
ATP6蛋白影响异沙叶蝉对小麦蓝矮植原体的传播效率
2021-12-08 01:48陆文静杨彦君吴云锋
昆虫学报 2021年11期
丁 磊,陆文静,2,杨彦君,马 欢,吴云锋,*
(1.西北农林科技大学植物保护学院,旱区作物逆境生物学国家重点实验室,农业部西北黄土高原作物有害生物综合治理重点实验室, 陕西杨凌 712100;2.黑龙江省农业科学院水稻研究所,黑龙江佳木斯 154026)
小麦蓝矮病是我国西北麦区一种重要植原体病害,对小麦危害严重(Wuetal.,2010)。该病害由异沙叶蝉Psammotettixalienus持久专化性传播,介体叶蝉传毒成为病害流行的中心环节(顾沛雯等,2007)。明确小麦蓝矮植原体(wheat blue dwarf phytoplasma,WBDp)虫传相关蛋白与何种叶蝉蛋白互作及叶蝉传播小麦蓝矮植原体的分子机制,探索植原体病害防治新途径成为当前面临的关键问题。
作者前期经过序列比对发现,pWBD-2-ORF4(WBDp第2个质粒编码的ORF4,P2-4)与泡桐丛枝植原体虫传蛋白pPaWBNy-1-ORF5的氨基酸序列(耿显胜,2013)一致性为79.14%(Chenetal.,2014)。随后进行虫传和嫁接对比实验,qRT-PCR检测表明传染初期叶蝉传播长春花植株中的P2-4表达量要明显高于嫁接接种WBDp的植株中的,推测P2-4参与WBDp介体传播。但如何影响没有研究报道。因此,本研究构建了异沙叶蝉分离泛素酵母双杂交膜系统cDNA文库,用P2-4蛋白从文库中筛选,获得ATP6等异沙叶蝉互作蛋白;利用RNAi技术沉默互作蛋白,明确互作蛋白在WBDp传播过程中的功能,对于阻断异沙叶蝉的传播过程和WBDp病害绿色防控具有重大意义。
1 材料与方法
1.1 供试材料
WBDp毒源、无毒介体异沙叶蝉由本实验室提供。异沙叶蝉饲养于小麦小偃6号幼苗上,人工气候室控制温度24℃,相对湿度20%~40%,光周期16L∶8D,每15 d更换一次小麦幼苗。NMY51酵母菌株、pNubG-Fe65,pTSU2-APP,pBT3STE,pPR3-N和pOST1-NubI载体质粒购自上海海科生物技术有限公司,T7 RiboMAXTMExpress RNAi System购自Promega公司。
1.2 异沙叶蝉分离泛素酵母双杂交膜系统cDNA文库构建
异沙叶蝉总RNA的提取参照Trizol®Reagent (Invitrogen)操作手册进行,分别用1.3%琼脂糖凝胶电泳和微量紫外分光光度计检测RNA质量和浓度。使用Oligotex mRNA Kits(Qiagen)分离纯化样本mRNA。参照SMART cDNA Library Construction方法(Zhuetal.,2001)合成ds cDNA,将cDNA纯化产物和pPR3-N同时用SfiⅠ酶切连接完成异沙叶蝉cDNA文库的构建,并检测文库质量和滴度。
1.3 P2-4诱饵载体的构建
根据P2-4基因(GenBank登录号:JX668988)序列设计带有SfiⅠ酶切位点的克隆引物对P2-4-pBT3STE-F/R(表1),使用CTAB法提取带WBDp小麦叶片总DNA,再以总DNA为模板扩增P2-4基因,通过同源重组将目的基因构建到酵母双杂交诱饵载体pBT3STE上,用引物P2-4-F/R(表1)和引物pBT3-F1/R1(表1)分别扩增目的片段无误后,测序验证。PCR反应体系(25 μL):DNA 模板2 μL,2×Taq Master Mix 12.5 μL,正反向引物(10 μmol/L)各 1 μL以及 ddH2O 8.5 μL。反应条件:94℃ 3 min;94℃ 30 s,51℃ 60 s,72℃ 90 s,30个循环;最终延伸 72℃ 10 min。
1.4 诱饵载体pBT3STE-P2-4筛选叶蝉cDNA文库
参照DUALhunter System操作方法(Möcklietal.,2008),将诱饵载体pBT3STE-P2-4和异沙叶蝉cDNA文库质粒共转化到NMY51酵母菌中,将转化产物涂布在DDO和QDO选择培养基平板上,28℃培养4~5 d。挑取单菌落重新在四缺培养基(QDO)培养基上培养并进行α-半乳糖苷酶检测,挑选阳性克隆进行PCR鉴定并测序,测序结果在NCBI BLAST比对分析。同时对筛选到的互作蛋白进行GO注释以及KEGG通路分析。
1.5 ATP6基因片段扩增
利用Primer Premier 5.0软件在ATP6基因(GenBank登录号:KX437742)功能区设计引物ATP6-F/R(表1)扩增ATP6基因片段。绿色荧光蛋白基因GFP(GenBank登录号:KF718991)片段通过特异性引物(表1)从本实验室保存的pCambia1302-GFP质粒上扩增。将ATP6和GFP片段纯化后分别与pMD-18T载体(TaKaRa)连接得到pMD-18T-ATP6和pMD-18T-GFP,转化大肠杆菌EscherichiacoliDH5α感受态细胞,鉴定出阳性克隆后提取质粒送测序验证。
1.6 RNAi沉默ATP6基因后异沙叶蝉传播WBDp能力的检测
使用引物ATP6-TF/R(表1)和GFP-TF/R(表1)从1.5节构建的重组载体pMD-18T-ATP6和pMD-18T-GFP中PCR扩增目的基因ATP6和GFP,反应体系和反应条件见1.3节。以PCR产物为模板合成dsRNA。利用Promega公司的T7 Ribomax Express RNAi System试剂盒合成ATP6和GFP的dsRNA,dsGFP用于对照组。将2 μL dsRNA (5 μg/μL)与20 μL液体人工饲料混匀后,在玻璃双通管内饲喂无毒异沙叶蝉3龄若虫2 d(Fuetal.,2001;田宏刚,2009),在饲喂结束后的第1,3,5,7,9和11天取样进行qRT-PCR检测异沙叶蝉3龄若虫体内ATP6的表达情况,引物序列见表1。PCR反应体系(20 μL):cDNA模板1 μL,UltraSYBR Mixture 10 μL,正反向引物(10 μmol/L)各0.5 μL以及 ddH2O 8 μL。反应条件:95℃ 10 min;95℃ 15 s,60℃退火/延伸 60 s,40个循环。生物学重复和技术重复各3次,每个生物学重复来自3头个体,内参基因为Actin,内参基因引物序列为qActin-F/R(表1)。参照 2-ΔΔCt方法计算基因的相对表达量变化(Livak and Schmittgen,2001),使用Microsoft Excel 2010对数据进行整理并作图。
转接无毒异沙叶蝉3龄若虫至含有dsRNA的人工饲料饲喂2 d后再饲喂WBDp 7 d,每株小麦转接3头异沙叶蝉若虫传毒7 d,培养生长14 d后PCR检测植株感病情况并分析沉默ATP6基因及对异沙叶蝉传播WBDp能力的影响。先提取小麦叶片总DNA,再以总DNA为模板使用引物P1/P7(表1)和引物R16F2n/R2(表1)进行巢式PCR检测小麦带WBDp的情况。PCR反应体系和反应条件见1.3节。之后统计dsATP6组和dsGFP组(每处理组实验重复3次,每次检测小麦30株)带WBDp的小麦植株数量,分别计算两组小麦带WBDp的百分率,即为异沙叶蝉对WBDp的传播率。
1.7 数据分析
基因的相对表达量和WBDp传播率著性检验方法采用SAS 9.2软件中独立样本T检验(P<0.01)进行,结果表示为3次重复的平均值±标准误。
2 结果
2.1 异沙叶蝉分离泛素酵母双杂交膜系统cDNA文库构建和质量检测
电泳结果显示提取的异沙叶蝉总RNA未降解,完整性较好,纯化出3.9 μg mRNA,条带清晰且分布均匀,符合建库需要(图1)。mRNA反转录产物ds cDNA经SfiⅠ酶切后连接pPR3-N文库载体,获得了以pPR3-N为载体的酵母双杂交膜系统cDNA文库,原始文库库容量约为1.25×107cfu。PCR检测显示插入片段平均长度大于1.2 kb,文库阳性率为100%(图2)。扩繁文库菌液并抽提质粒,得到文库质粒2.5 mg。
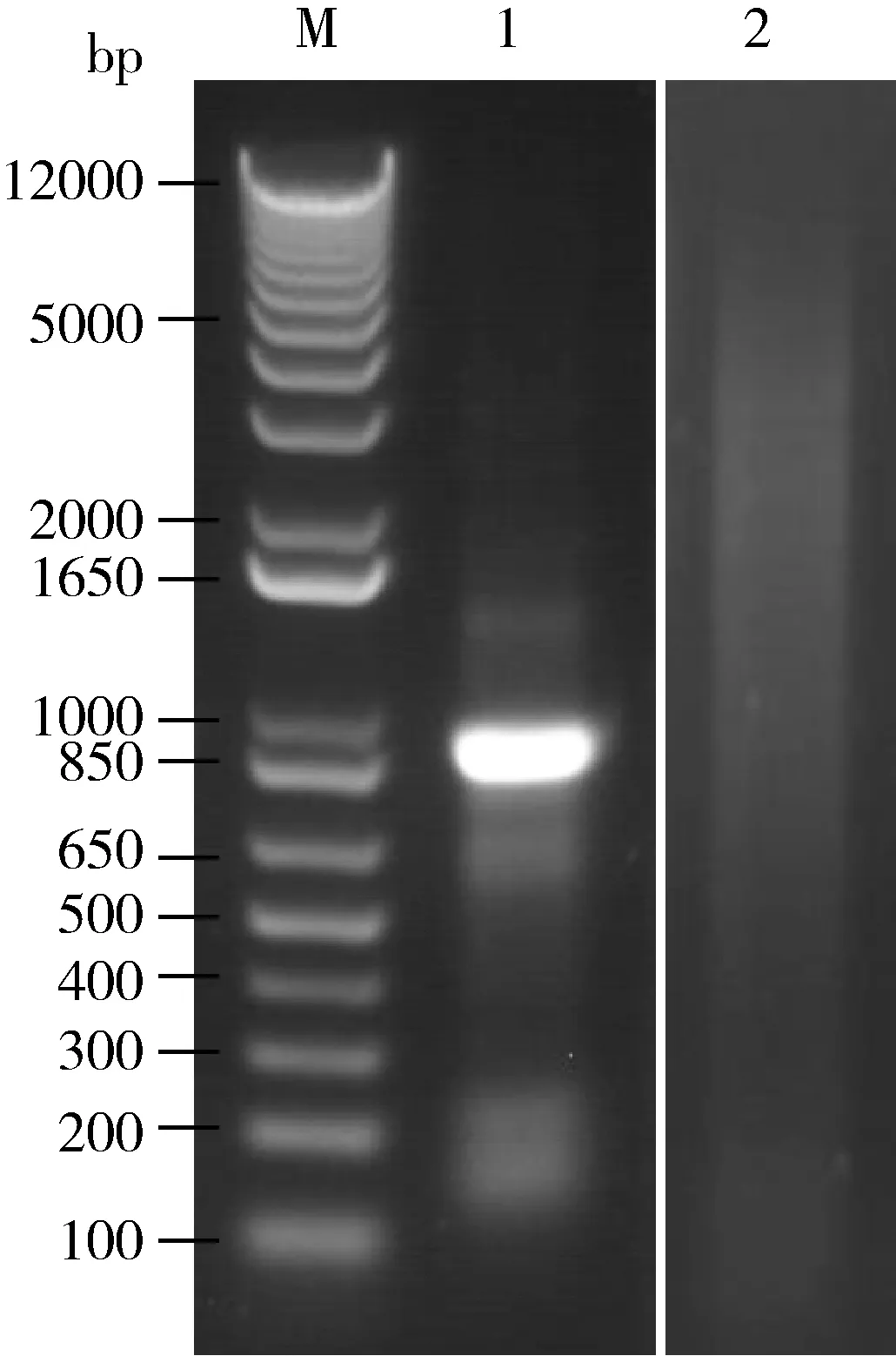
图1 异沙叶蝉总RNA和纯化mRNA电泳Fig.1 Electrophoresis of total RNA and purified mRNA isolated from Psammotettix alienusM:DL12000;1:总RNA Total RNA;2:纯化的mRNA Purified mRNA.

图2 异沙叶蝉cDNA文库插入片段的PCR检测Fig.2 Detection of inserted fragments of cDNA library of Psammotettix alienus by PCRM:DL12000;1-24:24个随机选取的克隆24 clones randomly selected.
2.2 P2-4杂交异沙叶蝉分离泛素酵母双杂交膜系统cDNA文库筛选互作蛋白
使用带酶切位点的P2-4-pBT3STE-F/R引物扩增目的基因,经DNA凝胶回收试剂盒回收后将P2-4同源重组法连接至pBT3STE载体,构建了诱饵蛋白载体pBT3STE-P2-4。使用P2-4-F/R和pBT3-F1/R1引物扩增分别得到489 bp和591 bp条带(图3),送测序验证重组质粒序列无误。

图3 小麦蓝矮植原体的P2-4基因及其重组质粒pBT3STE-P2-4的PCR电泳检测Fig.3 Gel electrophoresis of P2-4 gene of wheat blue dwarf phytoplasma and its recombinant plasmid pBT3STE-P2-4 detected by PCRM:DS5000;1:P2-4;2:pBT3STE-P2-4.
将诱饵载体pBT3STE-P2-4与异沙叶蝉文库质粒共转化酵母NMY51感受态细胞,在DDO和QDO培养基上进行培养。结果获得总菌落数为1.57×106cfu,转化效率为5.39×104cfu/μg。提取酵母菌阳性克隆质粒并转入到大肠杆菌中进行测序,将测序结果在BLAST分析后,获得ATP synthase F0subunit 6(ATP6)等8种候选蛋白(表2),回复验证和α-半乳糖苷酶检测结果显示P2-4与以上8种蛋白互作(图4),其中与ATP6为强互作。

图4 异沙叶蝉cDNA文库中与WBDp的P2-4互作蛋白阳性克隆的α-半乳糖苷酶检测Fig.4 Detection of α-galactosidase of positive clones interacting with P2-4 of WBDp in the cDNA library of Psammotettix alienusP:阳性Positive;N:阴性Negative;1-8:阳性克隆,对应的蛋白信息见表2 Positive clones,their corresponding proteins as in Table 2.

表2 从异沙叶蝉cDNA文库筛选出与小麦蓝矮植原体的P2-4互作的蛋白Table 2 Putative proteins screened in the cDNA library of Psammotettix alienus interacting with P2-4 of wheat blue dwarf phytoplasma
对筛选到的互作蛋白进行GO注释,结果显示上述蛋白参与的生物学进程包括ATP合成耦合质子运输(ATP synthesis coupled proton transport)、三磷酸鸟苷生物合成过程(GTP biosynthetic process)、凋亡过程(apoptotic process)、调节肌肉细胞细胞内稳态(muscle cell cellular homeostasis)、消化过程(digestive process)、运输(transport)及细胞质翻译(cytoplasmic translation)等;分子功能包括质子跨膜转运体活性(proton transmembrane transporter activity)、核糖体结构成分(structural constituent of ribosome)、催化活性(catalytic activity)、细胞色素C的凋亡释放(apoptotic release of cytochrome c)、肌动蛋白与金属离子结合(actin and metal ion binding)、半胱氨酸型肽酶活性(cysteine-type peptidase activity)及脂质结合(lipid binding)等;细胞组成包括线粒体(mitochondrion)、细胞膜(membrane)、细胞质溶胶(cytosol)、线粒体内膜(mitochondrion inner membrane)和细胞骨架(cytoskeleton)等(图5)。参考KEGG数据库,上述蛋白参与异沙叶蝉体内能量代谢、跨膜运动和其他代谢调控过程。

图5 异沙叶蝉cDNA文库中与小麦蓝矮植原体的P2-4互作蛋白的GO注释Fig.5 GO annotation of proteins in cDNA library of Psammotettix alienus interacting with P2-4 of wheat blue dwarf phytoplasmaA:分子功能 Molecular function;B:生物学进程 Biological process;C:细胞组成 Cellular component.
2.3 ATP6片段扩增和dsRNA的合成
经基因扩增,得到525 bp的ATP6和284 bp的GFP片段条带(图6),分别纯化后插入pMD-18T载体,得到pMD-18T-ATP6和pMD-18T-GFP,以从克隆载体中扩增的目的基因的PCR产物为模板合成dsRNA,经过洗涤和纯化,获得ATP6的dsRNA和GFP的dsRNA。
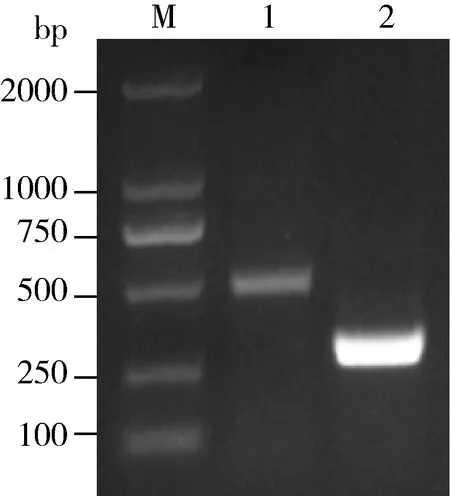
图6 ATP6和GFP扩增片段的电泳检测Fig.6 Gel electrophoresis of ATP6 and GFP fragmentsM:DS2000;1:ATP6;2:GFP.
2.4 沉默ATP6对异沙叶蝉传毒能力的影响
用ATP6和GFP的dsRNA 分别饲喂无毒异沙叶蝉3龄若虫后,在6个时间点取样进行qRT-PCR检测,结果表明饲喂ATP6的dsRNA第1-11天异沙叶蝉体内ATP6的表达量降低,仅为对照组(dsGFP)的30%~40%(P<0.01),第5天下降最明显(图7),表明饲喂dsATP6能够降低异沙叶蝉若虫体内ATP6 mRNA水平。用饲喂dsRNA 2 d后的异沙叶蝉3龄若虫进行传毒能力测定,结果表明dsATP6饲喂组叶蝉的WBDp传播率比对照组(dsGFP饲喂组)下降了23.33%±3.33%(图8)。说明筛选到的ATP6蛋白对异沙叶蝉传播小麦蓝矮植原体的能力有一定影响,是一种重要的传毒相关蛋白。

图7 RNAi干扰ATP6对健康异沙叶蝉3龄若虫中ATP6的相对表达量的影响Fig.7 Effect of RNAi of ATP6 on the relative expression level of ATP6 in the 3rd instar nymphs of healthy Psammotettix alienus图中数据为平均值±标准误;柱上星号表示差异显著(P<0.01,T检验)。Data in the figure are mean±SE.Asterisk above bars indicates significant differences (P<0.01,T-test).下同The same below.
3 讨论
本研究运用SMART技术构建了高质量的异沙叶蝉分离泛素酵母双杂交膜系统cDNA文库,并以P2-4蛋白为诱饵,筛选到ATP合成酶等8种互作蛋白(表1),可能与WBDp传播相关。利用RNAi技术沉默ATP6基因表达后,异沙叶蝉3龄若虫传播WBDp的效率明显降低了23.33%(图8),说明异沙叶蝉ATP6蛋白影响了WBDp的持久专化性传播。从异沙叶蝉体内鉴定出的ATP6互作蛋白为研究WBDp与昆虫介体互作机制和植原体病害的田间防治提供了理论基础。
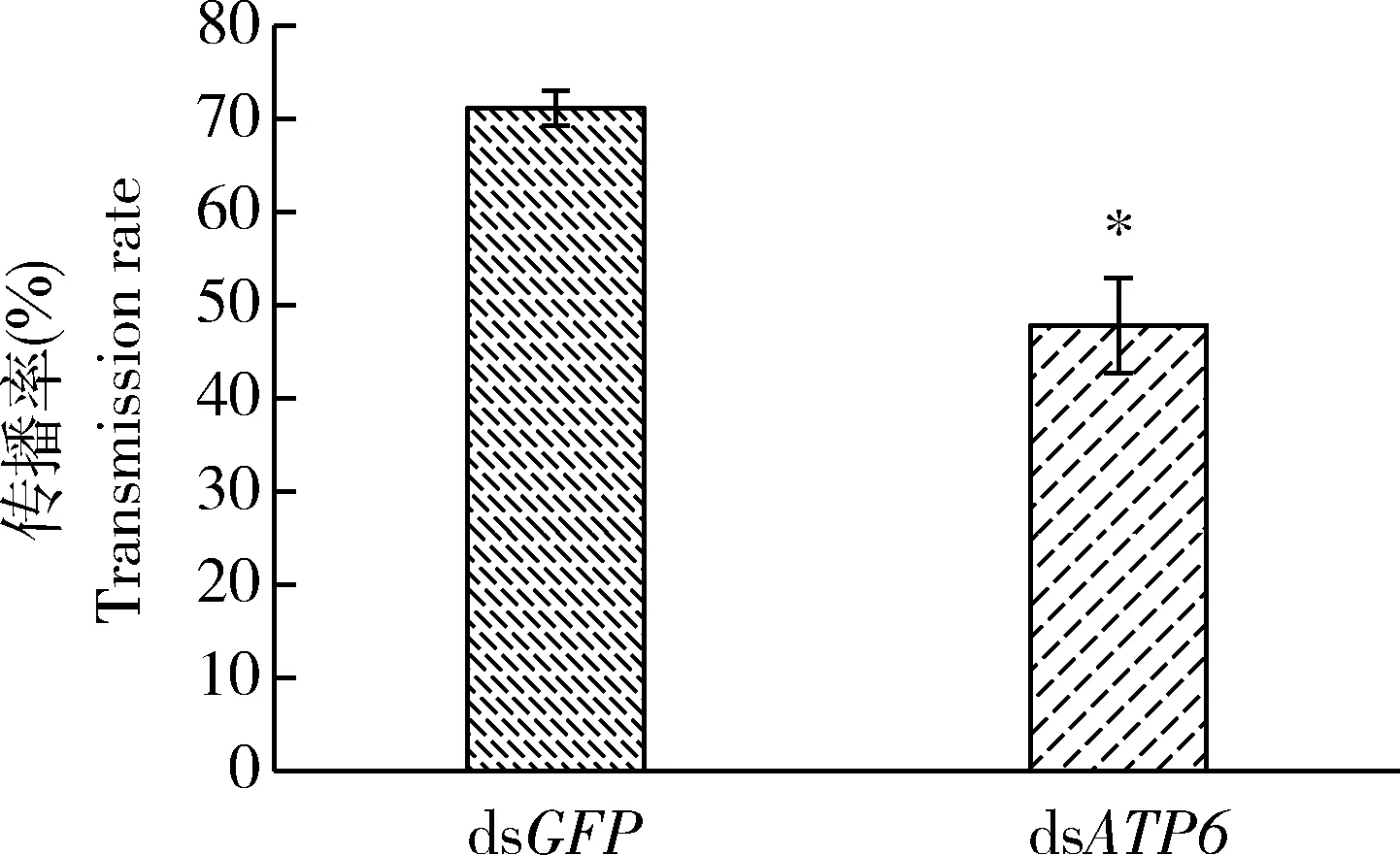
图8 ATP6 RNAi对异沙叶蝉3龄若虫的WBDp传播率的影响Fig.8 Effect of RNAi of ATP6 on the WBDp transmission rate by the 3rd instar nymphs of Psammotettix alienus将无毒异沙叶蝉3龄若虫饲喂含有dsRNA的人工饲料2 d后再饲喂WBDp 7 d,之后每株小麦转接3头异沙叶蝉若虫传毒7 d,接虫14 d后PCR检测小麦植株带WBDp的情况;每处理组实验重复3次,每次检测30株小麦。The 3rd instar nymphs of healthy P. alienus were fed with WBDp for 7 d after fed with the artificial diet containing dsRNA for 2 d,and then three nymphs were transferred to a wheat plant for 7 d.The presence of WBDp in wheat plants were detected at 14 d after nymph infestation.Three repeat experiments were performed for each treatment,and a total of 30 wheat plants were examined for each repeat.
本实验利用P2-4蛋白筛选异沙叶蝉酵母文库,共获得8种可能与WBD植原体传播相关的异沙叶蝉蛋白。其中,ATP6的克隆数为7个,远多于其他7种互作蛋白的克隆数。已有研究表明ATP合成酶通过两个结构域F1和F0相互作用与细胞膜结合催化ATP的合成(Boyer,1997),为各种细胞活动提供能量。由于植原体本身缺乏ATP合成途径(Oshimaetal.,2004;Jietal.,2010),在一定程度上依赖介体昆虫体内的ATP完成运输和繁殖(Christensenetal.,2005;Hogenhoutetal.,2008)。我们推测异沙叶蝉ATP6蛋白参与WBDp的传播并首选ATP6蛋白进行后续验证。利用RNAi技术沉默ATP6后,异沙叶蝉传播WBDp的效率明显下降(图8)。表明异沙叶蝉ATP6蛋白是参与WBDp传播的关键蛋白。太平洋白虾LitopenaeusvannameiATP synthase β与白斑综合症病毒(white spot syndrome virus,WSSV)结合,并在宿主侵袭过程中发挥作用(Liangetal.,2010)。同时作为病毒受体,介导病毒颗粒进入宿主细胞(Fongsaranetal.,2014;Liangetal.,2015)。ATP synthase β也与植原体的免疫膜蛋白互作并参与植原体介体传播(Galettoetal.,2011)。本研究中WBDp的P2-4蛋白和ATP6存在特异性互作,推测WBDp在进入介体消化系统的过程中,利用叶蝉体内ATP完成运输和繁殖。本研究结合酵母双杂交和RNAi技术,鉴定出一种参与WBDp传播的关键蛋白ATP6,但是对于P2-4与ATP6蛋白的结合方式及互作机理仍不明确。在接下来的研究中,我们将利用GST-pull down和免疫共沉淀(co-immunoprecipitation,Co-IP)等方法进一步验证二者的互作关系,并结合免疫荧光(immunofluorescence,IF)等技术揭示异沙叶蝉传播WBDp的分子机理。
RNAi技术已经有20余年的发展历史,而且在害虫控制中显示出重要价值和巨大的应用潜力(Zhangetal.,2017)。RNAi不仅是基因功能鉴定的有效分子工具,也是开发害虫和病害控制的新途径,对环境影响小,目标特异性强,已经广泛应用于害虫防治(Gu and Knipple,2013;Lietal.,2013;Jogaetal.,2016;Kanakala and Ghanim,2016)。本研究通过饲喂法将ATP6 dsRNA导入异沙叶蝉体内后沉默效果明显(图7),表明ATP6在结合WBDp进入异沙叶蝉体内发挥重要作用。同时ATP6也有望成为降低异沙叶蝉对小麦蓝矮植原体的传播效率的有效靶标,对小麦蓝矮植原体病害的防治具有重要意义。
致谢本研究异沙叶蝉RNAi实验得到西北农林科技大学植物保护学院田宏刚老师的指导和帮助,特此致谢。
猜你喜欢
湘潮(上半月)(2022年7期)2022-12-06
今日农业(2022年16期)2022-11-09
中国农学通报(2022年12期)2022-06-01
中国糖料(2022年2期)2022-04-06
今日农业(2021年8期)2021-11-28
中国种业(2021年11期)2021-11-25
金桥(2021年10期)2021-11-05
今日农业(2021年13期)2021-08-14
教学考试(高考生物)(2021年2期)2021-05-31
猪业科学(2021年3期)2021-05-21
