
miR-27b-3p 对脓毒症肺内皮细胞YAP 信号表达的调控机制研究
2021-03-24 07:22黄爱华
浙江中西医结合杂志 2021年3期
黄爱华 王 旸
败血症是重症监护病房入院的主要原因,其死亡率和发病率均极高。在败血病患者中,内皮细胞活化和功能障碍继发的弥漫性血管渗漏和炎症反应是多器官衰竭的主要原因[1-3]。脓毒症中的内皮激活导致多种促炎细胞因子的产生以及粘附分子选择素(E/P-selectins)和细胞间粘附分子-1(ICAM-1)的上调,导致多形核中性粒细胞大量浸润,过度炎症,组织损伤,并最终导致器官功能障碍[4-5]。Yes 相关蛋白(YAP)是Hippo 信号通路中的关键分子,在多种肿瘤中发挥着促癌作用[6]。此前研究发现,脓毒症模型中,内皮细胞YAP 表达量增加,而内皮特异性缺失YAP 小鼠加重脓毒症中血管功能障碍和弥漫性炎症反应,提示YAP 在脓毒症内皮细胞活化过程中起到负反馈调节的作用[7]。然而脓毒症中YAP 表达量增加的原因尚未明确。研究表明,多种MicroRNA 在脓毒症的多种细胞中发挥着重要的调控作用[8-9]。因此本研究拟探讨脓毒症小鼠miRNA 对YAP 的调控机制。
1 实验材料
1.1 动 物 40 只SPF 级C57BL/6 小鼠(雄性,6~8周龄,体质量20~25g)购买并饲养于杭州医学院实验动物中心,生产许可证号SCXK(浙)2019-0002,使用许可证号SYXK(浙)2019-0011。饲养条件:SPF 级屏障中,白天黑夜各12h,给予充足食物和无菌水。所有操作均符合杭州医学院实验动物伦理委员会审查要求(伦理批号:20200126)。
1.2 试 剂 小鼠肺微血管内皮细胞完全培养基(批号025646)购自武汉普诺赛生物科技有限公司;肿瘤坏死因子-α(TNF-α,批号1321)、白介素-6(IL-6,批号03216、85165)等炎症因子检测试剂盒购自美国R&D Systems。抗体:YAP 兔单抗(批号16651)、ICAM 兔单抗(批号a2365)、选择素(E-selectin)兔单抗(批号a32356)、磷酸化核因子激活的B 细胞的κ-轻链增强p65 亚基(p-p65)兔单抗(批号a21320)、核因子激活的B 细胞的κ-轻链增强p65 亚基(p65)兔单抗(批号a15135)均购自美国CST 公司。甘油醛-3-磷酸脱氢酶(GAPDH)作为内参。mimic NC 及miR-27b-3p mimic(批号15461)购自西格玛奥德里奇(上海)贸易有限公司。
1.3 仪 器 酶标仪SpectraMax 5 购自美谷分子仪器(上海)有限公司;CFX96 Touch 荧光定量PCR 仪,Trans-Blot Turbo 全能型蛋白转印系统均购自伯乐生命医学产品(上海)有限公司;Odyssey TMLi-COR 检测系统购自上海基因有限公司;-80℃超低温冰箱购自美国Thermo fisher。
2 实验方法
2.1 小鼠CLP 模型构建 盲肠结扎穿孔(CLP)模型构建参考文献[10]进行操作。将40 只SPF 级C57BL/6 小鼠按照随机数字表法分为对照组、CLP 组、miRNA 类似物(mimic NC)组、miR-27b-3p 类似物(miR-27b-3p mimic)组,每组10 只。CLP 组、mimic NC 组、miR-27b-3p 组使用三棱针穿刺盲肠轻轻地压缩挤出少量盲肠内容物诱导小鼠脓毒症发生,对照组进行假手术。术后miR-27b-3p 组小鼠予尾静脉注射miR-27b-3p mimic(1×108/mL,200μL/只),mimic NC 组小鼠予尾静脉注射mimic NC 阴性对照物(1×108/mL,200μL/只)。24h 后取小鼠肺组织分离小鼠原代肺内皮细胞进行后续实验。
2.2 小鼠原代肺内皮细胞培养及处理 将上述四组小鼠原代肺内皮细胞提取出来后进行体外培养及后续实验。小鼠原代肺内皮细胞利用小鼠肺微血管内皮细胞完全培养基进行培养,细胞长至90%时传代或铺板。取2~4 代细胞进行后续实验。
2.3 小鼠肺内皮上清TNF-α、IL-6 含量测定 上述四组细胞体外铺板培养24h 后,取上清经过离心,过0.22μm 滤膜,应用酶联免疫吸附试验(ELISA)试剂盒检测上清TNF-α、IL-6 含量,应用SpectraMax 5检测450nm 处吸光度值,取3 次独立重复实验数据进行统计分析。
2.4 qRT-PCR 检测miR-27b-3p、YAP、ICAM、Eselectin mRNA 水平 将上述四组细胞铺板于6 孔板中,24h 后提取细胞总RNA,经过逆转为cDNA后,进行qRT-PCR(荧光定量PCR),qRT-PCR 引物如下:miR-27b-3p(Forward:5' ATGACACCAAGGACCAGAGC 3',Reverse:5' GTGTAAGGACCCATCGGAGA 3');YAP(Forward:5' CAAGAAAGCAGGCTCACAGAA 3',Reverse:5' GCTGGGTGTTAGGGCTTCG 3');ICAM(Forward:5' GAAATGCCACCTTTTGACAGTG 3',Reverse:5' CTGGATGCTCTCATCAGGACA 3');E-selectin(Forward:5' GGCGTTAGAAAGCATCCTTCC 3',Reverse:5' GCAGAGGGCACACTCAAAGT 3')。所有引物均购自南京金斯瑞生物科技有限公司。
2.5 Western blot 法检测YAP、ICAM、E-selectin 及p-p65 蛋白水平 将上述四组细胞铺板24h 后,应用蛋白裂解液裂解细胞蛋白,经过BCA 法测定蛋白浓度后各取20μg/孔进行蛋白免疫印迹(Western blot),经过电泳、转膜、封闭后,于4℃孵育一抗过夜,一 抗 包 括YAP、ICAM、E -selectin、p -p65、p65 及GAPDH。洗去一抗后室温孵育荧光二抗1h。应用Odyssey TMLi-COR 检测系统进行荧光定量检测。
2.6 统计学方法 应用SPSS 20.0 软件进行统计分析,计量资料采用均数±标准差()表示,多组间比较采用单因素方差分析,两两比较采用SNK-q 检验。P<0.05 表示差异有统计学意义。
3 实验结果
3.1 CLP 模型小鼠内皮细胞YAP、ICAM、E-selectin mRNA 表达水平 应用CLP 诱导小鼠脓毒症模型,取小鼠肺组织进行HE 染色,病理观察显示,对照组小鼠肺泡结构基本完整,无炎性细胞浸润。而CLP 组小鼠肺组织具有严重的组织学改变,包括肺泡充血、渗出液和炎性细胞浸润。进一步qRT-PCR 结果显示,与假手术组小鼠比较,CLP 组小鼠肺内皮细胞YAP、ICAM、E-selectin mRNA 表达水平明显升高(P<0.05)。见表1,图1。
表1 各组小鼠肺内皮细胞YAP、ICAM、E-selectin mRNA相对表达量()

表1 各组小鼠肺内皮细胞YAP、ICAM、E-selectin mRNA相对表达量()
注:对照组小鼠进行假手术操作;CLP 组小鼠通过CLP 诱导脓毒症;CLP 为盲肠结扎穿孔;YAP 为Yes 相关蛋白;ICAM 为细胞间粘附分子;E-selectin 为选择素;与对照组比较,aP<0.05
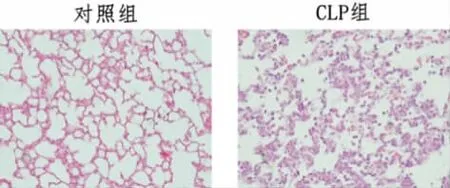
图1 CLP 模型小鼠肺组织形态(HE 染色×400)
3.2 CLP 小鼠肺内皮细胞miR-27b-3p 表达水平miRNA 靶基因数据库PITA 预测调控YAP 的miRNA 为miR-27b-3p,预测结果显示,miR-27b-3p 与YAP mRNA 3' UTR 序列存在6 个完全互补配对碱基UGUGAA。进一步qRT-PCR 检测CPL 组小鼠肺内皮细胞miR-27b-3p 表达,结果显示,与对照组(1.00±0.11)比较,CLP 组miR-27b-3p(0.31±0.15)表达明显下调(P<0.05)。见图2。

图2 miR-328-5p 靶向SPON2 基因3'UTR 序列
3.3 miR-27b-3p 调控YAP 表达及内皮激活 与对照组比较,CLP 组YAP、ICAM、E-selectin 表达水平明显升高,同时p-p65 磷酸化活化水平明显升高;与mimic NC 组比较,miR-27b-3p mimic 组ICAM、Eselectin 表达水平明显升高,同时p-p65 磷酸化活化水平明显升高。见图3。
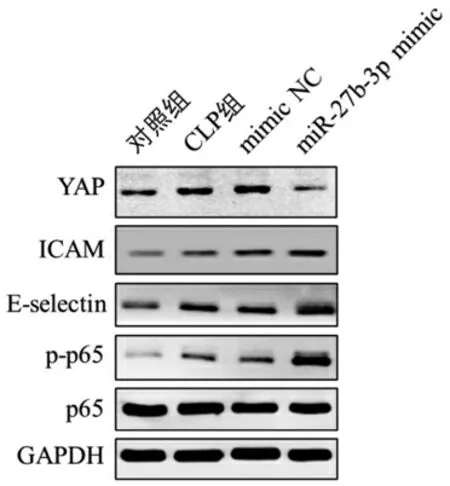
图3 各组小鼠肺内皮YAP、ICAM、E-selectin 表达及p65 活化水平
3.4 miR-328-5p 直接靶向SPON2 抑制其表达 与对照组比较,CLP 组小鼠内皮细胞上清液中TNF-α、IL-6 的含量明显升高(P<0.05);与mimic NC 组比较,miR-27b-3p mimic 组小鼠内皮细胞上清液中TNF-α、IL-6 的含量明显升高(P<0.05)。见表2。
表2 各组内皮细胞上清液TNF-α、IL-6 水平比较(pg/mL,)

表2 各组内皮细胞上清液TNF-α、IL-6 水平比较(pg/mL,)
注:对照组小鼠进行假手术操作;CLP 组小鼠通过CLP 诱导脓毒症;mimic NC 组小鼠通过CLP 诱导脓毒症,同时术后尾静脉注射mimic NC 阴性对照物;miR-27b-3p mimic 组小鼠通过 CLP 诱导脓毒症,同时术后尾静脉注射miR-27b-3p mimic;TNF-α 为肿瘤坏死因子-α;IL-6 为白介素-6;CLP 为盲肠结扎穿孔;与对照组比较,aP<0.05;与mimic NC 组比较,bP<0.05
4 讨论
创伤、休克及感染导致的血管内皮功能障碍和炎症反应是脓毒症发生的主要机制[11-12]。调控内皮细胞异常激活对防止脓毒症发生、降低死亡率具有重要的意义。miRNA 是一类微小RNA,在基因转录调控中发挥着重要的作用。Essandoh 和Fan[13]通过芯片筛选发现,脓毒症组和健康对照组人群之间共有22个差异表达的miRNA,其中9 个表达上调、13 个表达下调。Qiu 等[14]发现长非编码RNA TUG1 通过调控miR-34b-5p 表达,参与脓毒症过程中内皮细胞的活化和炎症反应。本研究发现,miR-27b-3p 在脓毒症内皮细胞中低表达(P<0.05),提示miR-27b-3p 在内皮细胞稳态中可能发挥着重要的调控作用。
YAP 是一种转录共激活因子,通过促进细胞增殖和抑制细胞凋亡而成为组织生长和器官大小的关键调控分子。心脏和血管平滑肌细胞YAP 在心血管发育过程中也起到重要的调控作用[15-16]。而在脓毒症研究中发现,内皮细胞缺失YAP 小鼠内皮细胞异常激活,炎症反应异常加剧[17]。本研究同样发现,脓毒症模型小鼠内皮细胞YAP 表达量明显升高(P<0.05),同时在脓毒症模型小鼠尾静脉注射miR-27b-3p mimic 后,内皮细胞YAP 表达量明显下调(P<0.05)。研究报道,内毒素作用于内皮细胞可以激活打孔蛋白(Gasdermin D),进而释放线粒体DNA 到胞质中,后者作为第二形式参与调控YAP1 信号来抑制内皮细胞炎症反应[18]。然而YAP 调控的炎症下游信号并未明确。基于此,本研究结果显示,miR-27b-3p mimic 组p-p65 亚基磷酸化水平明显升高,下游激活标志物ICAM、E-selectin 表达明显上调,同时内皮细胞分泌的炎症因子TNF-α、IL-6 水平明显上调(P<0.05),表明miR-27b-3p 可能负向调控YAP 表达,参与调控脓毒症过程中内皮细胞的活化以及炎症反应。
综上所述,脓毒症内皮细胞miR-27b-3p 表达下调,通过靶向调控YAP 转录水平,促进炎症相关信号p65 亚基的激活,导致下游内皮细胞活化相关蛋白ICAM、E-selectin 表达增加,加剧脓毒症炎症反应。
猜你喜欢
航天电子对抗(2022年4期)2022-10-24
体育科技文献通报(2022年4期)2022-10-21
中国中医急症(2022年9期)2022-10-12
世界科学技术-中医药现代化(2022年3期)2022-08-22
现代临床医学(2022年2期)2022-04-19
中国药理学通报(2022年1期)2022-01-14
现代临床医学(2021年5期)2021-11-02
滨州医学院学报(2021年2期)2021-05-13
中华养生保健(2020年4期)2020-11-16
中国科技纵横(2018年2期)2018-11-29
