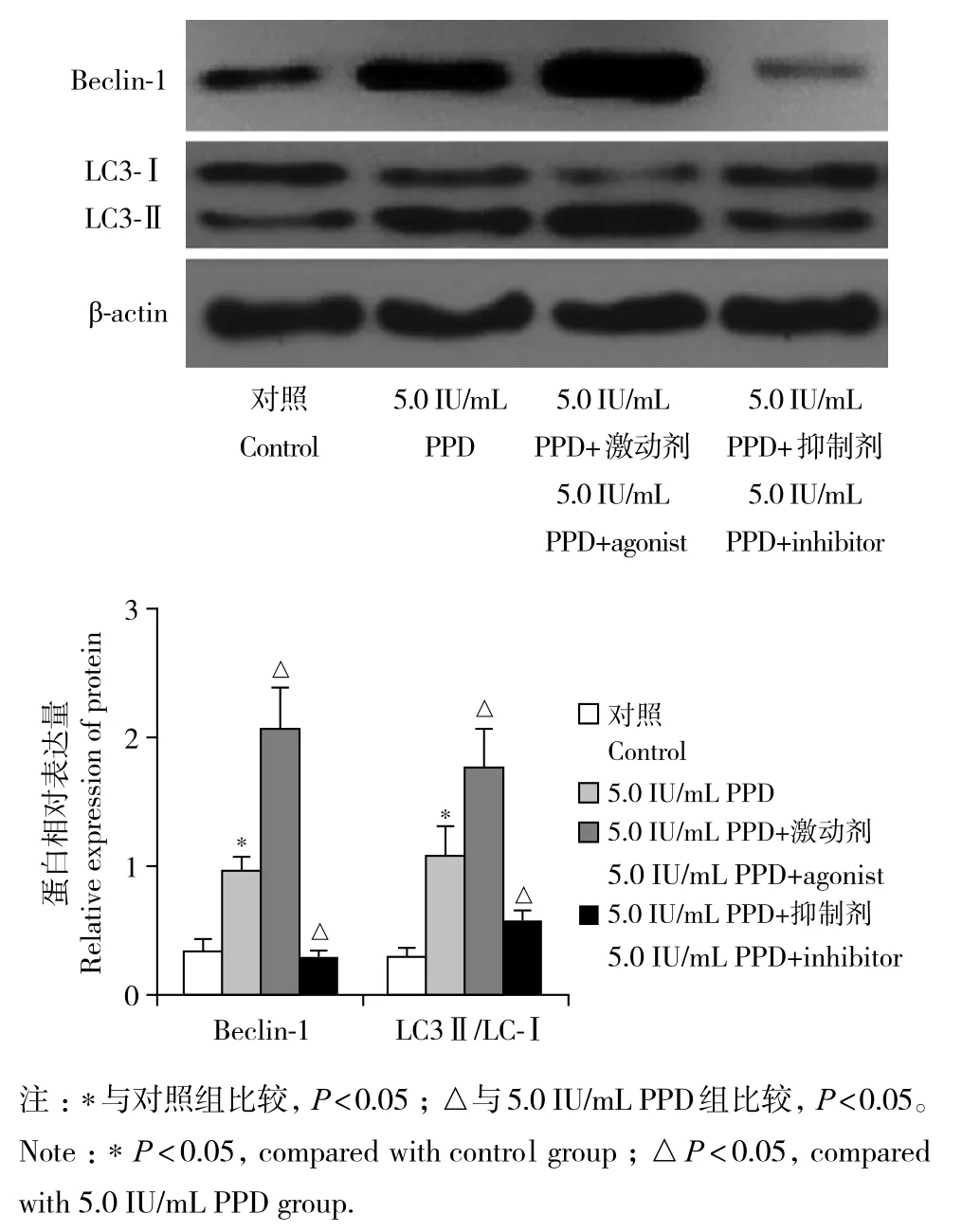
图7 蛋白质印迹法检测自噬激动剂及抑制剂对5.0 IU/mL PPD诱导后RAW264.7细胞中Beclin-1、LC3表达的影响Fig. 7 Effects of autophagy agonists and inhibitors on expression of Beclin-1 and LC3 in RAW264.7 cells after 5.0 IU/mL PPD induction detected by Western blotting
2.5 自噬激动剂及抑制剂对5.0 IU/mL PPD 诱导后RAW264.7细胞分化为破骨细胞的影响
与对照组比较,5.0 IU/mL PPD 组RAW264.7细胞分化为破骨细胞的数量及培养基中TRAP的活力增高,差异有统计学意义(P< 0.05,图8);与5.0 IU/mL PPD组比较,5.0 IU/mL PPD+ 激动剂组RAW264.7细胞分化为破骨细胞的数量及培养基中TRAP的活力增高,5.0 IU/mL PPD+ 抑制剂组RAW264.7细胞分化为破骨细胞的数量及培养基中TRAP的活力降低,差异均有统计学意义(P< 0.05,图8)。
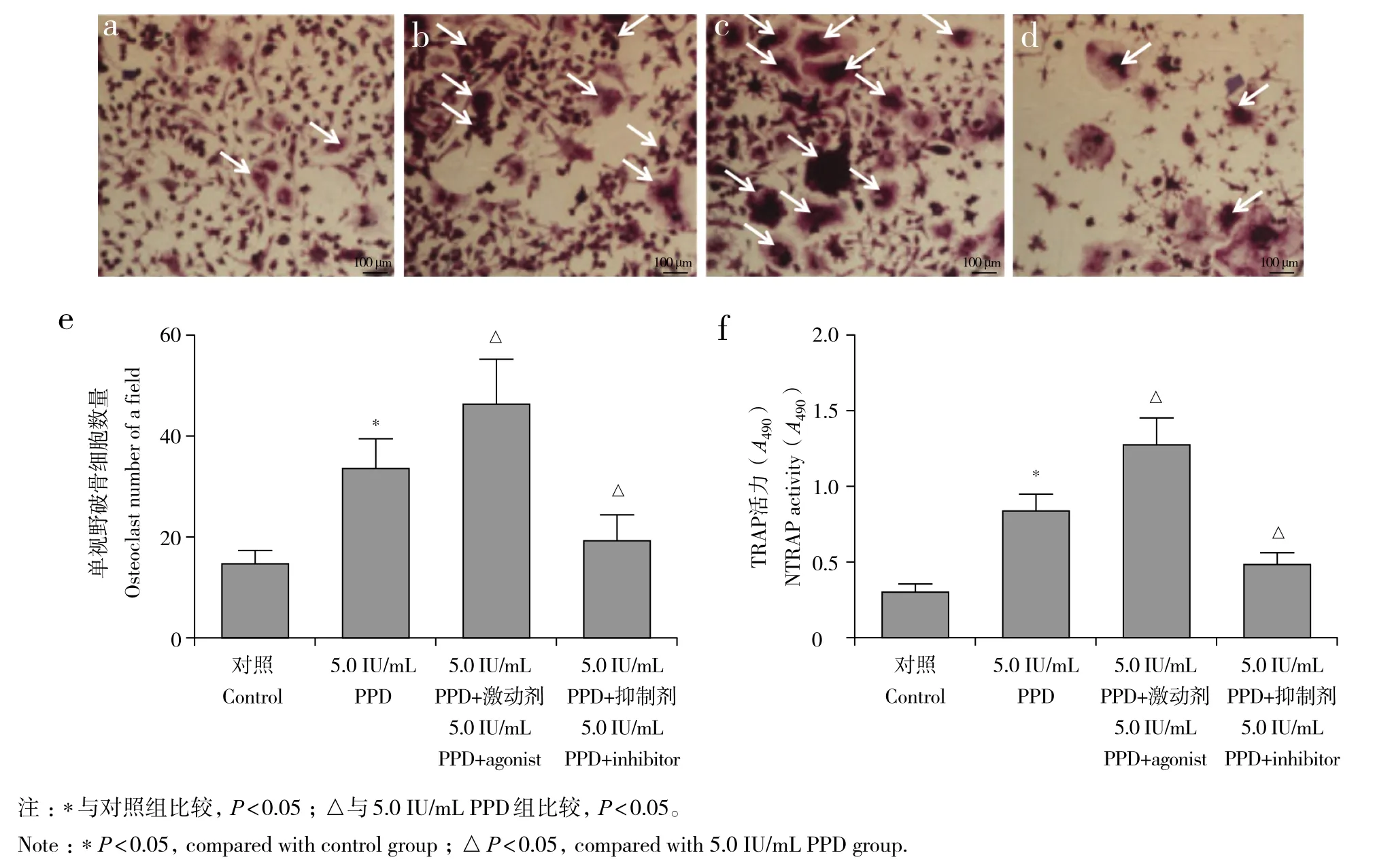
图8 自噬激动剂及抑制剂对5.0 IU/mL PPD诱导后RAW264.7细胞分化为破骨细胞的影响Fig. 8 Effect of autophagy agonist and inhibitor on differentiation of RAW264.7 cells after 5.0 IU/mL PPD induction into osteoclasts
3 讨 论
进行性的骨质破坏是脊柱结核的特征,主要累及椎体,可引起椎体塌陷、脊髓压迫,严重者出现脊柱后凸畸形或截瘫[10]。破骨细胞是体内唯一一种介导骨质吸收功能的终末细胞,在正常组织中含量极少,在病理条件下可由单核巨噬细胞分化而来。在脊柱结核发生过程中,破骨细胞与骨质破坏密切相关,多项研究发现PPD能够促进单核巨噬细胞向破骨细胞分化。梁思敏等[3,11]使用1.0 IU/mL和10.0 IU/mL PPD 诱导RAW264.7细胞形成破骨细胞,但10.0 IU/mL PPD 会影响RAW264.7细胞活力。本研究在此基础上对1.0 ~ 10.0 IU/mL 的浓度区间进行了细化,使用1.0、2.5、5.0、10.0 IU/mL的PPD对RAW264.7细胞进行诱导,发现仅10.0 IU/mL PPD 会使RAW264.7细胞活力降低,其余3种浓度PPD不影响RAW264.7细胞活性 ;1.0 IU/mL PPD 虽不影响细胞活性,但也未能促进RAW264.7细胞形成破骨细胞,而其余3种浓度PPD能够明显增加破骨细胞数量,具有促进RAW264.7细胞形成破骨细胞的作用。
目前,PPD诱导破骨细胞形成的作用已经受到广泛关注,但与之相关的分子机制尚不十分清楚。有研究[12-14]报道,结核分枝杆菌感染对单核巨噬细胞的自噬具有激活作用,自噬的激活可能有利于病原菌的清除,但过度自噬也可能引起组织损伤。本研究采用不同浓度PPD诱导RAW264.7细胞,结果发现1.0、2.5、5.0、10.0 IU/mL的PPD均能激活自噬,自噬小体数目及自噬基因Beclin-1、LC3-Ⅱ/LC-Ⅰ的表达均明显增加,与既往研究结果一致。
骨质疏松的多项研究[8-9,15-16]发现,自噬与骨代谢密切相关,成骨细胞及破骨细胞均受到自噬调控,当自噬发生抑制时,破骨细胞的分化明显受阻。为了阐明PPD激活自噬在RAW264.7细胞形成破骨细胞中的作用,本研究分别使用了自噬激动剂雷帕霉素和自噬抑制剂3-MA进行实验,在5.0 IU/mL PPD诱导RAW264.7细胞形成破骨细胞的过程中加用雷帕霉素或3-MA,结果发现,激活自噬能够增强PPD诱导破骨细胞形成的作用、抑制自噬能够削弱PPD诱导破骨细胞形成的作用,表明自噬在PPD诱导RAW264.7细胞形成破骨细胞的过程中起重要作用。
综上所述,PPD诱导破骨细胞形成的作用与自噬激活有关,这为今后研究脊柱结核发生过程中骨质破坏的发生机制提供了新思路,也为研究脊柱结核新的防治靶点提供了理论参考。但目前PPD激活自噬的机制尚不十分清楚,有研究[17-18]认为,mTOR、AMPK等经典的自噬调控通路在PPD激活自噬中起调控作用,也有研究[19-20]认为,miR-18a、miR-125b-5p等非编码RNA在PPD激活自噬中起调控作用,但具体机制仍有待今后更多的研究证实。
猜你喜欢
中国老年学杂志(2022年19期)2022-11-21
传染病信息(2022年3期)2022-07-15
传染病信息(2022年3期)2022-07-15
中国兽医学报(2022年4期)2022-06-17
安徽农业科学(2022年6期)2022-04-11
中国当代医药(2020年6期)2020-04-01
家庭医药(2020年2期)2020-03-17
家庭医学(2020年12期)2020-01-20
百科探秘·海底世界(2018年6期)2018-08-03
小哥白尼·趣味科学画报(2009年5期)2009-06-19
