
热暴露对大鼠胸主动脉内皮细胞钟基因BMAL1 和细胞周期蛋白的影响
2021-06-22 08:28张瀚文常笑语杨春丽武晓敏李光华
宁夏医科大学学报 2021年6期
张瀚文 , 耿 瑶 , 常笑语 , 穆 乐 , 杨春丽 , 武晓敏 , 李光华 ,
(1.宁夏医科大学基础医学院生理学系,银川 750004; 2.宁夏医科大学护理学院,银川 750004;3.宁夏医科大学公共卫生与管理学院,银川 750004)
应激是机体在各种内外环境因素及社会、心理因素刺激后所做出的一系列非特异性生理反应的总和。长时间处于高温环境,可引起全身的非特异性适应反应,从而引起各种病理生理学变化。热应激可导致大鼠小肠发生病理性损伤、昼夜节律相关基因mRNA 表达异常,可诱导细胞凋亡并导致血管损伤,影响心血管功能[1]。许多重要的细胞周期基因,如 p53、p21、wee1、c-myc、cyclind1 等都是钟控基因,其表达受钟基因调控,具有昼夜节律性。细胞周期正常有序的运转过程受到许多细胞周期基因和其产物的调控,这些调控因子构成了一个复杂且有序的分子网络,监管着细胞周期进程有序运转,以确保细胞增殖和凋亡的平衡。当这些细胞周期的调节因子发生异常和改变时,导致细胞周期紊乱,继而使细胞增殖和凋亡平衡失控最终导致疾病的发生[2-6]。生物钟和细胞周期之间存在紧密的双向相位耦合[7]。研究[8-9]表明,钟基因在人类口腔黏膜和皮肤中的表达与特定的细胞周期时相有关,CyclinB1 基因Cnnb1 的峰值表达与BMAL1 钟基因的峰值一致,而PER1 的转录与P53 mRNA 水平的峰值在G1 晚期一致。为探讨长时间热暴露对动物昼夜节律及细胞周期潜在的影响,本文对大鼠热暴露后胸主动脉和内皮细胞昼夜节律相关基因 BMAL1 和细胞周期蛋白 CDK1、CDK4、CDK6、CyclinB1 表达进行研究,进一步探讨此现象发生的机制。
1 材料与方法
1.1 实验动物、细胞、试剂及仪器
选用健康雄性SD 大鼠20 只,体质量180~220 g。所有大鼠每日自由进食[动物及饲料均由宁夏医科大学实验动物中心提供,伦理号:宁医大伦理第 2020-326 号;许可证号:SCXK(宁)2015-0001],并给予充足的纯净水饮用,分笼饲养,每笼5 只,光照明暗周期为12 h/12 h,即每天6:00~18:00 为暗,18:00~次日 6:00 为明,环境温度(24±0.1)℃。体外实验,选用大鼠胸主动脉内皮细胞株(RTAEC),购自上海赛齐生物工程有限公司,置于37 ℃、5%CO2的培养箱中,进行常规细胞培养。
WB 一抗:BMAL1(货号:DF10308-200;美国Affinity Biosciences 公司),CyclinB1(货号:#4138;Cell Signaling Technology 公司),CDK1(货号:bs-0542R;北京博奥森生物技术有限公司),CDK4(货号:11026-1-AP)、CDK6(货号:14052-1-AP)均购自美国 Proteintech Group 公司,WB二抗:辣根酶标记山羊抗小鼠IgG(货号:ZB-2305)、β-Actin(货号:TA-09)均购自北京中杉金桥生物技术有限公司,Goat Anti-rabbit IgG/HRP(货号:bs-0259G-HRP;北京博奥森生物技术有限公司)、BCA 蛋白含量检测试剂盒、全蛋白提取试剂盒(KeyGEN BioTECH),光学显微镜(日本奥林巴斯公司),人工气候舱、酶标仪、4 ℃离心机、超净工作台、37℃二氧化碳细胞培养箱(美国赛默飞世尔科技有限公司),电泳仪、电泳槽(美国Bio-Rad 公司),震荡混悬器(江苏海门其林贝尔仪器制造有限公司),-80 ℃冰箱、-150 ℃超低温冰箱(美国-纽艾尔公司),化学发光成像仪(美国通用电气公司),细胞培养瓶(美国 Corning 公司)。
1.2 实验动物的分组和干预方法
将20 只SD 大鼠,随机分为正常对照组和热暴露组各10 只,环境温度为24 ℃,适应性饲养1 周;开始模型制备;正常对照组,大鼠饲养在24 ℃温度环境中;热暴露组大鼠饲养在32 ℃温度环境中;两组饲养环境相对湿度均为(32±4.3)%,干预过程中,每3 d 进行1 次体质量测量并记录,持续时间为14 d。两组大鼠分别干预14 d 后,在取材前夜禁食,自由饮水;取材前对大鼠进行称重,戊巴比妥钠配制成1%生理盐水溶液腹腔注射对大鼠进行麻醉,给药容积一般为5~10 mg·kg-1,麻醉完成后,将大鼠固定于解剖台上,暴露大鼠的整个胸部和腹部,修剪去腹部毛发,75%乙醇消毒。从剑突向上剪开皮肤至颌下,向两侧钝性分开皮肤与皮下组织,暴露胸廓及颈浅肌层;沿正中剪开胸廓、胸膜,避免损伤胸腺的胸腔器官。暴露心脏,分离其他器官组织,沿着心脏主动脉弓在大鼠胸腔后壁脊柱侧分离胸主动脉,用止血钳夹住胸主动脉上下端,眼科剪剪取胸主动脉,置于生理盐水反复涮洗去除血管中的血液,剪成数段分别进行组织固定及冻存,以备后续实验使用。
1.3 大鼠胸主动脉内皮细胞的培养和处理
复苏胸主动脉内皮细胞株,待其稳定生长,进入对数生长期,对细胞进行干预;对照组置37 ℃、5%CO2的培养箱中,热暴露组置 40 ℃、5%CO2的培养箱中,用含10%新生牛血清以及青链霉素各100 U·mL-1的 DMEM 培养液培养 24 h。
1.4 HE 染色观察热暴露大鼠胸主动脉血管形态结构变化
取新鲜的大鼠胸主动脉血管浸泡在4%的多聚甲醛液中固定24 h,自来水冲洗2 h,70%乙醇中过夜。次日,从75%乙醇捞出组织包埋盒,分别经过80%、90%、95%乙醇2 h、无水乙醇Ⅰ30 min、无水乙醇Ⅱ30 min 依次进行组织脱水。组织包埋盒从无水乙醇中取出,去掉多余乙醇,放入盛有新鲜二甲苯的容器Ⅰ和Ⅱ中各30 min。浸蜡、包埋、切片、脱蜡、脱水、苏木素溶液中进行细胞核的染色、0.5%分化液中(成分:75%乙醇和浓盐酸)分化、伊红染色、滴加中性树脂,封片进行后续拍片观察。
1.5 免疫组织化学染色检测热暴露大鼠胸主动脉BMAL1 蛋白表达
将制成蜡块的大鼠胸主动脉血管进行组织切片,将组织切片迅速放入二甲苯Ⅰ和Ⅱ脱去组织中的蜡,脱水,高温抗原修复,37 ℃温箱内孵育15 min;用PBS 清洗;滴加正常用山羊血清放入湿盒内,37 ℃温箱封闭1 h。滴加一抗置于湿盒内,4 ℃过夜,37 ℃复温 1~2 h,PBS 清洗;甩干滴加反应增强液,37 ℃孵育20 min;PBS 清洗,滴加相应二抗,37 ℃孵育 1 h;PBS 清洗 3 次,进行DAB 显色,去离子水中清洗3~5 min,滴加苏木素染色液中染核2 min;去离子水中清洗,置于0.5%分化液中(成分:75%乙醇和浓盐酸)分化5 s,变色即捞出,放入水中清洗,清洗结束后依次经50%、75%、80%、90%、95%乙醇、无水乙醇Ⅰ、无水乙醇Ⅱ各30 s;最后转入二甲苯Ⅰ和二甲苯Ⅱ各2~3 min;加一滴中性树脂于组织中心,盖上与载玻片相配套的盖玻片,赶尽多余气泡,放于室温下晾干,进行后续拍片观察,用Image J软件对目的蛋白表达阳性细胞数进行统计。
1.6 Western blot 检测热暴露大鼠胸主动脉血管组织和胸主动脉内皮细胞BMAL1 及细胞周期蛋白 CDK1、CDK4、CDK6、CyclinB1 的表达
将制备的组织和细胞从-80 ℃冰箱取出,放冰上备用,提前配制组织和细胞裂解液,放置冰上预冷,组织蛋白提取需要提前用液氮将研钵预冷,将组织置于研钵中,研磨成组织粉末,并转移至离心管中,根据组织重量大小,加入组织裂解液,充分震荡混匀,离心,收集蛋白;细胞蛋白的提取则省去液氮研磨步骤,直接将配制好的细胞蛋白裂解液根据细胞数加入细胞,充分振荡混匀,离心并收集蛋白,然后采用BCA 法进行组织和细胞蛋白含量的测定。最后将蛋白变性后经10%SDS-聚丙烯酰胺凝胶电泳后转移至PVDF 膜上,用含 5%脱脂奶粉的 TBST(Tris-Hcl 20 mmol·L-1,NaCl 137 mmol·L-1含 0.1% Tween-20)封闭 1 h,用特异性Western blot 目的蛋白一抗4 ℃孵育过夜,TBST 洗涤3 次,每次5 min,相应的二抗孵育1 h,TBST 洗涤 3 次,每次 5 min,配制超敏发光液:将超敏发光液试剂盒中的A 液和B 液按1∶1比例配制,每张膜200 μL 的用量按照实际情况配制;发光;用Image J 软件进行目的条带灰度值分析,GraPhPad Prism6 软件进行统计分析并作图。
1.7 统计学方法
运用SPSS 22.0 统计学软件和GraPhPad Prism6软件对数据进行分析。满足正态分布的计量资料以均数±标准差()表示,组间比较采用 t 检验,多组间比较采用单因素方差分析,检验水准取 α=0.05。
2 结果
2.1 热暴露对大鼠体质量的影响
随着时间的变化,两组大鼠的体质量均呈上升的趋势,相同时间点两组大鼠体质量的差异均无统计学意义(P 均>0.05),见表 1。
表1 热暴露对大鼠体质量的影响(,g)
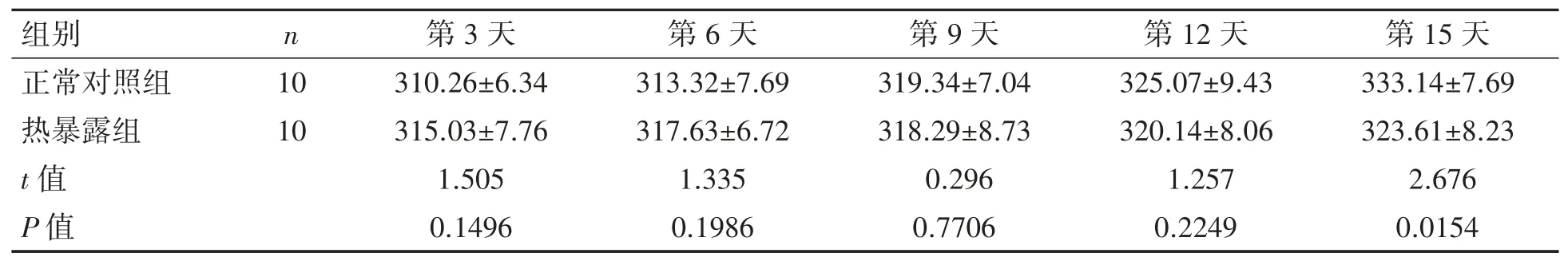
表1 热暴露对大鼠体质量的影响(,g)
组别 n 第3 天 第6 天 第9 天 第12 天 第15 天正常对照组 10 310.26±6.34 313.32±7.69 319.34±7.04 325.07±9.43 333.14±7.69热暴露组 10 315.03±7.76 317.63±6.72 318.29±8.73 320.14±8.06 323.61±8.23 t 值 1.505 1.335 0.296 1.257 2.676 P 值 0.1496 0.1986 0.7706 0.2249 0.0154
2.2 热暴露对大鼠胸主动脉血管形态结构的影响
大鼠胸主动脉血管HE 染色结果显示:正常对照组大鼠胸主动脉血管管壁完整,弹性纤维排列整齐;热暴露组大鼠胸主动脉血管内皮细胞明显缺失,弹性纤维排列紊乱,断裂,不规则,见图1。
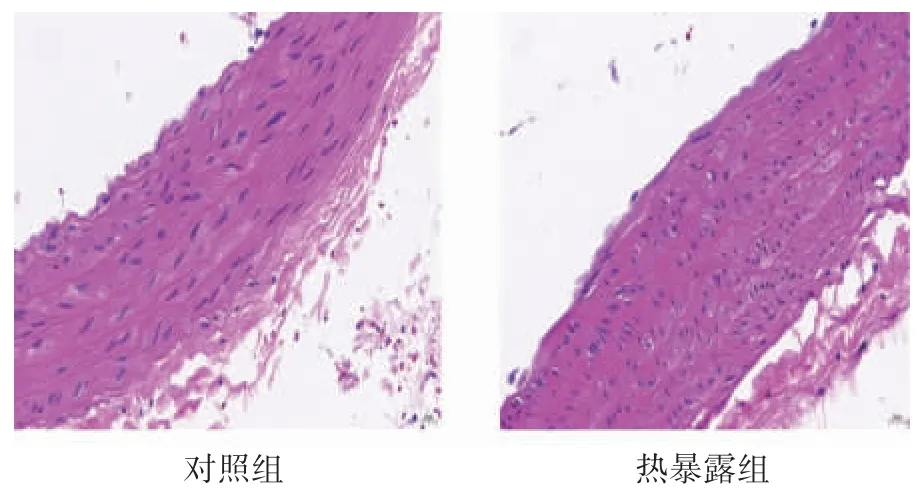
图1 热暴露对大鼠胸主动脉血管形态结构的影响(HE×40)
2.3 免疫组织化学检测热暴露大鼠胸主动脉BMAL1 蛋白的表达情况
棕色颗粒为目的蛋白的表达,颜色越深代表目的蛋白表达越高,颗粒越多表明目的蛋白表达阳性细胞数目越多。与正常对照组比较,热暴露组BMAL1 蛋白表达升高,阳性细胞数目增多(P<0.001),见图 2、图 3。

图2 热暴露对大鼠胸主动脉血管BMAL1 蛋白表达的影响(IHC×40)

图3 两组大鼠胸主动脉血管BMAL1 蛋白表达情况
2.4 Western blot 检测热暴露大鼠胸主动脉血管BMAL1 及细胞周期蛋白 CDK1、CDK4、CDK6、CyclinB1 的表达情况
与对照组比较,热暴露组大鼠胸主动脉血管BMAL1、CDK1、CyclinB1 蛋白表达均升高(P 均<0.05),见图 4。
2.5 Western blot 检测大鼠胸主动脉内皮细胞BMAL1 及细胞周期蛋白 CDK1、CDK4、CDK6、CyclinB1 的表达
与对照组比较,热暴露组大鼠胸主动脉血管组织 BMAL1、CDK1、CyclinB1 蛋白表达均升高(P均<0.01),见图 5。
3 讨论
课题组前期研究[10]表明,热应激可以使胸主动脉血管对去甲肾上腺素(NE)的收缩反应性发生变化,因不同时间热暴露引起的这种血管反应性的变化,推测这种变化可能和钟基因的调控机制相关,血管的收缩反应改变可能与内皮功能改变相关,其作用机制尚未明确。昼夜节律钟是以24 h 周期组织细胞功能的自主计时机制。在哺乳动物中,主要的昼夜节律振荡由转录-翻译反馈环组成[11-13]。BMAL1 是核心钟基因的关键元件,也是缺失后导致哺乳动物所有节律完全消失的惟一时钟基因。钟基因形成转录-翻译反馈环,维持昼夜节律[13]。昼夜节律的紊乱可影响各种行为和生理功能增加各种疾病的风险,当这些昼夜节律振荡器的功能被年龄环境或基因突变所扰乱时,细胞功能的时间协调性就失去了,这就降低了机体的健康和适应能力,引起代谢综合征和心血管疾病等。血管内皮细胞(VECs)是位于心、血管和淋巴管内表面的单层扁平上皮,具有多种生理功能。生物钟功能障碍可引起内皮功能障碍,内皮功能障碍是导致心血管疾病的关键环节,心血管系统又受到昼夜节律分子成分的影响。

图4 大鼠胸主动脉血管BMAL1 及细胞周期蛋白CDK1、CDK4、CDK6、CyclinB1 的表达情况

图5 大鼠胸主动脉内皮细胞BMAL1 及细胞周期蛋白CDK1、CDK4、CDK6、CyclinB1 的表达情况
细胞周期表现为一个振荡器,细胞周期蛋白的循环表达以顺序和单向的方式调节细胞周期进程[14]。细胞周期蛋白在细胞周期的特定阶段产生,并与其各自组成性表达的细胞周期蛋白依赖性激酶(CDK)结合。细胞周期蛋白-CDK 复合物(Cyclin/CDK)的激酶活性在细胞周期的特定时间触发各种事件[15]。各种Cyclin/CDK 复合物的数量和活性的振荡对细胞周期进程至关重要。本研究结果显示,热暴露组胸主动脉血管内皮细胞明显缺失,弹性纤维紊乱、断裂;热暴露组大鼠胸主动脉血管BMAL1 阳性表达增多;体内实验中,热暴露组大鼠胸主动脉血管中BMAL1、CyclinB1、CDK1 蛋白表达升高;体外实验,热暴露组大鼠胸主动脉内皮细胞中 BMAL1、CyclinB1、CDK1 蛋白表达升高。由此推测,热暴露可使钟基因表达水平发生改变,也可以影响细胞周期蛋白的表达,且周期蛋白表达变化比较明显,其中主要是调控G2/M 期的CyclinB1、CDK1。提示热暴露可以引起大鼠胸主动脉内皮损伤及组织结构紊乱;大鼠胸主动脉及大鼠胸主动脉内皮细胞钟基因BMAL1 和细胞周期蛋白表达变化,其主要影响了G2/M 期主要蛋白的表达,对G1/S 期蛋白表达无影响,且在体内外实验结果呈现一致,提示热暴露可以改变钟基因的表达水平,从而影响周期蛋白表达水平,可能通过影响了钟基因表达及G2/M 期蛋白表达来影响胸主动脉及内皮细胞的功能。
综上,热暴露可以引起大鼠胸主动脉内皮结构改变;大鼠胸主动脉内皮细胞钟基因BMAL1和细胞周期蛋白CyclinB1、CDK1 表达上调,提示热暴露可以改变钟基因与细胞周期蛋白的表达水平,此变化可能是通过影响G2/M 期蛋白表达来实现的。
猜你喜欢
体育科技文献通报(2022年4期)2022-10-21
中国医药导报(2022年27期)2022-10-20
中国眼耳鼻喉科杂志(2022年3期)2022-05-29
中国循证心血管医学杂志(2022年1期)2022-03-15
皮肤性病诊疗学杂志(2021年5期)2021-11-27
服饰导报·鞋世界(2021年4期)2021-05-17
三农资讯半月报(2021年1期)2021-01-27
科学大观园(2021年1期)2021-01-11
中国医药导报(2016年33期)2017-03-06
现代养生·下半月(2015年8期)2015-11-16
