
伪狂犬病病毒变异株UL3、UL46和UL50基因失活株的构建与生物学特性分析
2021-11-11 08:40殷涵臻张传健曾荣愚费荣梅王继春
畜牧与兽医 2021年11期
殷涵臻,张传健,曾荣愚,费荣梅,王继春
(1. 南京农业大学动物医学院,江苏 南京 210095;2. 江苏省农业科学院动物免疫工程研究所,江苏 南京 210014;3. 江苏省农业科学院国家兽用生物制品工程技术研究中心,江苏 南京 210014;4. 江苏省动物重要疫病与人兽共患病防控协同创新中心,江苏 扬州 225009;5. 天康制药(苏州)有限公司,江苏 苏州 215000)
伪狂犬病病毒(pseudorabies virus,PRV)又称猪疱疹病毒1型(suid herpesvirus,SuHV-Ⅰ),最早由匈牙利科学家所分离[1]。该病毒可引发伪狂犬病(pseudorabies,PR),许多家畜和野生哺乳动物感染该病会出现神经、呼吸和生殖系统等的症状,如瘙痒(小鼠)、繁殖障碍(母猪)、呼吸困难(成年猪)、呕吐抽搐(仔猪)等[2]。20世纪50年代,我国猪场首次暴发PR,后使用源自匈牙利的弱毒苗Bartha-K61使该病得到了有效控制[3]。然而,2011年以来,PRV变异株在我国猪群中暴发流行,其致病力和传播力明显高于传统毒株,传统的Bartha-K61已无法提供完全保护[4-5],新型疫苗的研制迫在眉睫。
TK、PK、gE/gI等为PRV重要的毒力因子,对病毒在哺乳动物机体中的潜伏、入侵有重要作用[6]。科研工作者常把这些基因作为基因工程疫苗的首选靶基因。1985年,Kit等[7]构建了第一株PRV TK基因缺失疫苗。目前,应用变异株研发的gE/gI、gE/TK和TK/gE/gI缺失株可提供有效的保护,但对初生PRV阴性仔猪存在严重的安全隐患[8],暗示其他基因可能影响其毒力。
UL3属于早期核蛋白,编码237个氨基酸,大小约为34 kDa[9]。目前UL3的功能尚未明确。研究发现,UL3对单纯疱疹病毒1型(herpes simplex virus type 1,HSV-1)体外复制无显著影响[10],但可与UL4、ICP22等相互作用从而影响蛋白质在细胞内的定位[11]。Barbara等[9]发现,UL3蛋白缺失不影响PRV在兔肾细胞上增殖,但关于UL3对动物致病力的影响还未见报道。
UL46基因,编码PRV晚期蛋白。常与UL47、UL48、UL49基因共同成簇,是病毒被膜蛋白主要成分[12]。研究发现,HSV-1和马立克氏病病毒(Marek’s disease virus,MDV)的UL46皆为非必需蛋白,缺失这类蛋白会影响病毒复制[13-14]。PRV UL46突变株(基因移码)对小鼠的致病力无显著影响[15]。
UL50基因,编码240个氨基酸,具有焦磷酸酶活性。研究发现诸多γ疱疹病毒的UL50可进行免疫调控,但不依赖于焦磷酸酶活性[16-17]。2017年,Zhang等[18]研究发现,PRV和HSV-1的UL50均可诱导Ⅰ型干扰素受体降解,从而拮抗干扰素反应。然而,目前尚无UL50对毒力影响的报道。
本研究以PRV变异株的UL3、UL46和UL50为研究对象,运用细菌人工染色体技术(bacterial artificial chromosome,BAC)构建了PRV AH02LA变异株UL3、UL46和UL50基因失活株:PRV-UL3knock、PRV-UL46knock和PRV-UL50knock,并对其生物学活性进行了分析,为PRV致病机制的研究提供了参考。
1 材料与方法
1.1 细胞、病毒、BAC与实验动物
PRV AH02LA株由本研究室分离于安徽六安某猪场PR阳性猪[19];含BACPRV(以PRV AH02LA株为亲本毒株,mini-F替换gI和gE基因)的大肠杆菌GS1783由本研究室制备[20];猪睾丸(swine testis,ST)细胞于本研究室保存。85只SPF级实验小鼠购自南京市青龙山动物养殖场(合格证编号:20210125Abzz01000000805)。
1.2 主要试剂
LATaqDNA聚合酶、PrimeSTAR Max聚合酶、DL10000 DNA Marker、DL15000 DNA Marker和250 bp DNA ladder购自TaKaRa公司;胎牛血清、DMEM培养液购自Gibco公司;DNA提取液购自Solarbio公司;DNA提取试剂盒购自QIAGEN公司;脂质体转染试剂盒购自Thermo Fisher公司。
1.3 引物设计与合成
引物由擎科生物科技有限公司合成,如表1所示。

表1 引物序列
1.4 重组菌株的构建
以卡那霉素(KAN)抗性基因为模板,分别以UL3knock-KAN F/R、UL46knock-KAN F/R和UL50knock-KAN F/R为引物,PCR扩增含各基因起始密码子前后同源臂序列的KAN抗性基因。PCR片段回收后,导入含BACPRV的GS1783中进行电转重组(电压:1 500 V;电阻:200 Ω;电容:25 μF),获得3株重组菌株:BACPRV-UL3knock-KAN、BACPRV-UL46knock-KAN和BACPRV-UL50knock-KAN。第二次重组中,2 mL 3株重组菌株感受态分别加入含2%阿拉伯糖的等量液体培养基,42 ℃振摇15 min,诱导环化重组酶表达,进行Red重组,敲除KAN抗性基因,筛选3株阳性克隆,分别命名为:BACPRV-UL3knock、BACPRV-UL46knock和BACPRV-UL50knock。应用引物UL3 F/R、UL46 F/R和UL50 F/R对3株BAC进行PCR和测序鉴定。
1.5 重组病毒的获得与鉴定
应用DNA提取试剂盒提取PRV AH02LA病毒DNA,以gE/gI F/R为引物,PCR扩增回收含同源臂的gE/gI序列片段。BACPRV-UL3knock、BACPRV-UL46knock和BACPRV-UL50knockDNA通过碱裂解法提取。应用脂质体转染法,将10 μL含同源臂的gE/gI序列片段分别与15 μL 3株重组BAC的DNA片段共转染单层ST细胞,转染后24 h,在紫外光(488 nm)激发下可见未发荧光的病毒蚀斑,连续挑斑纯化3代,获得3株重组病毒,分别命名为:PRV-UL3knock、PRV-UL46knock和PRV-UL50knock。PCR和测序鉴定gE/gI基因序列。
1.6 重组病毒遗传稳定性检测
重组病毒PRV-UL3knock、PRV-UL46knock和PRV-UL50knock分别在ST细胞上连续传代20次,提取F20代病毒基因组DNA,用UL3 F/R、UL46 F/R和UL50 F/R分别对UL3、UL46和UL50基因片段进行扩增,并通过测序鉴定各基因起始密码子是否发生回补或突变。
1.7 重组病毒体外生长动力学检测
将PRV AH02LA、PRV-UL3knock、PRV-UL46knock和PRV-UL50knock病毒分别以0.01感染复数(multiple of infection,MOI)接种ST细胞。于接种后6、12、24、36、48和72 h收集细胞和病毒混合液。经-70 ℃和37 ℃反复冻融3次,12 000 r/min离心10 min,检测上清病毒滴度,重复3次。应用SPSS 11.0进行统计学分析。
1.8 重组病毒对小鼠致病力测定
选取85只SPF级实验小鼠,随机分为17组,每组5只(表2)。取PRV AH02LA(107.75TCID50/mL)、PRV-UL3knock(107.2TCID50/mL)、PRV-UL46knock(106.5TCID50/mL)和PRV-UL50knock(107.3TCID50/mL)种毒,10倍倍比稀释,稀释度由100~10-3。攻毒采用颈部皮下注射法,每只小鼠注射0.1 mL的病毒液,对照组注射等量DMEM培养液。攻毒后每日观察记录小鼠临床症状,累计14 d,并统计死亡情况。Reed-Muench法计算PRV AH02LA、PRV-UL3knock、PRV-UL46knock和PRV-UL50knock对小鼠的LD50。

表2 小鼠试验分组
2 结果
2.1 重组菌株BACPRV-UL3knock、BACPRV-UL46knock和BACPRV-UL50knock的获得与鉴定
通过第一步Red重组,BACPRV插入KAN抗性基因的同时删除UL3、UL46和UL50基因起始密码子,PCR检测发现,对应菌株的UL3、UL46和UL50基因序列大小增加约1 000 bp(图1)。然后通过第二步Red重组,敲除KAN抗性基因,分别命名为:BACPRV-UL3knock、BACPRV-UL46knock和BACPRV-UL50knock。PCR(图1)和测序表明UL3、UL46和UL50起始密码子已成功敲除。

M. Marker;1. UL3;2. 敲除KAN基因;3. 插入KAN基因;4. UL46;5. 敲除KAN基因;6. 插入KAN基因;7. UL50;8. 敲除KAN基因;9. 插入KAN基因
2.2 重组毒株PRV-UL3knock、PRV-UL46knock和PRV-UL50knock的获得与鉴定
BACPRV-UL3knock、BACPRV-UL46knock和BACPRV-UL50knockDNA分别与带有同源臂的gE/gI序列片段共转染ST细胞,拯救病毒的同时,gE/gI序列替换mini-F序列,在紫外光(488 nm)下可见未发荧光的病毒蚀斑,经3轮挑斑纯化,获得重组病毒:PRV-UL3knock、PRV-UL46knock和PRV-UL50knock(图2)。gE/gI序列经PCR(图3)和测序鉴定正确。

图2 重组病毒PRV-UL3knock、PRV-UL46knock和PRV-UL50knock的纯化

M. DL15000 DNA Marker; 1. PRV AH02LA; 2. PRV-UL3knock; 3. PRV-UL46knock; 4. PRV-UL50knock
2.3 重组毒株PRV-UL3knock、PRV-UL46knock和PRV-UL50knock遗传稳定性测定
将重组病毒PRV-UL3knock、PRV-UL46knock和PRV-UL50knock于ST细胞中传代20次,提取F20代重组病毒DNA,对UL3、UL46和UL50基因进行PCR和测序鉴定,结果表明UL3、UL46和UL50起始密码子未有发生回补或突变(数据未列出),表明遗传稳定性良好。
2.4 重组毒株PRV-UL3knock、PRV-UL46knock和PRV-UL50knock体外生长动力学检测
PRV AH02LA、PRV-UL3knock、PRV-UL46knock和PRV-UL50knock分别以0.01 MOI接种ST细胞,在指定时间点检测上清与细胞混合物滴度,并绘制其生长曲线(图4)。PRV-UL3knock、PRV-UL46knock和PRV-UL50knock在感染后12 h(PRV-UL3knock:102.6TCID50/mL; PRV-UL46knock:0;PRV-UL50knock:102.75TCID50/mL)和36 h(PRV-UL3knock:106.5TCID50/mL;PRV-UL46knock:106.38TCID50/mL;PRV-UL50knock:106.55TCID50/mL)病毒滴度明显低于亲本毒株(12 h:103.0TCID50/mL;36 h:107.5TCID50/mL)。PRV-UL46knock在感染后6 h和12 h未检测到毒价。以上结果表明,UL3、UL46和UL50可能介导病毒复制。PRV AH02LA、PRV-UL3knock、PRV-UL46knock和PRV-UL50knock滴度峰值分别为:107.5TCID50/mL、106.8TCID50/mL、106.7TCID50/mL和107.1TCID50/mL。

图4 PRV AH02LA、PRV-UL3knock、PRV-UL46knock和PRV-UL50knock体外生长动力学
2.5 PRV AH02LA、PRV-UL3knock、PRV-UL46knock和PRV-UL50knock对小鼠致病力分析
根据表3统计,计算PRV-UL3knock、PRV-UL46knock和PRV-UL50knock的LD50依次为:104.44TCID50、104.5TCID50和104.05TCID50,与PRV AH02LA亲本毒均相似(LD50:104.91TCID50)。表明UL3、UL46和UL50对小鼠致病性无显著影响。
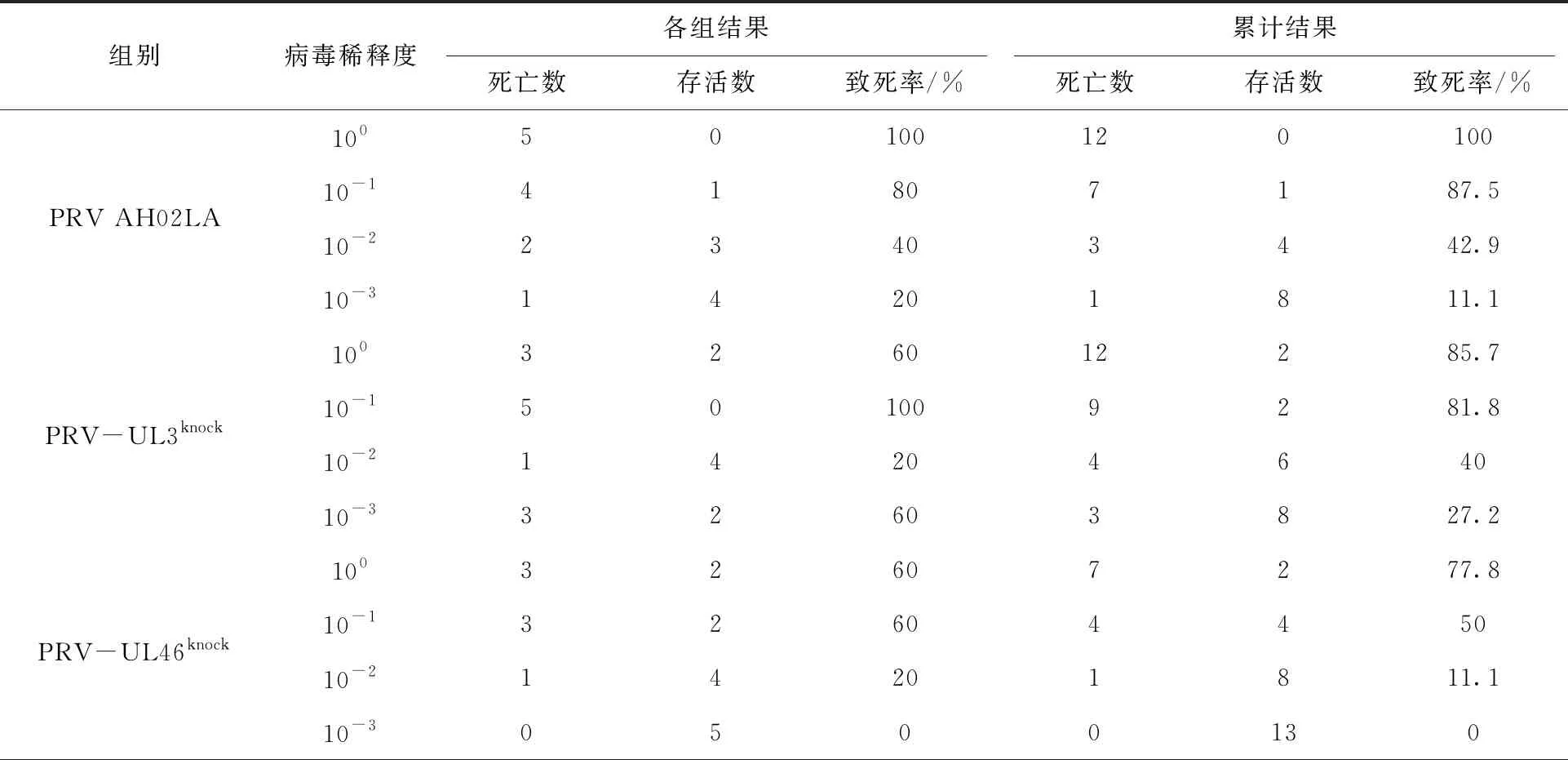
表3 小鼠试验结果

续表3
3 讨论
2011年以来,PRV变异株给我国养猪业带来了巨大损失[21-22],新型疫苗的研发是控制该病的关键,一些未知毒力基因的鉴定尤为重要。目前关于UL3、UL46和UL50对PRV复制和毒力的影响尚不明确。因此,本研究通过BAC以及配套的EnPassant技术敲除UL3、UL46和UL50基因起始密码子,构建3种基因失活株,并进行了体外生长特性和对小鼠致病性的研究。
本研究所采取BAC及其相关的EnPassant技术是疱疹病毒研究的最先进技术,BAC技术使得对疱疹病毒基因组进行缺失、突变和插入的分子生物学操作可以在大肠杆菌中进行,能大大提高研究效率,且基因不易发生变异[23]。本研究第一步重组是将包含UL3、UL46和UL50基因起始密码子前后同源臂序列的KAN抗性基因电转至大肠杆菌GS1783,与BACPRV进行同源重组。第二步重组基于温敏启动子调控,阿拉伯糖诱导Cre酶表达,完成KAN抗性基因的敲除,从而构建3种基因失活株。
体外生长性能试验发现,UL3、UL46和UL50突变失活株在感染后12和36 h滴度显著低于PRV AH02LA亲本株,暗示UL3、UL46和UL50可能介导病毒复制。UL3编码病毒的早期核蛋白,先前研究表明UL3缺失对PRV在兔肾细胞上的复制无显著影响[9],因此,关于UL3对PRV在不同细胞复制的影响及其机制需要进一步研究。PRV UL46蛋白可通过拮抗细胞Ⅰ型干扰素反应[24],从而促进了病毒对细胞的入侵。此外,UL46蛋白可调节核膜破裂(nuclear envelope breakdown,NEBD),NEBD可促进病毒排出细胞[25-26],从而加快对其他细胞的入侵。对HSV-1研究发现,UL46缺失降低HSV-1复制水平,减少蚀斑大小[27]。研究表明,PRV与HSV-1 UL50均可以通过降解干扰素受体,抑制α-干扰素反应[14],促进病毒免疫逃逸。以上研究在一定程度上解释了UL3、UL46和UL50失活株在感染12和36 h低于亲本株的原因,具体作用机制还需进一步研究。此外,后续研究将应用q-PCR对病毒含量进行验证,也将通过兔肾细胞和BHK-21细胞对3种失活株生长动力学进行鉴定。
小鼠作为PRV的非自然宿主,可引起极高的致死率。对小鼠致病力试验发现,PRV-UL3knock、PRV-UL46knock、和PRV-UL50knock对小鼠的LD50与PRV AH02LA亲本株相似,表明3种蛋白对小鼠致病性无显著影响。为准确鉴定UL3、UL46和UL50基因是否失活,之后还需通过Western blot进行验证。
综上所述,本研究通过BAC以及配套的EnPassant技术构建了UL3、UL46和UL50突变失活株。UL3、UL46和UL50突变失活株在感染12和36 h滴度显著低于PRV AH02LA亲本株,表明UL3、UL46和UL50可能介导病毒复制,具体作用机制需要进一步研究。此外,本研究首次分析了UL3、UL46、UL50基因对PRV毒力的影响,为PRV致病机制的研究提供了参考。
猜你喜欢
传染病信息(2021年6期)2021-02-12
生物学通报(2020年11期)2020-10-22
科海故事博览·下旬刊(2019年6期)2019-04-16
中成药(2018年7期)2018-08-04
生物医学工程学进展(2015年1期)2015-02-28
河南科技(2015年2期)2015-02-27
化学工业与工程(2015年1期)2015-02-10
茶叶通讯(2014年4期)2014-02-27
食品科学(2013年19期)2013-03-11
中国烟草学报(2012年2期)2012-04-09
