
原生质体再生法复壮秀珍菇菌株初探
2021-04-03 08:42丛倩倩王庆武
食用菌 2021年2期
崔 晓 丛倩倩 王庆武
(泰安市农业科学研究院食用菌研究所,山东泰安 271000)
秀珍菇(Pleurotus pulmonarius)学名为肺形侧耳[1],20 世纪后期由我国台湾省引进并推广[2],其营养丰富,蛋白质高于香菇、草菇,且纤维含量少,口感脆嫩鲜美,深受欢迎[3]。秀珍菇的扩大再生产需要菌种的多次扩繁,而多次扩繁会导致菌种退化,子实体的质量和产量下降等问题随之而来。常见的复壮方法如挑取不同部位菌丝、组织分离、改良培养基配方等的效果不佳[4-8],国内外鲜有利用原生质体再生的方法复壮秀珍菇的研究报道。笔者首次通过原生质体再生的方法复壮秀珍菇,旨在验证该方法在秀珍菇菌株复壮的有效性。
1 材料与方法
1.1 供试材料
秀珍菇菌株秀-T。经多次扩繁后,该菌株有母种在平板上出现角变,扩繁种长速变慢甚至停止生长,出菇试验产量下降等退化现象。
1.2 母种培养基配方
PDA 培养基:马铃薯(去皮)200 g,葡萄糖20 g,KH2PO43 g,MgSO41.5 g,琼脂20 g,加水定容至1 L。每个平板定量为20 mL。
TB3 再生培养基:酸水解酪蛋白3 g,酵母提取物3 g,蔗糖200 g,葡萄糖10 g,琼脂8 g,加水定容至1 L。
1.3 原生质体制备与再生
供试秀-T 菌株原生质体制备与再生的方法参考文献[9]。
1.4 秀珍菇栽培试验
以出发菌株为对照,进行复壮秀珍菇菌株栽培试验,每处理重复3次。聚乙烯塑料折角袋装料(约装干料400 g),高压高温(115 ℃)灭菌8 h,待料温降至25 ℃以下,按无菌操作规范两端接种。栽培试验配方及管理参照文献[10]。记录菌丝满袋期、现原基时间、子实体形成时间、第1 潮采收时间,计产并计算生物学效率。
1.5 菌丝长速测定
母种以“十字交叉法”测量平板中菌丝直径并计算菌丝长速,原种及栽培种采用“划线法”测量并计算菌丝长速。每处理重复10次。
1.6 酶活性定义及复壮菌株酶活性测定
测定复壮菌株的羧甲基纤维素酶(CMC 酶)、木聚糖酶、漆酶活性,以出发菌株为对照(CK)。酶活性测定方法参考文献[11]。
酶活性定义:特定条件下,1 L 酶液1 min 内转化生成1 μmol 葡萄糖所需的酶量,称为一个国际单位(U)。酶活性用X表示,单位为酶活性单位每升(U/L)。木聚糖酶、CMC 酶活性计算参照文献[12]。
1.7 标准曲线绘制
(1)葡萄糖标准曲线绘制
准确称取0.1 g 烘干后的葡萄糖,溶于水中,定容至100 mL 备用。将不同浓度的葡萄糖溶液与DNS 试剂反应,530 nm 测OD 值,绘制葡萄糖标准曲线[11]。
(2)木糖标准曲线绘制
准确称取0.1 g 烘干后的木糖,溶于水中,定容至100 mL 备用。将不同浓度的木糖溶液与DNS 试剂反应,530 nm 测OD 值,绘制木糖标准曲线[11]。
1.8 数据处理
相关数据采用Excel 2010 进行整理并进行方差分析。采用Duncan’s 新复极差法统计分析。
1.7 得到葡萄糖标准曲线回归方程为:y=0.9922x+0.0595,R2=0.9924;木糖标准曲线回归方程为:y=0.9106x+0.047,R2=0.9932。
2 结果与分析
2.1 原生质体再生菌株筛选
2.1.1 原生质体再生双核菌株镜检
秀珍菇菌株原生质体再生后,挑取的35个再生菌落中,通过分离培养及镜检获得20个具有明显锁状联合(图1)的双核菌丝体。
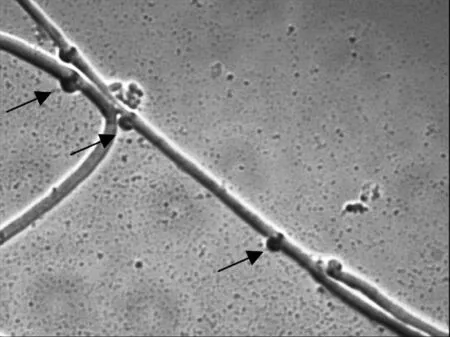
图1 锁状联合结构(箭头所指)
2.1.2 原生质体再生菌株初步筛选
将出发菌株与原生质体再生得到的20 株双核菌株分别接种于平板培养基中观察,结果见表1。
由表1 可知,Re-1、Re-2、Re-5、Re-6、Re-9 这5株再生菌株具有明显生长优势。
2.2 出菇试验
2.2.1 再生菌株扩繁种生长情况
由图2、表2可知,经再生复壮后的菌株,其原种和栽培种的菌丝平均长速均高于出发菌株。原种菌丝平均长速依次为:Re-5>Re-6>Re-2>Re-1>Re-9>秀-T(CK),Re-5 生长最快,满瓶时间比出发菌株早6 d;栽培种菌丝平均长速依次为:Re-6>Re-5>Re-2>Re-9>Re-1>秀-T(CK),且Re-2、Re-5与Re-6 之间差异不显著,Re-2 与Re-5 生长最快,菌丝满袋时间比出发菌株早10.5 d。

表1 异核体分离株的菌丝生物学特性

图2 出发菌株与复壮菌株扩繁种生长情况
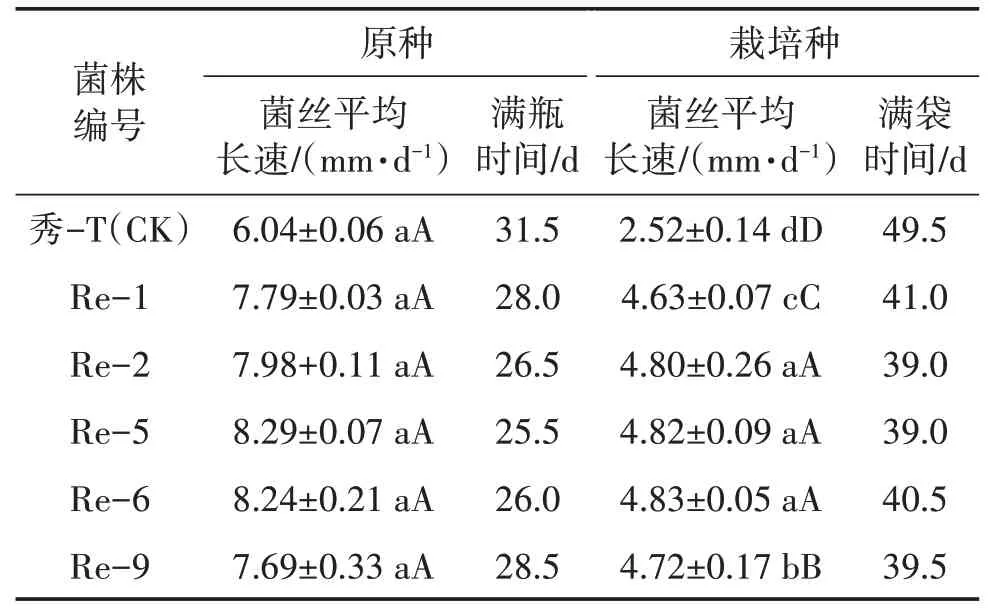
表2 出发菌株与复壮菌株扩繁种菌丝生长情况
2.2.2 再生菌株栽培结果
由图3、表3可知,经再生复壮后的菌株现原基、子实体形成、第1 潮菇采收时间均比出发菌株早。并且再生复壮后的菌株子实体分化快,出菇时间集中,出菇整齐,产量均有所提高。生物学效率由高到低依次为Re-2>Re-5>Re-9>Re-6>Re-1>秀-T(CK),其中Re-2 生物学效率最高,为62.17%,增产27.90%。

图3 出发菌株与复壮菌株子实体栽培出菇情况
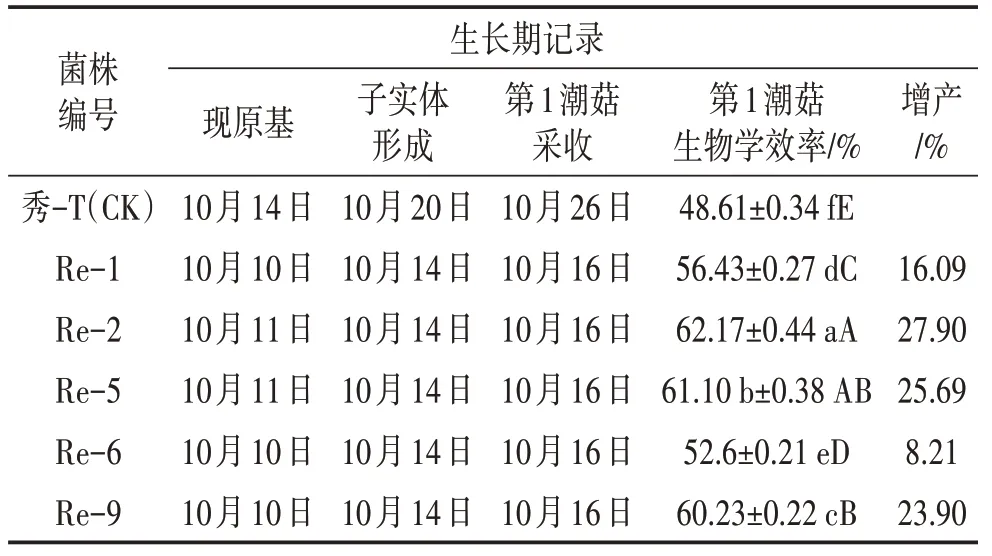
表3 出发菌株与复壮菌株栽培结果
2.3 原生质体再生菌株胞外酶活性
秀珍菇为一种木腐菌,能够分泌一系列的胞外酶降解木质纤维素,这些酶能够通过协同作用将外界的木质纤维素降解成小分子的物质以供自身的利用[13]。结果表明(表4),复壮后的菌株羧甲基纤维素酶、木聚糖酶、漆酶活性均高于出发菌株。复壮后菌株羧甲基纤维素酶活性由高到低依次为Re-2>Re-9>Re-5>Re-6>Re-1>秀-T(CK);木聚糖酶活性由高到低依次为Re-2>Re-9>Re-5>Re-6>Re-1>秀-T(CK);漆酶活性由高到低依次为Re-2>Re-9>Re-6>Re-5>Re-1>秀-T(CK)。
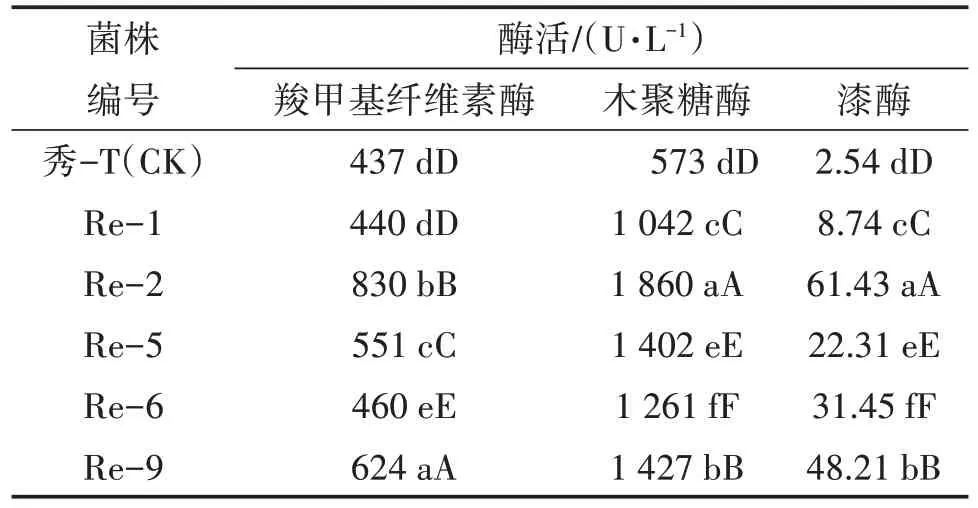
表4 出发菌株与复壮菌株胞外酶酶活
3 小结与讨论
在食用菌的菌种复壮中,采用的方法主要有挑取菌丝尖端法、组织分离法、原生质体再生法等。其中,挑取菌丝尖端法操作简易、技术难度较低,但往往要挑取多次,周期较长,且经出菇验证其复壮效果并不理想。组织分离法需用退化菌株的子实体分离,栽培出菇周期较长,而且一旦菌株退化严重或不出菇影响组织分离。原生质体再生法的缺点是技术要求相对较高,优点是复壮效果好,复壮周期短,减少了后续试验的工作量。
近年来,原生质体再生法在其他食用菌如平菇[14]、茯苓[15]复壮中都取得良好效果。樊晓琳等[4]采用灰色系统关联度分析法将不同方法得到的提纯复壮双孢蘑菇菌株与出发菌株相比较,得出原生质体再生法的提纯复壮效果最好。伍虹蓉[16]利用原生质体再生法得到毛木耳复壮菌株,并且证明其干耳产量同CMC 酶、木聚糖酶活性呈正相关。由此可见,原生质体再生技术已被广泛应用于食用菌菌种复壮,但对秀珍菇鲜有报道。
秦俊哲等研究表明,纤维素酶活性高低可以反映菌株活性的强弱[17],聂荣荣等[18]将漆酶作为金针菇液体菌种活力检测的生物标记物。Ivana Eichlerová 等[19]在研究不同低温存储方式对糙皮侧耳菌种影响时,也是以漆酶作为主要参考指标。试验通过原生质体再生法复壮秀珍菇,得到的再生菌株在平板培养基中菌丝恢复到退化前的长势强、菌丝健壮、菌落边缘整齐的菌丝状态;复壮后菌株菌丝胞外酶活性有所提高;出菇试验中,菇蕾分化及出菇时间提前,生物学效率提高。其中Re-2各方面指标较为理想,可逐步推广。另外,退化的秀珍菇菌株秀-T 的菌丝菌落不规则,存在角变,与缺乏氮源菌丝菌落形态相似[20]。笔者推测秀珍菇秀-T 的退化表现之一为营养元素特别是氮素的吸收不良。由此可见,针对退化秀珍菇的复壮方法,除了原生质体再生法,是否可以通过调节母种或栽培料中配方配比来达到复壮效果(这种复壮方法已有复壮杏鲍菇、草菇报道[7,21]),还有待进一步研究。
猜你喜欢
浙江农业学报(2022年2期)2022-03-03
军事文摘(2021年18期)2021-12-02
福建农业学报(2021年2期)2021-05-31
江苏广播电视报·新教育(2021年34期)2021-01-03
食用菌(2020年3期)2020-12-22
新农业(2020年21期)2020-11-19
农民致富之友(2019年10期)2019-05-22
新农业(2017年6期)2017-07-15
农家顾问(2016年12期)2017-01-06
食品工业科技(2014年23期)2014-03-11
