
HCG11靶向miR-1297对肺癌细胞增殖、迁移、侵袭的影响及其机制
2021-07-27 08:19李亚明潘双陈晖刘松张进召
中国老年学杂志 2021年14期
李亚明 潘双 陈晖 刘松 张进召
(西安医学院第一附属医院 西安医学院全科医学院呼吸与危重症医学科,陕西 西安 710077)
肺癌是起源于支气管上皮细胞或支气管腺体的恶性肿瘤,其发病率和死亡率均居恶性肿瘤首位〔1〕。尽管肺癌的综合诊疗技术不断提高,但由于肺癌极易扩散和转移,对人类的健康和生命已构成严重威胁〔2,3〕。长链非编码RNA(LncRNA)是一类长度大于200个核苷酸的非编码RNA,越来越多的研究证实LncRNA与恶性肿瘤的发生发展密切相关〔4,5〕。研究发现,人类白细胞抗原复合体(HCG)11在非小细胞肺癌(NSCLC)组织中表达下调,HCG11的下调程度与存活时间显著相关,是NSCLC的新型生物标志物和诊断靶标〔6〕。此外,有研究指出HCG11的异常表达参与对肝癌和宫颈癌细胞的生物学行为调控〔7,8〕。但目前HCG11对肺癌细胞增殖迁移等生物行为的影响及其机制目前尚不清楚。本研究探究HCG11在肺癌转移中的作用机制。
1 材料与方法
1.1实验材料 46例肺癌组织及与其配对的癌旁组织由西安医学院第一附属医院于2016年1月至2018年1月收集。所有患者以前未经过化疗和放疗。本研究在开始前经医院伦理委员会批准,患者均签订知情同意书。肺癌细胞A549购自美国ATCC,DMEM培养基、胎牛血清、青链霉素双抗溶液、LipofectamineTM2000转染试剂和Trizol试剂均购自Invitrogen公司;四甲基偶氮唑蓝(MTT)细胞增殖检测试剂盒和Western印迹相关试剂均购自中国碧云天公司;Transwell小室和基质胶均购自BD公司;pcDNA3.1、pcDNA3.1-HCG11、miR-NC、miR-1297 mimics、si-NC、si-HCG11、WT-HCG11和MUT-HCG11的构建和测序均由上海生工公司提供;细胞周期蛋白(Cyclin)D1抗体、P21抗体、E-钙黏附蛋白(E-cadherin)抗体和基质金属蛋白酶(MMP)-2抗体均购自Abcam公司;辣根过氧化物酶(HRP)标记的二抗购自北京中杉金桥生物技术有限公司。
1.2方法
1.2.1细胞培养、转染和实验分组 采用DMEM培养基(含有10%胎牛血清和1%青链霉素双抗)于常规细胞培养箱(37℃、含5%CO2、湿度饱和)培养A549细胞。细胞转染:将对数期的A549细胞按照每孔2×105个细胞的密度接种于6孔板,当细胞融合度达到60%时,用不含血清培养基同步化12 h后进行转染。将pcDNA3.1或pcDNA3.1-HCG11溶解于Opti-MEM培养基中室温条件孵育5 min,同时另取LipofectamineTM2000加入到Opti-MEM培养基室温孵育5 min,然后将两者混合后室温放置20 min,最后将含pcDNA3.1或pcDNA3.1-HCG11的混合物分别加入到A549细胞中,培养6 h后,更换为DMEM培养基继续培养24、48、72 h后进行后续实验。为进一步证明HCG11是通过调控miR-1297表达进而影响A549细胞的增殖和凋亡,将pcDNA3.1-HCG11和miR-1297 mimics同时转染入A549细胞,检测A549细胞的增殖和凋亡情况。实验分组如下:pcDNA3.1组(转染pcDNA3.1)、pcDNA3.1-HCG11组(转染pcDNA3.1-HCG11)、pcDNA3.1-HCG11+miR-NC组(同时转染pcDNA3.1-HCG11和miR-NC)和pcDNA3.1-HCG11+ miR-1297组(同时转染pcDNA3.1-HCG11和miR-1297 mimics)。
1.2.2实时荧光定量-聚合酶链反应(qRT-PCR)检测 利用Trizol试剂分别提取46例肺癌组织和与其配对的癌旁组织应激各组A549细胞的总RNA。利用逆转录酶合成cDNA,并以cDNA为模板按照说明书进行qRT-PCR扩增。用2-ΔΔCt法计算HCG11和miR-1297的相对表达量,分别以GAPDH和U6为内参。引物序列如下:miR-1297上游引物:5′-ACACTCCAGCTGGGTTCAAGTAATTCAGG-3′,下游引物:5′-CTCAACTGGTGTCGTGGAGTCGGCAATTC-3′,HCG11上游引物:5′-GCTCTATGCCATCCTGCTT-3′,下游引物:5′-TCCCATCTCCATCAACCC-3′,GAPDH上游引物:5′-TGTTCGTCATGGGTGTGAAC-3′,下游引物:5′-ATGGCATGGACTGTGGTCAT-3′,U6上游引物:5′-GATTTCTCCCTCATCGCTTACAG-3′,下游引物:5′-CTGCTTCATGATCGTTGTTGCTTG-3′。
1.2.3双荧光素酶报告基因检测 利用Starbase进行靶基因预测发现,HCG11与miR-1297存在连续结合位点,猜测miR-1297是HCG11的靶基因,并利用双荧光素酶报告基因检测进行验证。构建野生型WT-HCG11和突变型MUT-HCG11的HCG11-3′UTR荧光素酶报告基因载体,利用LipofectamineTM2000将miR-1297模拟物和miR-NC分别与WT-HCG11或MUT-HCG11同时转染入A549细胞,常规培养48 h后,利用双荧光素酶报告基因检测试剂盒测定各组细胞的荧光素酶活性。
1.2.4MTT比色法检测细胞活力 分别取各组对数生长期细胞,胰酶消化后接种(2×103个/孔)于96孔板,分别在24、48和72 h时加入MTT试剂(20 μl/孔)孵育4 h,弃去上清液后每孔再加入DMSO试剂(150 μl/孔),继续孵育2 h至结晶完全溶解。全自动酶标仪检测490 nm处各孔的OD值。
1.2.5Transwell法检测细胞迁移和侵袭能力 侵袭实验:取50 μl已稀释的基质胶包被于Transwell小室上室膜,凝固后备用。细胞转染48 h后,用胰酶消化细胞,用DMEM培养基(不含血清)重悬细胞调整细胞浓度约为2×105/ml。上室加入100 μl的细胞悬液,下室加入DMEM培养基(含10%血清),于细胞培养箱常规培养24 h后,用棉签擦去上室膜未穿膜的细胞,用甲醇固定下室膜30 min,0.5%的结晶紫染液染色10 min,洗去多余染液,倒置于显微镜下观察下室室面的细胞个数,随机选取5个视野进行拍照,计数,取均值为细胞侵袭数目。迁移实验采用未包被基质胶的Transwell小室,其他步骤同上。
1.2.6Western印迹检测 采用RIPA裂解液提取各组A549细胞的总蛋白,用BCA试剂盒进行定量,调整蛋白浓度。取适量蛋白样品,上样后进行十二烷基硫酸钠-聚丙烯酰胺凝胶电泳(SDS-PAGE),电转至聚偏氟乙烯(PVDF)膜后,用Western封闭液室温封闭60 min,Western洗涤液洗膜后,加入已稀释的相应的Ⅰ抗4℃侧摆摇床孵育过夜,Western洗涤液洗膜后,加入相应的已稀释的Ⅱ抗室温条件孵育1 h,化学发光显色后拍照,成像扫描分析系统测定目的条带的灰度值。
1.3统计学方法 采用SPSS20.0软件进行t检验、单因素方差分析。
2 结 果
2.1HCG11和miR-1297在肺癌组织和癌旁组织中的表达 与癌旁组织组比较,肺癌组织组HCG11的表达显著下调,miR-1297的表达显著上调(均P<0.05)。见表1。
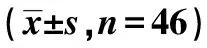
表1 HCG11、miR-1297在肺癌组织和癌旁 组织中的表达
2.2过表达HCG11对细胞A549增殖的影响 与pcDNA3.1组比较,pcDNA3.1-HCG11组A549细胞HCG11的表达显著上调(P<0.05),表明成功构建了过表达HCG11的A549细胞株。与pcDNA3.1组比较,pcDNA3.1-HCG11组A549细胞在24 h、48 h和72 h细胞活力显著降低,促增殖蛋白CyclinD1的表达显著降低,增殖抑制蛋白P21的表达显著上调(P<0.05)。表明过表达HCG11可抑制A549细胞的增殖。见表2、图1。

表2 过表达HCG11对细胞A549增殖的影响

图1 过表达HCG11对细胞A549增殖蛋白表达的影响
2.3过表达HCG11对细胞A549迁移、侵袭的影响 与pcDNA3.1组相比,pcDNA3.1-HCG11组A549细胞迁移侵袭促进蛋白MMP-2的表达显著降低,迁移侵袭抑制E-cadherin的表达显著升高,迁移侵袭细胞数目显著减少(P<0.05)。见图2、表3、图3。
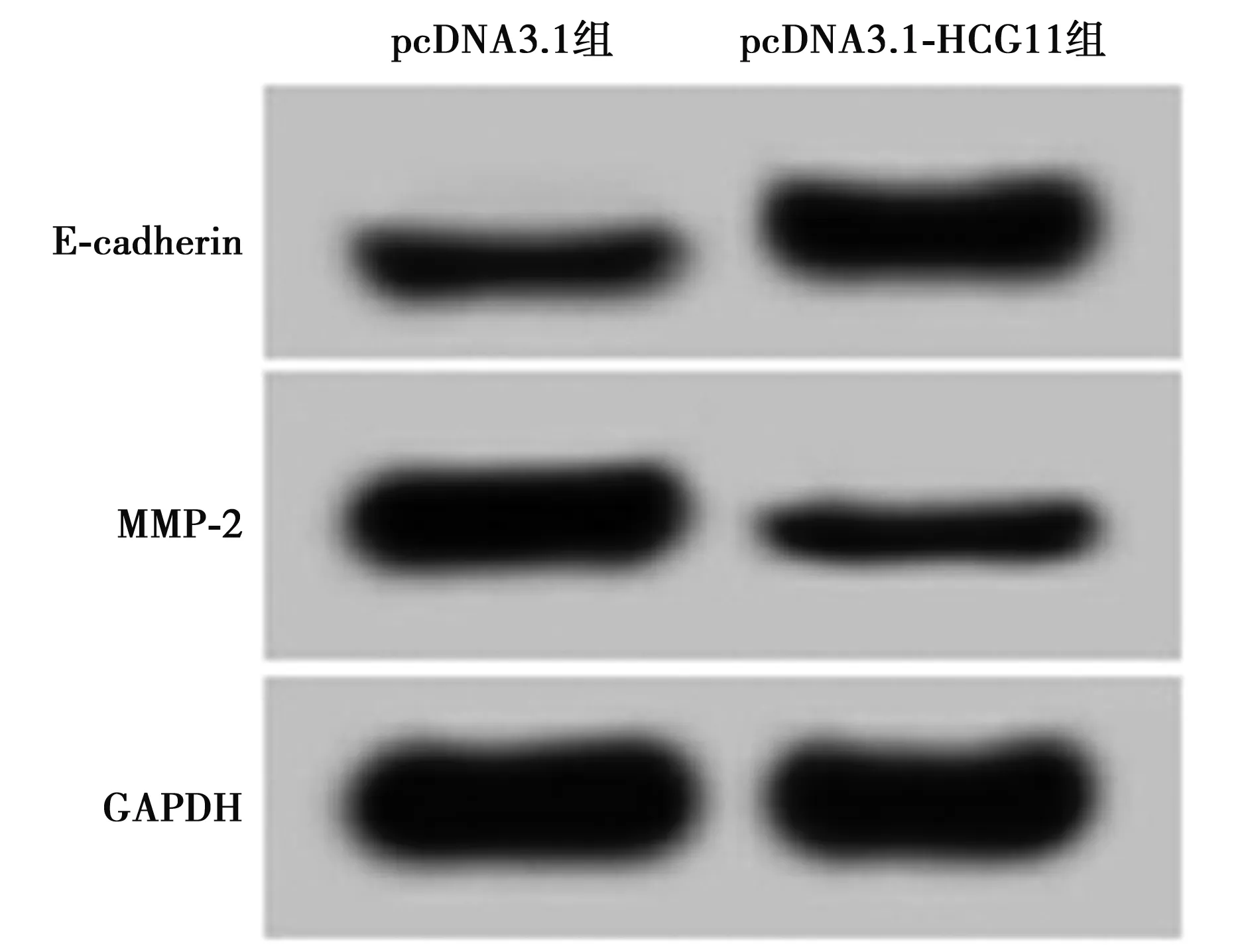
图3 过表达HCG11对细胞A549迁移、侵袭的影响
表3 过表达HCG11对细胞A549迁移、侵袭的影响
图2 过表达HCG11对细胞A549迁移、侵袭的 影响(结晶紫染色,×200)
表明过表达HCG11可抑制A549细胞的迁移和侵袭。
2.4HCG11靶向调控miR-1297的表达 Starbase在线预测显示,HCG11与miR-1297存在连续结合位点,见图4。当上调miR-1297表达后,转染WT-HCG11的3′UTR报告基因载体的A549细胞荧光素酶活性显著降低(P<0.05),而转染MUT-HCG11的3′UTR报告基因载体的A549细胞荧光素酶活性无显著变化(P>0.05)。见表4。与pcDNA3.1组A549细胞miR-1297的表达(0.88±0.09)相比,pcDNA3.1-HCG11组(0.30±0.03)显著降低;与si-NC组(0.89±0.09)相比,si-HCG11组(1.43±0.14)显著升高(P<0.05)。表明miR-1297是HCG11的靶基因,miR-1297可靶向负性调控HCG11的表达。

图4 HCG11靶向miR-1297

表4 双荧光素酶报告实验
2.5过表达miR-1297能逆转HCG11对细胞A549增殖的抑制作用 与pcDNA3.1组相比,与pcDNA3.1-HCG11+miR-NC比较,pcDNA3.1-HCG11+miR-1297组A549细胞miR-1297和Cyclin D1蛋白的表达显著升高,P21蛋白的表达显著降低,A549细胞在24 h、48 h和72 h细胞活力显著增强(P<0.05)。见图5、表5。表明过表达miR-1297可逆转HCG11对A549细胞的增殖抑制作用。
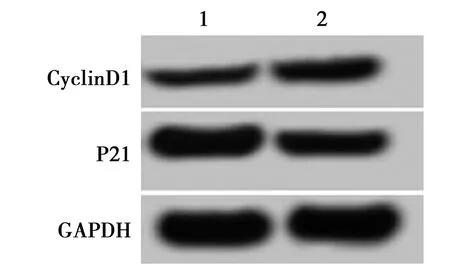
1~2:pcDNA3.1-HCG11+miR-1297组,下图同图5 过表达miR-1297能逆转HCG11对细胞A549 增殖蛋白表达的影响
表5 过表达miR-1297能逆转HCG11对细胞A549增殖的抑制作用
2.6过表达miR-1297能逆转HCG11对细胞A549迁移、侵袭的抑制作用 与pcDNA3.1-HCG11+miR-NC组相比,pcDNA3.1-HCG11+miR-1297组A549细胞E-cadherin蛋白的表达显著降低,MMP-2蛋白的表达显著升高,迁移和侵袭细胞数目显著增多(P<0.05)。见图6、表6。表明过表达miR-1297可逆转HCG11对A549细胞迁移和侵袭的抑制作用。
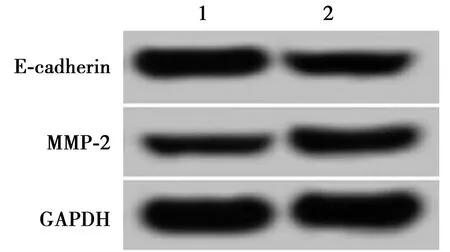
图6 过表达miR-1297能逆转HCG11对细胞A549迁移、 侵袭蛋白表达的影响
表6 过表达miR-1297能逆转HCG11对细胞A549迁移、侵袭的抑制作用
3 讨 论
肺癌是世界上最常见的实体肿瘤,NSCLC约占所有肺癌病例的80%,肺癌细胞的侵袭转移是肺癌相关死亡的主要原因〔9,10〕。但是,对肺癌细胞的侵袭转移的机制仍知之甚少,且目前尚无有效治疗肺癌细胞侵袭转移的有效方法。因此,了解肺癌细胞的侵袭转移机制,寻找新的治疗靶点,对提高肺癌治疗和改善患者预后具有重要意义。
LncRNA参与调控恶性肿瘤的发生发展已被证实〔11〕。Liu等〔12〕研究发现,HCG11在乳腺癌中表达上调,HCG11的异常表达与乳腺癌患者较差的总体生存率相关;Xu等〔7〕研究发现,HCG11在肝癌组织中也呈高表达,敲减HCG11导致细胞活力、增殖和迁移能力下降;然而,Zhang等〔13〕研究发现,HCG11在前列腺癌中表达下调,Wang等〔14〕进一步研究发现过表达HGC11通过下调miR-543表达抑制PI3K/AKT信号通路,进而抑制前列腺癌细胞的增殖、迁移和侵袭,并诱导细胞凋亡。本研究通过qRT-PCR证实HGC11在肺癌组织中呈低表达,这与常芬等〔6〕研究结果相一致。本研究结果表明HGC11在肺癌中发挥抑癌基因作用。HGC11下游靶基因众多,意味着HGC11有多种调控通路。在直肠癌中,HGC11通过靶向下调miR-144-3p,进而上调转录因子E盒结合锌指蛋白(ZEB)1的表达,促进直肠癌的发生和转移〔15〕;陈晓杰等〔16〕指出HGC11可能通过调控miR-590-3p表达进而影响宫颈鳞状细胞癌患者的预后发挥抑癌基因作用;HGC11还可直接作用于miR-4425,促进肿瘤转移相关基因(MTA)3表达,进而抑制胶质瘤的生长〔17〕。可见HGC11参与调控多种恶性肿瘤的生物学行为,但HGC11在肺癌中的分子机制尚不清楚。
miR-1297是基于不同的癌症细胞而发挥癌基因或抑癌基因作用的微小RNA〔18〕。卜文瑾〔19〕研究发现,miR-1297在肺癌细胞中表达上调,抑制miR-1297表达细胞增殖率降低。本研究发现miR-1297是HGC11的潜在靶基因。因此,推测HGC11/miR-1297分子轴参与调控肺癌细胞的侵袭和转移。本研究结果表明,HCG11通过靶向下调miR-1297表达从而抑制肺癌细胞的增殖、迁移和侵袭,遏制肺癌的恶性发展。
猜你喜欢
天津医科大学学报(2021年4期)2021-08-21
中日友好医院学报(2021年1期)2021-04-14
山东医药(2020年9期)2020-05-20
中成药(2017年9期)2017-12-19
中成药(2017年5期)2017-06-13
中国药理学通报(2015年2期)2016-01-12
华南农业大学学报(2015年5期)2015-12-04
热带农业科学(2015年9期)2015-10-14
中国医学科学院学报(2015年5期)2015-03-01
中国医药导报(2015年27期)2015-02-28
