
甜菜氮代谢相关蛋白BvAMT3-3基因的克隆及生物信息学分析
2021-08-09 09:36:56李佳佳刘铭曦马龙彪刘大丽
黑龙江大学自然科学学报 2021年3期
魏 多, 李佳佳, 高 卓,2, 刘铭曦,马龙彪, 兴 旺, 刘大丽
(1.黑龙江大学 现代农业与生态环境学院 省高校甜菜遗传育种重点实验室, 哈尔滨150080;2.黑龙江大学 生命科学学院, 哈尔滨 150080; 3.哈尔滨师范大学 生命科学与技术学院, 哈尔滨 150025)
0 引 言
作为世界第二大糖料作物,甜菜是除了甘蔗之外糖的主要来源之一,世界上约35%的糖是由甜菜供应而来的[1]。甜菜属于耐寒作物,在中国的种植区域主要位于东北三省、新疆和内蒙古等地。由于耕地的过度使用以及作物的不规范化生产,导致甜菜在种植过程中常常遭受氮胁迫等环境逆境的限制。植物在生长的过程中无时无刻不需要氮元素,并通过将无机氮转化为有机氮的方式来吸收氮素[2]。合理施用氮肥可以有效提高作物产量,但施用过度会导致病虫害增加和环境污染等问题[3-5]。基于此,使用铵基肥料和硝化抑制剂可减轻氮肥对环境的影响[6],根系对养分的吸收和运输能力与不同植物器官的需求相匹配,这也进一步决定了植物能否健康茁壮地生长。如果营养素可利用性下降或非生物性胁迫抑制了根系的正常功能,则会导致植物营养不足,降低它们的生态竞争力和农业生产力[7]。

本文在对甜菜AMTs基因的前期研究中,通过对受氮逆境胁迫的甜菜进行转录组测序发现,在参与甜菜氮逆境胁迫相关的众多基因中,BvAMT3-3基因的表达极其显著。因此,利用RT-PCR的方法克隆了甜菜BvAMT3-3基因,并通过qPCR分析氮胁迫下目的基因的表达特性,结合生物信息学数据库和生物软件对BvAMT3-3的基因结构和编码的蛋白质特点进行了分析和预测,为进一步了解甜菜AMTs基因以及甜菜中铵盐的吸收机制奠定基础,对提高甜菜氮素的利用效率具有重要意义。
1 材料与方法
1.1 植物材料
供试甜菜为国家甜菜种质资源中期库提供的780016B/12优。种子通过蛭石培养萌发后,将两片子叶期的甜菜幼苗培养于N浓度为10 mmol·L-1的Hoagland营养液中,培养两周后,取甜菜幼叶组织用于RNA的提取。
1.2 实验方法
1.2.1 RNA的提取及cDNA的合成
利用Invitrogen公司的Trizol reagent提取甜菜新鲜材料的总RNA[17],并根据北京TransGen公司的反转录试剂盒合成cDNA,于-20 ℃保存备用。
1.2.2BvAMT3-3基因的克隆
以甜菜总RNA反转录的cDNA为模板,利用KOD-Plus-Neo高保真酶(TOYOBO)PCR扩增该基因CDS全长。根据甜菜BvAMT3-3基因的碱基序列设计引物如下:
BvAMT3-3-F (5′-3′): CGGAA TTCATG GGA GAC ATA TTG CCA CCC
BvAMT3-3-R (5′-3′): GCTCT AGATTA TAC AAC TTG AGT TGC ACC AA
PCR产物电泳检测后,将回收产物与载体pEASY Blunt连接,转入T1感受态细胞。加入含LB的培养基后倒平板并摇置涂匀,37 ℃摇菌过夜后,筛选出阳性克隆的菌落进行PCR检测,选择电泳检测结果正确的菌液送上海生工生物工程公司进行测序。
1.2.3 生物信息学分析
在NCBI数据库中搜索甜菜BvAMT3基因家族成员;同时BvAMT3-3的开放阅读框和CDS通过NCBI的ORF finder分析得到;利用NCBI保守结构域数据库预测该基因所编码蛋白的保守结构域。通过PSORT对甜菜AMT3-3蛋白进行亚细胞定位;通过TMHHM分析跨膜结构域数目。利用Prot-Param和ProtScale分析蛋白质理化性质,SignalP 5.0 Server进行信号肽预测分析。利用MEGA7软件将甜菜AMT2家族的基因序列与其他植物的AMT2成员进行聚类分析和Neighbor-Joining进化树的构建。
1.2.4 植物培养及逆境处理
甜菜种子于蛭石中培养一周,待种子萌发后,选取生长良好的甜菜幼苗移栽入N浓度为0、1.5和10 mmol·L-1的Hoagland营养液中,N0为缺氮,N1.5为低氮,以10 mmol·L-1生长的甜菜幼苗为对照。取处理0、24和48 h时间点植株的相同部位叶片作为样品,然后将甜菜幼苗移至正常培养条件,取恢复生长24和48 h的植株样品,经研磨后用于测量相关基因的表达量。
1.2.5BvAMT3-3基因的实时荧光定量PCR分析
根据甜菜BvAMT3-3基因(XM_010669058)碱基序列信息设计特异引物,以Trizol法提取的甜菜叶片RNA为模板,以BvGAPDH(NC_024800)基因为内参,利用SuperReal PreMix Plus (TianGen)进行qRT-PCR扩增,扩增时采用SYBR Green为标记物。每个样品3次重复,利用2-△△CT方法对目的基因表达的数据进行统计和分析。

表1 相关qRT-PCR引物序列及产物大小
2 结果与分析
2.1 甜菜BvAMT3-3基因的克隆
以甜菜总RNA反转录合成的cDNA为模板,用带有EcoRI和Xbal酶切位点的特异性引物进行RT-PCR扩增,获得了一条大约1 420 bp的BvAMT3-3扩增片段,如图1所示。
将BvAMT3-3基因经载体连接,Kana阳性克隆筛选后,对所得的阳性克隆进行质粒提取,双酶切电泳检测。得到了大小为3 928 bp的载体pEASY-Blunt条带和大小为1 427 bp的目的基因片BvAMT3-3的条带,如图2所示。

M: Trans 2K Plus II Marker(TransGen)
2.2 甜菜BvAMT3-3基因的序列分析
甜菜BvAMT3-3基因的CDS全长为1 427 bp,编码475个氨基酸,如图3所示。根据ProtParam分析计算,这个基因所编码蛋白的分子量(Molecular weight)为51.055 kDa,等电点(Theoretical pI)为8.98。蛋白的不稳定指数(Instability index)为29.61,属于稳定蛋白。根据SignalP 5.0分析,该蛋白没有信号肽,不属于分泌蛋白。对甜菜AMT蛋白二级结构进行预测分析,结果表明BvAMT3-3蛋白以α-螺旋为主,占43.79%,其次为无规则卷曲,占34.53%,而延伸链和β-转角所占比例较小,如表2所示。亚细胞定位预测BvAMT3-3定位于内质网膜结构上。


图3 甜菜BvAMT3-3基因序列信息

表2 BvAMT3-3蛋白的二级结构分析和亚细胞定位
2.3 BvAMT3-3蛋白的亲水性和疏水性预测
用 ProtScale 软件对蛋白质进行疏水性/亲水性分析,正值越大表示越疏水,负值越大表示越亲水,介于+0.5~-0.5之间的主要为两性氨基酸。结果表明,BvAMT3-3蛋白的疏水区域大于亲水区域,如图4所示,预测该蛋白为疏水蛋白,这与ProtParam分析结果一致。

图4 BvAMT3-3蛋白亲疏水性预测
2.4 BvAMT3-3蛋白跨膜结构域预测
运用TMHMM Server 2.0对BvAMT3-3蛋白的跨膜结构域进行预测。结果表明,BvAMT3-3含有11个跨膜螺旋区,属于跨膜蛋白,这与AMTs基因家族含有11个跨膜结构域的结果相一致,如图5所示。
2.5 甜菜BvAMT3-3蛋白保守结构域分析
利用NCBI保守结构域数据库CD-Search预测该基因所编码蛋白的保守结构域。结果显示,甜菜BvAMT3-3蛋白存在一个铵转运保守结构域,位置在25~443 aa,这保证了AMT在转运铵盐中发挥稳定的作用。

图5 BvAMT3-3蛋白的跨膜螺旋区分析
2.6 甜菜BvAMT3-3蛋白三级结构预测

图6 BvAMT3-3蛋白的三级结构预测图Fig.6 Prediction for tertiary structure of BvAMT3-3 protein
将甜菜BvAMT3-3的氨基酸序列提交到SWISSMODEL在线软件,采用同源建模法预测该蛋白的三级结构,结果如图6所示。结果显示,该蛋白的三级结构主要由无规则卷曲、延伸链和α-螺旋构成,与二级结构的预测结果基本一致。
2.7 甜菜BvAMT3-3基因的结构及进化树分析
外显子-内含子的数量和结构可以在一定程度上反映一些基因家族在进化过程中的变化情况[18]。通过NCBI数据库进行BLAST搜索到5个甜菜AMT基因家族成员,利用GSDS 2.0对5个AMT基因结构进行分析。结果表明,BvAMT3-3基因含有2个内含子,BvAMT1-1和BvAMT1-4均含有1个内含子,而BvAMT1-2和BvAMT1-3均没有内含子,如图7所示。对来自甜菜、菠菜、水稻、谷子等6个物种中的12个AMT3基因进行系统进化分析发现,甜菜BvAMT3-3基因与菠菜SoAMT3-1基因位于同一进化分支,如图8所示。

图7 甜菜AMT基因结构特征

图8 甜菜与其他物种之间的AMT3家族进化树
2.8 甜菜BvAMT3-3基因在低氮胁迫下的表达特性分析
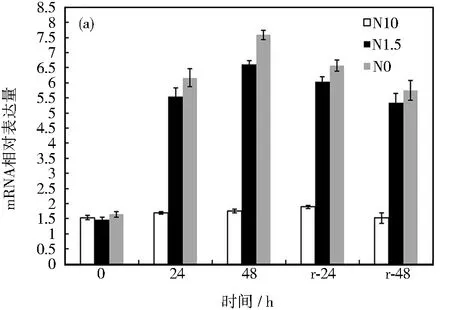
AMT基因是参与植物氮逆境胁迫应答的主要基因之一,它调控了植物体内铵转运蛋白的合成。在对甜菜进行氮胁迫处理前,各组甜菜中BvAMT3-3的表达量几乎基本处于相同水平,甜菜叶片部位的相对表达量在1.54左右,根部的表达量在1.27左右。当缺氮(N0)和低氮(N1.5)处理24 h后,BvAMT3-3在叶片和根部的表达量均表现出明显的上调,并且随着处理时间的增加,表达量呈现逐步上升的趋势,且缺氮条件比低氮更容易诱导BvAMT3-3的表达。其中,叶片和根部的BvAMT3-3表达量均在48 h时达到最大,分别为0 h的4.62和3.32倍,可以看出N0处理更能诱导BvAMT3-3的表达。同时,该基因在叶片的表达比根部更为活跃,如图9所示。恢复处理24和48 h后,BvAMT3-3表达量较胁迫处理时有所下降,但仍维持在较高水平。这说明即便恢复正常生长条件,氮逆境造成的细胞内反应还依旧存在,并且很有可能通过BvAMT3-3介导合成铵转运蛋白,来调控氮代谢平衡。
3 讨 论

4 结 论
本文利用RT-PCR克隆了基因BvAMT3-3,与生物信息学分析的结果相结合,发现该基因编码的蛋白可能会作为转运蛋白参与到甜菜对氮素的吸收和转运网络中。当受到氮逆境胁迫时,BvAMT3-3基因表达量随处理时间的增长呈现上升趋势,表达的水平远大于未受到氮胁迫处理时的水平,表明该基因参与了甜菜对铵态氮吸收循环再利用的过程。甜菜受到氮逆境胁迫时,BvAMT3-3基因可能会参与到逆境胁迫的应答网络中,可能通过直接或间接的作用降低外界铵态氮含量的变化对细胞的损害,从而提高甜菜抵抗氮逆境胁迫的能力。研究结果有助于下一步研究不同氮水平条件下的基因表达特征、染色体定位和启动子顺式作用元件,同时也为加深植物吸收氮素的生理和分子机制的研究奠定基础。
猜你喜欢
做人与处世(2022年6期)2022-05-26 10:26:35
疯狂英语·新阅版(2022年5期)2022-04-29 05:07:07
黑龙江大学自然科学学报(2021年4期)2021-11-19 07:05:04
小哥白尼(趣味科学)(2021年12期)2021-03-16 05:40:44
心肺血管病杂志(2019年1期)2019-04-22 01:12:00
教学考试(高考生物)(2017年4期)2017-12-13 09:02:41
中国糖料(2016年1期)2016-12-01 06:49:04
高中生学习·高三版(2014年3期)2014-04-29 06:12:54
新疆农垦科技(2014年3期)2014-02-28 19:19:32
华东理工大学学报(自然科学版)(2014年2期)2014-02-27 13:48:43
