
猪RNase L与PRRSV nsp4缺失突变体的构建及其互作研究
2021-11-11 08:40于莹李均同丛晓燕齐静刘思当郑乾坤孙文博王爱国郑龙祥吴香菊杜以军单虎
畜牧与兽医 2021年11期
于莹,李均同,丛晓燕,齐静,刘思当,郑乾坤,孙文博,王爱国,郑龙祥,吴香菊,*,杜以军*,单虎
(1. 青岛农业大学动物医学院,山东 青岛 266000;2. 山东省畜禽疫病防治与繁育重点实验室/山东省农业科学院畜牧兽医研究所,山东 济南 250100;3. 山东农业大学动物科技学院,山东 泰安 271018;4. 得利斯集团有限公司,山东 诸城 262216)
猪繁殖与呼吸综合征(procine reproductive and respiratory syndrome, PRRS)是由猪繁殖与呼吸综合征病毒(porcine reproductive and respiratory syndrome virus, PRRSV)引起的一种高度接触性传染病,通过呼吸道、胎盘、生殖道传播,通常表现为各年龄段猪的呼吸困难以及母猪的生殖衰竭,如晚期流产、死产和木乃伊化,致死率可高达80%~100%[1],目前仍然是世界范围内养猪业的主要威胁。PRRS于20世纪80年代末,几乎同时在北美和欧洲出现,然后在几年内迅速传播到世界其他地区。在1991年和1992年分别于荷兰和美国出现[2]。1996年在爱荷华州和美国其他州暴发了“急性PRRS”[3]。2006年高致病性PRRSV(HP-PRRSV)在中国以未知的方式大规模暴发,传播到了10多个省(自治市或自治区),感染了超过200万头猪,造成约40万死亡病例。该病具有唯一的感染宿主,而且一般伴随混合与继发性感染。
非结构蛋白nsp4是PRRSV编码的一种蛋白酶,可以下调免疫反应[4]。通过切割受体或接头分子IPS-1(interferon-β promoter stimulator 1)、NEMO(NF-κB essential modulator)或IκB的催化亚基IΚΚα显著抑制I型干扰素的产生,对IFN-β启动子的激活具有抑制作用。研究表明nsp4激活caspase-3、-8和-9诱导细胞凋亡,并且该诱导凋亡活性依赖于nsp4的丝氨酸蛋白酶活性。PRRSV nsp4的晶体结构[5]揭示了该蛋白由3个结构域组成,包括2个类糜蛋白酶β结构域和1个额外的C-末端α/β结构域,分别位于1~69 aa结构域Ⅰ,89~153 aa结构域II和157~199 aa结构域Ⅲ[6],其中结构域Ⅰ和Ⅱ形成典型的糜蛋白酶样两管折叠。C-末端结构域Ⅲ对于蛋白水解活性是必不可少的,可能参与微调复制酶多蛋白水解[7]。nsp4的活性位点位于结构域Ⅰ和Ⅱ之间,包含典型的催化三联体His39、Asp64和Ser118[8]。
核糖核酸内切酶L(RNase L)是一种由干扰素诱导产生的抗病毒蛋白(antiviral protein, AVP),参与机体内的特异性、非特异性免疫。RNase L结构包含3个结构域:N-端锚定蛋白重复区、蛋白激酶同源区、C-端核糖核酸酶区[9]。活化的RNase L切割单链RNA,包括病毒RNA和细胞RNA[10]。2-5A依赖的RNase L可以降解核糖体中的rRNA[11]。RNase L还可以诱导细胞凋亡等。本课题组前期探究到sRNase L具有抗PRRSV活性并筛选到sRNase L与PRRSV nsp4互作[12-13],Silverman等[14]研究发现R462Q的突变也终止了RNase L与2-5A结合区域的翻译,抑制了2-5A对RNase L的激活。Dong等[15]证明RNase L突变体W632A、D661A、R667A和H672A缺乏核糖核酸酶活性。Nakanishi等[16]试验证明Tyr712和Phe716对RNA的结合和切割功能起到重要的作用。本研究将分别构建sRNase L突变体、缺失体及nsp4的截短体,通过免疫共沉淀试验筛选两者的互作区段及位点,为深入探究sRNase L与nsp4互作抗PRRSV机制奠定基础,并为PRRSV感染的防控提供新的思路和方法。
1 材料与方法
1.1 材料
人胚胎肾上皮细胞(HEK-293T)、pXJ41-Flag-sRNase L、pXJ41-HA-nsp4和空载体pXJ41由本实验室保存。
1.2 主要试剂
DNA Marker DL5000、DNA Marker DL2000、Prime STAR HS DNA Ploymerase等购自TaKaRa宝生物工程(大连)有限公司;pfu Ultra High Fidelity DNA Ploymerase购自Stratagene公司;限制性内切酶HindⅢ、KpnⅠ、DpnⅠ、EcoRⅠ、XhoⅠ及T4 DNA Ligase购自Fermentas公司;质粒小提试剂盒、DNA回收试剂盒购自北京天根生物工程有限公司;WesternBright Sirius化学发光检测试剂盒购自Advansta公司;细胞裂解液、Western blot一抗稀释液购自碧云天公司;转染试剂Lipofectamine®3000购自Invitrogen公司;NC膜购自Bio-RAD公司;青链霉素混合液、Flag-beads、PMSF、Tween-20、cocktail购自Sigma公司;Opti-MEM、高糖型DMEM细胞培养基、胰蛋白酶购自Gibco公司;辣根过氧化物酶(HRP)标记羊抗鼠抗体、辣根过氧化物酶(HRP)标记羊抗兔抗体购自武汉博士德公司;DH5α感受态细胞购自北京全式金生物技术有限公司;血清购自BI公司;β-actin抗体购自Santa Cruz公司;兔源HA抗体、鼠源HA抗体、兔源Flag抗体、鼠源Flag抗体购自Sigma公司。
1.3 引物设计
根据文献报道,结合载体pXJ41的酶切位点,设计了3对含HA标签序列扩增nsp4 3个结构域的特异性引物;4对含Flag标签序列扩增RNase L突变体的特异性引物;1对含Flag标签序列扩增RNase L缺失体的特异性引物;以实验室保存的HA-nsp4、Flag-RNase L为模板,进行PCR扩增,引物送北京擎科生物技术有限公司合成,引物信息如表1所示。

表1 PCR引物及序列
1.4 pXJ41-Flag-sRNase L突变体的PCR扩增
以pXJ41-Flag-sRNase L质粒为模板,用Flag-sRNase L突变体上下游引物进行PCR扩增,反应体系为25 μL:pXJ41-Flag-sRNase L 100 ng,dNTP 1.5 μL,10×pfu buffer 2.5 μL,Flag-sRNase突变体上下游引物各0.7 μL,pfu Ultra High Fidelity DNA Ploymerase 0.5 μL,ddH2O补足25 μL。PCR反应条件为95 ℃预变性2 min;95 ℃变性20 s,58 ℃退火20 s,72 ℃延伸7 min,25个循环;72 ℃延伸7 min。PCR产物进行1.0%琼脂糖凝胶电泳。
1.5 pXJ41-Flag-sRNase L突变体重组质粒的构建及鉴定
用DpnⅠ酶切pXJ41-Flag-sRNase L突变体PCR扩增产物,将酶切产物转化到DH5α感受态细胞中,37 ℃过夜培养16 h,挑取单菌落接种于含有氨苄西林抗性的LB培养基中,37 ℃,200 r/min过夜培养,第2天提取质粒,送北京擎科生物科技有限公司测序。
1.6 缺失体Flag-sRNase L 1~586aa的PCR扩增
以pXJ41-Flag-sRNase L为模板,以Flag-sRNase L 1~586 aa-Fwd和Flag-sRNase L 1~586 aa-Rev作为上下游引物,PCR扩增缺失体Flag-sRNase L1~586 aa基因,PCR反应体系为25 μL:pXJ41-Flag-sRNase L 100 ng,dNTP 2 μL,5×Prime STAR Buffer 5 μL,Flag-sRNase L 1~586 aa-Fwd和Flag-sRNase L 1~586 aa-Rev各0.7 μL,Prime STAR HS DNA Ploymerase(2.5 U/μL)0.25 μL,ddH2O补足25 μL。PCR反应条件为98 ℃预变性2 min;98 ℃变性10 s,57 ℃退火30 s,72 ℃延伸3 min,30个循环;72 ℃延伸7 min。PCR产物进行1.0%琼脂糖凝胶电泳。
1.7 HA-nsp4截短体的PCR扩增
以pXJ41-HA-nsp4为模板,分别以HA-nsp4-D1/D2/D3-Fwd和HA-nsp4-D1/D2/D3-Rev作为上下游引物,PCR扩增HA-nsp4截短体基因,PCR反应体系为25 μL:pXJ41-HA-nsp4 100 ng,dNTP 2 μL,5×Prime STAR Buffer 5 μL,HA-nsp4-D-Fwd和HA-nsp4-D-Rev各0.7 μL,Prim STAR HS DNA Ploymerase(2.5 U/μL)0.25 μL,ddH2O补足25 μL。PCR反应条件为98 ℃预变性2 min;98 ℃变性10 s,57 ℃退火30 s,72 ℃延伸2 min,30个循环;72 ℃延伸7 min。PCR产物进行1.0%琼脂糖凝胶电泳。
1.8 重组质粒的构建及鉴定
用KpnⅠ和Hind Ⅲ分别双酶切缺失体Flag-sRNase L 1~586aa、HA-nsp4截短体基因和pXJ41载体的胶回收产物,分别将缺失体Flag-sRNase L 1~586aa基因和HA-nsp4截短体基因与pXJ41载体用T4 DNA Ligase 16 h过夜连接。次日,将连接产物转化到DH5α感受态细胞中,37 ℃过夜培养,挑取单菌落接种于含有氨苄西林的LB固体培养基中,37 ℃,200 r/min过夜培养,第2天提取质粒,进行KpnⅠ和Hind Ⅲ双酶切鉴定。鉴定为阳性的质粒送北京擎科生物科技有限公司测序。
1.9 转染
HEK-293T细胞在6 cm圆盘中进行培养,待细胞密度达到104个/mL时,利用Lipofectamine®3000转染试剂将2.5 μg的HA-nsp4分别与的2.5 μg的pXJ41、pXJ41-Flag-sRNase L、pXJ41-Flag-sRNase L R460A、pXJ41-Flag-sRNase L R675A、pXJ41-Flag-sRNase L Y720A、pXJ41-Flag-sRNase L F724A、缺失体pXJ41-Flag-sRNase L 1~586 aa按Lipofectamine®3000说明书共转染到细胞中;将2.5 μg的pXJ41-Flag-sRNase L分别与2.5 μg的pXJ41、pXJ41-HA-nsp4-D1、pXJ41-HA-nsp4-D2、pXJ41-HA-nsp4-D3按Lipofectamine®3000说明书共转染到HEK-293T细胞中。
1.10 免疫共沉淀(Co-IP)及Western blot
转染24 h后,将6 cm圆盘置冰上,弃上清,用 PBS洗2遍后加500 μL细胞裂解液裂解细胞,20 min后将样品收集到1.5 mL EP管中,4 ℃ 12 000 r/min离心10 min,取60 μL上清与上样缓冲液混合煮沸10 min标记为input样品,用于鉴定蛋白表达情况。剩余上清加入Flag-beads,4 ℃旋转孵育4 h,孵育结束后,4 500 r/min离心2 min弃上清,用1 mL细胞裂解液重悬,重复洗5遍,弃上清后与上样缓冲液混合煮沸15 min标记为免疫共沉淀样品(IP样品),用于检测互作情况。
将样品进行SDS-PAGE并转印到NC膜上。用5%脱脂奶粉室温封闭2 h,IP样品分别用一抗稀释液稀释的兔抗HA、兔抗Flag作为一抗;input样品分别用鼠抗HA、鼠抗Flag、鼠抗β-actin抗体作为一抗,4 ℃下过夜孵育,二抗室温下孵育1 h,用化学发光显色试剂盒WesternBright Sirius进行显色,BIO-RAD凝胶成像仪进行曝光。
2 结果与分析
2.1 目的基因的扩增
通过PCR扩增获得pXJ41-Flag-sRNase L R460A、pXJ41-Flag-sRNase L R675A、pXJ41-Flag-sRNase L Y720A、pXJ41-Flag-sRNase L F724A、缺失体Flag-sRNase L 1~586 aa、HA-nsp4-D1、HA-nsp4-D2、HA-nsp4-D3基因。扩增产物经过1.0%琼脂糖凝胶电泳检测到大小为6 525 bp(pXJ41-Flag-sRNase L)(图1A)、6 525 bp(pXJ41-Flag-sRNase L突变体)(图1A)、2 256 bp(Flag-sRNase L)(图1B)、1 782 bp(缺失体Flag-sRNase L 1~586aa)(图1B)、639 bp(HA-nsp4)(图1C)、267 bp(HA-nsp4-D1)(图1C)、317 bp(HA-nsp4-D2)(图1C)、177 bp(HA-nsp4-D3)(图1C)的特异性扩增片段,与预期片段大小相符。

A. pXJ41-Flag-sRNase L、pXJ41-Flag-sRNase L R460A、pXJ41-Flag-sRNase L R675A、pXJ41-Flag-sRNase L Y720A、pXJ41-Flag-sRNase L F724A基因的PCR扩增产物;B. Flag-sRNase L、缺失体Flag-sRNase L 1~586 aa基因的PCR扩增产物;C:HA-nsp4、HA-nsp4-D1、HA-nsp4-D2、HA-nsp4-D3基因的PCR扩增产物
2.2 pXJ41-Flag-sRNase L突变体质粒构建及鉴定
重组质粒pXJ41-Flag-sRNase L各突变体测序结果表明质粒pXJ41-Flag-sRNase L R460A、pXJ41-Flag-sRNase L R675A、pXJ41-Flag-sRNase L Y720A、pXJ41-Flag-sRNase L F724A构建成功(图2)。

A. pXJ41-Flag-sRNase L R460A;B. pXJ41-Flag-sRNase L R675A;C. pXJ41-Flag-sRNase L Y720A;D. pXJ41-Flag-sRNase L F724A
2.3 pXJ41-HA-nsp4截短体质粒构建及鉴定
重组质粒pXJ41-HA-nsp4-D1、pXJ41-HA-nsp4-D2、pXJ41-HA-nsp4-D3经EcoRⅠ/XhoⅠ双酶切后得到预期大小片段(图3A),测序结果表明质粒pXJ41-HA-nsp4-D1、pXJ41-HA-nsp4-D2、pXJ41-HA-nsp4-D3构建成功。
2.4 缺失体pXJ41-Flag-sRNase L 1-586aa质粒构建及鉴定
重组质粒缺失体pXJ41-Flag-sRNase L 1~586 aa经KpnⅠ/Hind Ⅲ双酶切后得到预期大小片段(图3B),测序结果表明质粒缺失体pXJ41-Flag-sRNase L 1~586 aa构建成功。
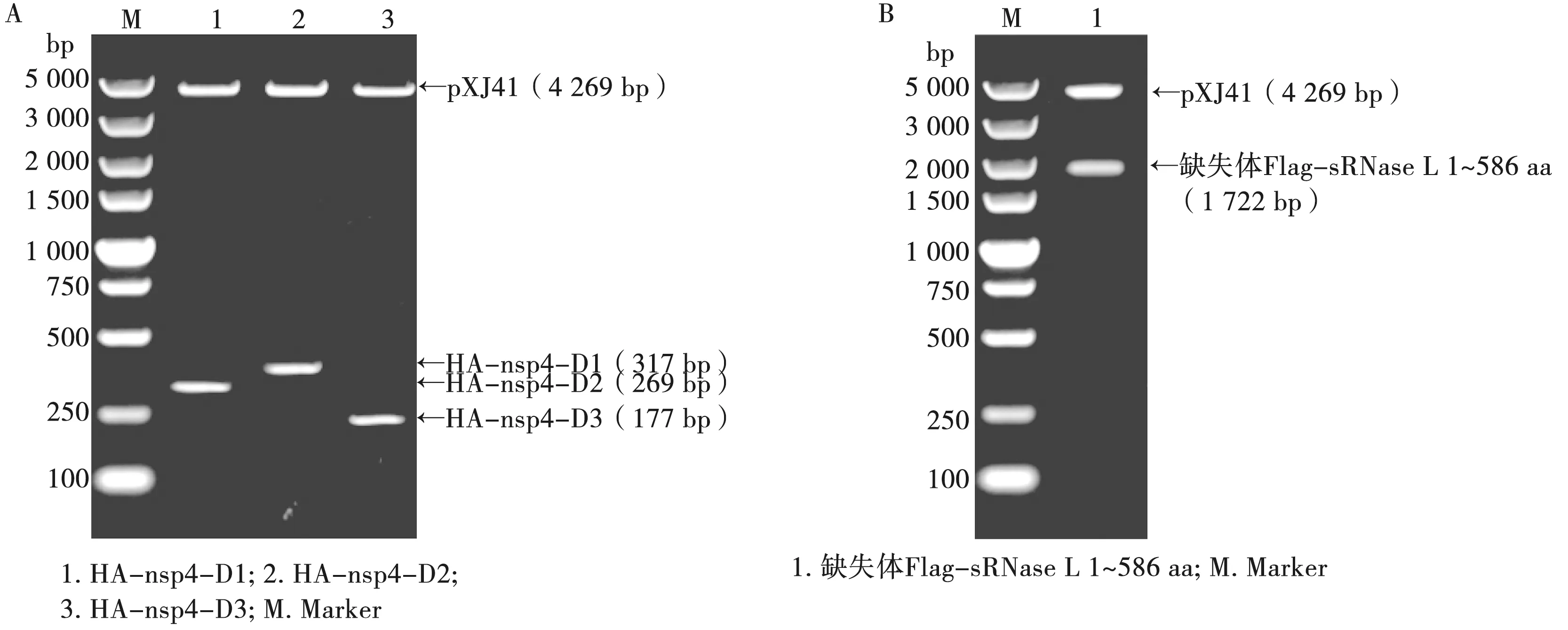
A. pXJ41-HA-nsp4-D1、pXJ41-HA-nsp4-D2、pXJ41-HA-nsp4-D3双酶切鉴定;B. 缺失体pXJ41-Flag-sRNase L 1~586 aa双酶切鉴定
2.5 免疫共沉淀筛选与nsp4互作的sRNase L位点及区段
pXJ41-HA-nsp4分别与pXJ41-Flag-sRNase L R460A、pXJ41-Flag-sRNase L R675A、pXJ41-Flag-sRNase L Y720A、pXJ41-Flag-sRNase L F724A、缺失体pXJ41-Flag-sRNase L 1~586aa、pXJ41-Flag-sRNase L、pXJ41共转染到HEK-293T细胞中,pXJ41-Flag-sRNase L R460A、pXJ41-Flag-sRNase L R675A、pXJ41-Flag-sRNase L Y720A、pXJ41-Flag-sRNase L F724A、缺失体pXJ41-Flag-sRNase L 1~586aa、pXJ41-Flag-sRNase L以及pXJ41-HA-nsp4质粒均成功表达;免疫共沉淀结果显示, pXJ41-Flag-sRNase L R460A、pXJ41-Flag-sRNase L R675A、pXJ41-Flag-sRNase L Y720A、pXJ41-Flag-sRNase L F724A、pXJ41-Flag-sRNase L分别与pXJ41-HA-nsp4发生了相互作用,但缺失体pXJ41-Flag-sRNase L 1~586aa未与nsp4发生互作,表明sRNase L第3结构域(587~744aa)为与nsp4互作区段,但不包括第675位精氨酸、第720位酪氨酸、第724位苯丙氨酸。见图4。

A. IP样品用兔源HA抗体做Western blot检测nsp4互作条带;B. IP样品用兔源Flag抗体做Western blot检测sRNase L条带;C. Input样品直接做Western blot检测sRNase L、sRNase L突变体、缺失体的表达情况;D. Input样品直接做Western blot检测nsp4的表达情况
2.6 免疫共沉淀筛选与sRNase L互作的nsp4区段
将pXJ41-Flag-sRNase L分别与pXJ41、pXJ41-HA-nsp4-D1、pXJ41-HA-nsp4-D2、pXJ41-HA-nsp4-D3共转染到HEK-293T细胞中。pXJ41-HA-nsp4-D1、pXJ41-HA-nsp4-D2、pXJ41-HA-nsp4-D3以及pXJ41-Flag-sRNase L质粒均成功表达;免疫共沉淀结果显示,pXJ41-HA-nsp4-D1与pXJ41-Flag-sRNase L发生了相互作用。见图5。
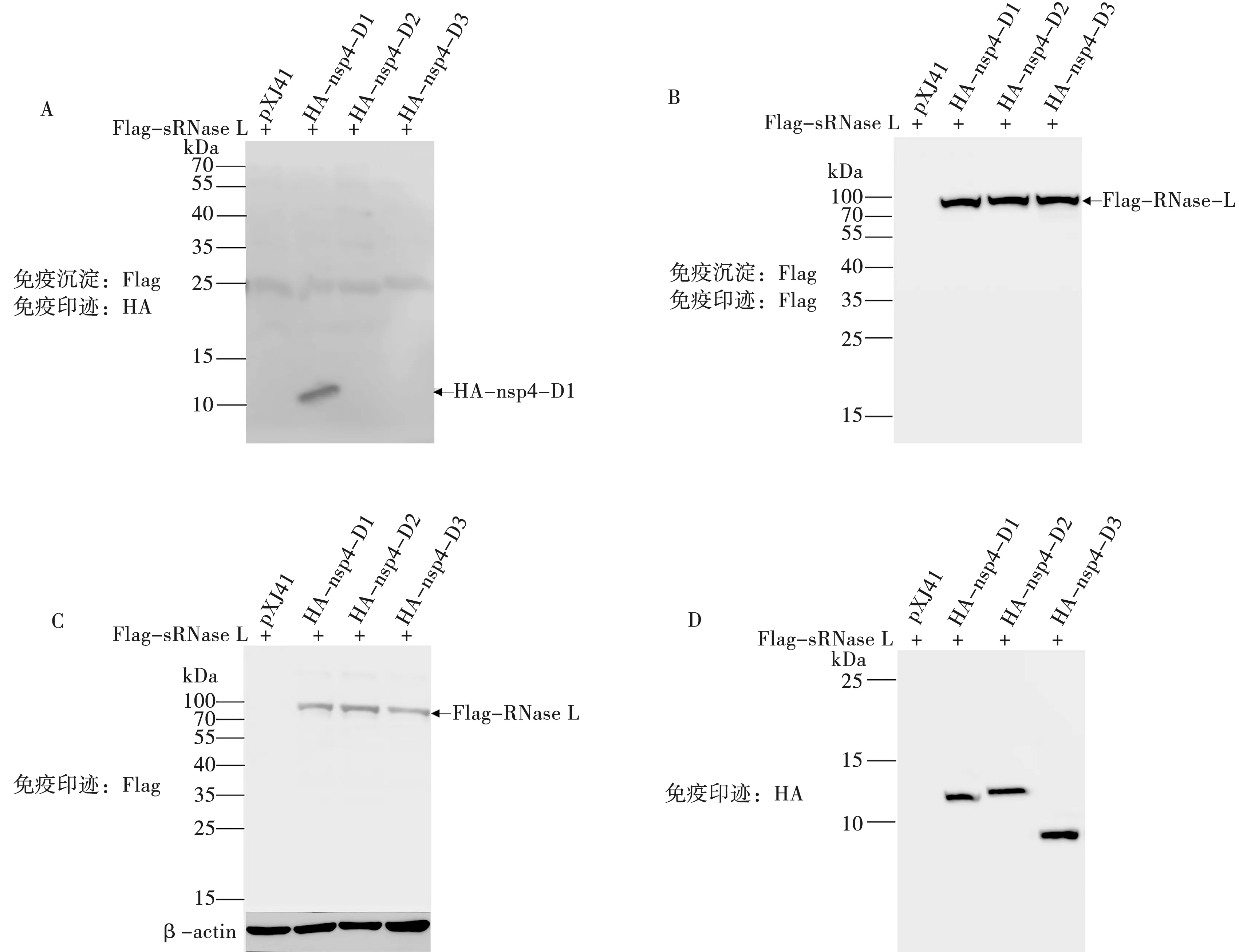
A. IP样品用兔源HA抗体做Western blot检测nsp4截短体互作条带;B. IP样品用兔源Flag抗体做Western blot检测sRNase L条带;C. Input样品直接做Western blot检测sRNase L的表达情况;D. Input样品直接做Western blot检测nsp4截短体的表达情况
3 讨论
PRRSV是目前影响世界养猪业最重要的病毒之一, nsp4参与了宿主的先天免疫应答,与病毒的免疫调节能力密切相关。nsp4蛋白酶具有3C样丝氨酸蛋白酶(3CLSP)活性[17]。本研究将sRNase L与hRNase L进行同源性比对,根据序列比对结果设计引物,利用本实验室保存的pXJ41-Flag-sRNase L质粒,构建得到质粒pXJ41-Flag-sRNase L突变体、缺失体pXJ41-Flag-sRNase L 1~586 aa;根据nsp4三个结构域设计引物,利用实验室保存的pXJ41-HA-nsp4质粒,构建得到质粒pXJ41-HA-nsp4-D1、pXJ41-HA-nsp4-D2、pXJ41-HA-nsp4-D3,在HEK-293T细胞中进行转染,通过SDS-PAGE和Western blot检测证实pXJ41-Flag-sRNase L突变体、缺失体pXJ41-Flag-sRNase L 1-586 aa、pXJ41-HA-nsp4-D1、pXJ41-HA-nsp4-D2、pXJ41-HA-nsp4-D3表达成功。
本研究将nsp4分成分别包含3个结构域的截短体pXJ41-HA-nsp4-D1(1~80 aa)、pXJ41-HA-nsp4-D2(60~156 aa)、pXJ41-HA-nsp4-D3(157~204 aa),通过Co-IP证实pXJ41-Flag-sRNase L能够与pXJ41-HA-nsp4-D1发生互作,其余2个结构域的截短体(pXJ41-HA-nsp4-D2、pXJ41-HA-nsp4-D3)均不能与sRNase L发生互作,说明nsp4的第一个结构域(pXJ41-HA-nsp4-D1)是与sRNase L发生互作的必要条件。图4A、图5A结果显示,免疫共沉淀后出现的条带为pXJ41-HA-nsp4和pXJ41-HA-nsp4-D1蛋白的大小,说明pXJ41-Flag-RNase L与pXJ41-HA-nsp4之间不是通过共价键结合的。
RNase L是一种内切单链RNA的核糖核酸酶,这种核酸酶只有很低的序列特异性[18]。目前人的RNase L(hRNase L)已研究比较透彻。Silverman等[14]研究发现R462Q的突变也终止了RNase L与2-5A结合区域的翻译,抑制了2-5A对RNase L的激活。Dong等[15]证明RNase L突变体W632A、D661A、R667A和H672A缺乏核糖核酸酶活性。Nakanishi等[16]试验证明Tyr712和Phe716对RNA的结合和切割功能起到重要的作用。sRNase L活性位点尚无报道,sRNase L和hRNase L氨基酸同源性为79.2%,且已报道的hRNase L活性位点在sRNase L序列中高度保守,我们根据hRNase L活性位点构建了对应的pXJ41-Flag-sRNase L R460A、pXJ41-Flag-sRNase L R675A、pXJ41-Flag-sRNase L Y720A、pXJ41-Flag-sRNase L F724A,Co-IP证实sRNase L第3结构域(587~744 aa)为与nsp4互作区段,但不包括第675位精氨酸、第720位酪氨酸、第724位苯丙氨酸。本研究确定与nsp4互作的sRNase L的区段为第3结构域(587~744 aa),并将与sRNase L互作的nsp4区段缩小到1~60 aa,为进一步探究sRNase L与nsp4互作抗PRRSV机制奠定了坚实的基础。
猜你喜欢
广州大学学报(自然科学版)(2019年1期)2019-05-07
食品科学(2018年10期)2018-05-23
安徽医科大学学报(2016年12期)2017-01-15
山东农业工程学院学报(2016年6期)2016-12-01
天津科技大学学报(2016年1期)2016-02-28
湖北师范大学学报(自然科学版)(2015年2期)2016-01-10
天津医科大学学报(2015年2期)2015-12-22
西南医科大学学报(2015年1期)2015-08-22
中国当代医药(2015年9期)2015-03-01
山东医药(2015年40期)2015-02-28
