
黄芪甲苷对非小细胞肺癌细胞A549、SPC-A1增殖凋亡的调控作用及其机制
2021-02-24 02:23吴安妮袁力邢婧王楠
山东医药 2021年3期
吴安妮,袁力,邢婧,王楠
中国人民解放军东部战区总医院,江苏南京210000
肺癌是一种实体的恶性肿瘤,其发病率和致死率在众多恶性肿瘤中都位居前列,在世界范围内被认为是人类生命和健康的严重威胁[[1-2]。根据病理特征,肺癌分成了两大类,包括非小细胞肺癌和小细胞肺癌,其中非小细胞肺癌是肺癌主要的一种亚型,占肺癌分型的85%左右[[3-6]。随着基础研究的深入和技术的进步,众多的治疗手段逐步应用于非小细胞肺癌的治疗,包括手术、化疗、免疫治疗、放疗和靶向治疗等[7-8]。目前,非小细胞肺癌对化疗的治疗手段并不敏感,且易产生耐药性,导致非小细胞肺癌患者的5 年生存率仅为10%~15%[9-10]。因此,新药物的研发以及作用机制的阐明对非小细胞肺癌的治疗有着重要的现实意义。
黄芪甲苷是从中药黄芪中提取到的主要活性成分,研究发现其在抗炎、抗氧化、降糖和改善心血管疾病等方面有重要作用[11-13]。近年,人们越来越重视中药在抗肿瘤领域的潜在价值,关于中药抗肿瘤的研究正逐步大规模展开。已有研究表明,黄芪甲苷可有效抑制多种类型肿瘤细胞,如肝癌细胞、人神经母细胞瘤细胞、胃癌细胞等的发育,主要是通过增强体内抑制肺癌的免疫应答而发挥抗肿瘤作用[14-16]。研究发现,黄芪甲苷可以有效抑制肺癌细胞的发育[17-18],但不同浓度条件下黄芪甲苷抗癌的活性以及确切机制有待进一步明确。本研究以非小细胞肺癌细胞A549 和SPC-A1 为体外细胞模型,研究不同剂量的黄芪甲苷对癌细胞增殖和凋亡的调控作用,探究其潜在作用机制,为黄芪甲苷的临床抗非小细胞肺癌的应用提供新的理论依据和潜在的作用靶点。
1 材料与方法
1.1 细胞、试剂、仪器 人非小细胞肺癌细胞A549和SPC-A1均购自中国科学院上海细胞库,体外培养于添加有10%胎牛血清、1%青霉素-链霉素双抗的RPMI-1640 培养液中,在37 ℃、5% CO2的孵育箱内培养。黄芪甲苷(上海源叶生物),用二甲基亚砜(DMSO,Sigma)溶解配成10 mmol/L 的原药贮存液,存于4 ℃冰箱中备用。苯甲基磺酰氟,蛋白酶抑制剂混合液、蛋白磷酸酶抑制剂混合液、高效RIPA 裂解液(上海碧云天公司),一抗兔抗多克隆anti-Ki67、兔抗多克隆anti-PCNA、兔抗多克隆anti-phosopho-Akt(Ser473)、鼠抗单克隆anti-Akt,二抗羊抗兔IgG、羊抗鼠IgG(北京博奥森公司),兔抗多克隆anti-Cleaved caspase9(ASP315)、兔 抗 多 克 隆anti-Cleaved caspase3(Asp175)(美 国Affinity 公 司)。MTT 试剂盒(北京索莱宝),TUNEL 试剂盒(瑞士Roche 公司),BCA 蛋白浓度测定试剂盒(上海碧云天)。多功能酶标仪(美国Biotek 公司),流式细胞仪(美国BD 公司),全自动荧光定量PCR 仪(Applied Biosystems)。
1.2 不同浓度的黄芪甲苷加入后A549 和SPC-A1细胞生长抑制率的测算 采用MTT 法。取对数生长期的A549和SPC-A1细胞,用胰酶进行消化处理,而后用含10%胎牛血清RPMI-1640培养液制备细胞悬液,细胞密度为1×105个/mL。吸取细胞悬液接种到96 孔板中,每孔200 μL。然后将96 孔板放置在37 ℃、5% CO2孵育箱中培养24 h。等到细胞贴壁后,加入0、10、20、40、60、80、100 μmoL/L 的黄芪甲苷替换掉培养液并继续培养24 h。黄芪甲苷处理后,用PBS 进行洗涤,洗涤后加入100 μL 的培养液和20 μL MTT(5 mg/mL),4 h 后终止培养,弃上清液。最后,往每孔中加入150 μL 的DMSO,并将96孔板放置在水平摇床上振荡10 min,在酶标仪上测定490 nm 波长的吸光值(OD)。按下列公式计算生长抑制率:生长抑制率=(1–加药组平均OD 值/对照组平均OD 值)×100%。计算出加入24 h 时黄芪甲苷对A549和SPC-A1细胞增殖的半数抑制浓度(IC50)。
1.3 IC50 的黄芪甲苷加入后A549 和SPC-A1 细胞凋亡率的测算 采用TUNEL 法。取对数生长期的A549和SPC-A1细胞,制备细胞悬液后,按照每孔中1×105个细胞接种到6 孔板中,并放置于37 ℃、5%CO2孵育箱中培养24 h 后分为黄芪甲苷组和空白对照组,等到细胞贴壁后,弃掉培养液,黄芪甲苷组加入IC50 浓度的黄芪甲苷继续培养24 h,随后收集细胞,用PBS 洗涤1 次,再用1 500 r/s 的转速对细胞悬液进行离心5 min,弃上清液。接着加入4%多聚甲醛固定细胞,而后置于摇床上缓慢摇动1 h。最后,用含0.1% Triton X-100 的PBS 重悬细胞,并在冰上孵育2 min,再按照TUNEL 试剂盒说明书采用流式细胞仪检测细胞凋亡率。
1.4 IC50 的黄芪甲苷加入后A549 和SPC-A1 细胞中Ki67、PCNA、Caspase-3、Caspase-9 mRNA 的 检测 采用荧光定量PCR(qRT-PCR)法。将A549 和SPC-A1细胞分为黄芪甲苷组及空白对照组,黄芪甲苷组加入IC50 的黄芪甲苷,空白对照组不加黄芪甲苷,培养24 h后,分别收集各组细胞。利用TRIzol试剂提取细胞的总RNA,利用逆转录试剂盒将总RNA逆转录为cDNA,接着用实时定量PCR 法检测Ki67、PCNA、Caspase-3 和Caspase-9。Ki67 上游引物:5"-GCAGGACTTCACTTGCTTCC-3",下 游 引 物:5"-TCATTTGCGTTTGTTTCACG-3";PCNA 上游引物:5"-AAGGGCTGAAGATAATGCT-3",下 游 引物:5"-AT‑CACCACAGCATCTCCAAT-3;"Caspase-3 上游引物:5"-GGAACGAACGGACCTGTG-3",下 游 引 物:5"-GCCTCCACTGGTATCTTCTG-3";Caspase-9 上 游 引物:5"-AAGCCAACCCTAGAAAACATTACC-3";下游引 物:5"-GACATCACCAAATCCTCCAGAAC-3";GAPDH 上 游 引 物:5"-CTCCTCCTGTTCGACAGT‑CAGC-3",5"-CCCAATACGACCAAATCCGTT-3"。以2-ΔΔCt表示目的基因mRNA表达水平。
1.5 IC50 的黄芪甲苷加入后A549 和SPC-A1 细胞Cleaved-caspase3、cleaved-caspase9、t-Akt、p-Akt 蛋白的检测 采用Western blotting 法。将对数生长期的细胞分别接种于6 孔培养板中(约2×105个/孔),用IC50 的黄芪甲苷(黄芪甲苷组)处理,同时设立空白对照组,处理24 h 时用RIPA 蛋白裂解液(Beyotime)裂解细胞并提取蛋白,接着用BCA 蛋白定量法对提取的总蛋白进行定量。分别取各组60 ng 总蛋白上样,用10%聚丙烯酰胺凝胶进行SDS-PAGE 分析,并将蛋白转移到PVDF 膜上。用TBST 洗涤带有蛋白条带的PVDF 膜后,用5%的马血清室温封闭膜30 min,然后与TBST 中稀释的特异性初级抗体在4 ℃孵育过夜。一抗结束后,用TBST 洗涤2~3 次,每次10 min,随后用二次抗体室温孵育膜2 h。用ECL 发光试剂盒,将所得蛋白进行荧光自显影。用凝胶成像系统记录蛋白电泳条带,利用Image J 软件测量其灰度值,以目的蛋白与内参的灰度值的比值代表目的蛋白的半定量值(将对照组目的蛋白的灰度值与内参的比值设为1)。实验重复3 次,取平均值。
1.6 统计学方法 采用GraphPad Prism 5.0 软件。计量资料以表示,比较采用t 检验。P<0.05 为差异有统计学意义。
2 结果
2.1 不同浓度黄芪甲苷加入后24 h A549、SPC-A1细胞的存活率 不同浓度的黄芪甲苷加入后,A549、SPC-A1细胞的增殖活性均随着黄芪甲苷浓度的升高而逐步降低(图1)。0、10、20、40、60、80 以及100 μmol/L 的黄芪甲苷作用后24 h,A549 细胞的存活 率 分 别100.47% ± 5.37%、88.53% ± 4.22%、73.83% ± 4.93%、37.63% ± 4.73%、26.20% ±5.52%、13.05% ± 3.47%、5.48% ± 2.79%;SPC-A1细胞的存活率分别为102.23% ± 5.75%、94.58% ±4.06%、65.73% ± 3.40%、42.64% ± 6.14%、22.72% ± 4.10%、12.66% ± 5.05%、4.63% ±2.17%。不同浓度的黄芪甲苷加入后,A549 和SPCA1 细胞活力均明显下降(t=3.434,P<0.05)。根据细胞增值活性曲线,计算出黄芪甲苷处理后24 h,A549 和SPC-A1 细胞增殖的半数抑制浓度(IC50)分别为32.6 μmol/L和29.4 μmol/L。
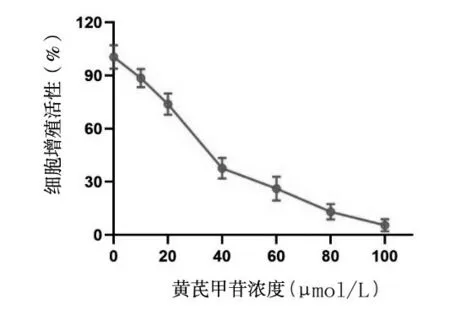
图1 不同浓度黄芪甲苷作用后A549细胞的增殖活性曲线
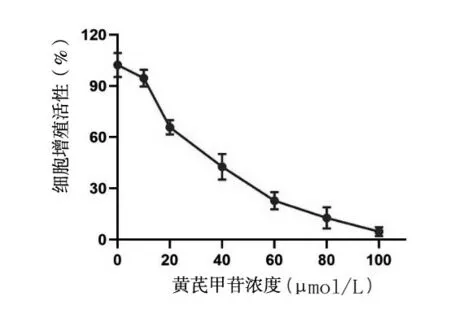
图2 不同浓度黄芪甲苷作用后SPC-A1细胞的增殖活性曲线
2.2 IC50的黄芪甲苷加入后,A549 和SPC-A1细胞凋亡率 IC50 的黄芪甲苷加入后24 h,A549 细胞凋亡率:黄芪甲苷组为29.08% ± 2.49%,空白对照组为8.80% ± 1.47%;,两组比较,t=12.58,P<0.05;SPC-A-1 细胞凋亡率:黄芪甲苷组为25.55% ±2.56%,空白对照组为6.11%±0.88%,两组比较,t=12.44,P<0.05。
2.3 IC50 的黄芪甲苷加入后肺癌细胞A549、SPCA-1 中Ki67、PCNA、Caspase-3、Caspase-9 基因的表达 在A549细胞中,黄芪甲苷组中Ki67 mRNA、PC‑NA mRNA、Caspase-3 mRNA、Caspase-9 mRNA 相对表达量分别为0.727±0.133、0.363±0.090、1.579±0.191、2.052±0.151,空白对照组为1。与空白对照组比较,黄芪甲苷组Ki67 mRNA、PCNA mRNA 相对表达量降低,Caspase-3 mRNA、Caspase-9 mRNA相对表 达 量 升高(t 分别为3.51、10.82、4.875、12.08,P 均<0.05)。SPC-A-1 细胞中,黄芪甲苷组中的Ki67 mRNA、PCNA mRNA、Caspase-3 mRNA、Caspase-9 mRNA 相对表达量分别为0.527±0.043、0.483 ± 0.076、1.287 ± 0.162、1.752 ± 0.171,空白对照组为1。与空白对照组比较,黄芪甲苷组Ki67 mRNA、PCNA mRNA 相对表达量降低,Caspase-3 mRNA、Caspase-9 mRNA 相对表达量升高(t 分别为19.07、8.591、2.965、7.565,P均<0.05)。
2.4 黄芪甲苷加入后,非小细胞肺癌细胞Cleaved‑caspase3、Cleaved-caspase9、t-Akt、p-Akt 蛋 白 表 达非小细胞肺癌细胞A549 中Cleaved-caspase3、Cleaved-caspase9、t-Akt、p-Akt 分 别 为0.845 ±0.136、1.467 ± 0.123、0.902 ± 0.071、0.474 ±0.039。 非小细胞肺癌细胞SPC-A1 中,Cleavedcaspase3、Cleaved-caspase9、t-Akt、p-Akt 分 别 为0.924 ± 0.156、1.578 ± 0.135、1.036 ± 0.052、0.635±0.072。在非小细胞肺癌细胞A549中,与空白对照组比较,黄芪甲苷组Cleaved-caspase3 和Cleaved-caspase9 的蛋白表达量均增加(t 分别为11.110 和5.423,P 均<0.05),p-Akt 降低(t=16.93,P<0.05),t-Akt 差异无统计学意义(t=1.549,P>0.05)。在非小细胞肺癌细胞SPC-A1 中,与空白对照组比较,黄芪甲苷组Cleaved-caspase3 和Cleavedcaspase9 的蛋白表达量均增加(t 分别为10.180、7.418,P<0.05),p-Akt 降低(t=9.025,P均<0.05),t-Akt差异无统计学意义(t=1.478,P>0.05)。
3 讨论
肺癌是全球范围内公认的发病率和致死率最高的恶性肿瘤之一。非小细胞肺癌患者的5年生存率不高,因而研究新型抗肿瘤药物用于治疗非小细胞肺癌具有非常重要的现实意义。中药黄芪提取物中含有一种主要的活性成分——黄芪甲苷,研究表明黄芪甲苷在抗炎、抗氧化、降糖和改善心血管疾病等方面扮演了重要的角色[11-13];同时,近期研究表明,黄芪甲苷在抗肿瘤方面也具有显著性的抑制作用[15-16,18],黄芪甲苷可能作用的信号通路是Wnt/βcatenin 信号通路[19]。黄芪甲苷抑制卵巢癌SKOV3细胞的体外增殖和迁移、侵袭能力,可能与其下调MMP2、MMP9 蛋白的表达有关[20]。目前,黄芪甲苷对非小细胞肺癌细胞的调控作用还不十分明确,作用机制还有待深入探究。本研究中,首先我们利用MTT 分析了不同剂量的黄芪甲苷对A549 和SPC-A1细胞增殖活性的影响,确定了黄芪甲苷处理后24 h的IC50 分别为32.6 μmol/L(A549 细胞)和29.4 μmol/L(SPC-A1 细胞),明确了黄芪甲苷对A549 和SPC-A1 细胞增殖的抑制作用具有剂量依赖性。同时,我们利用TUNEL 法检测了黄芪甲苷对A549 和SPC-A1 细胞凋亡的调控作用,研究发现,黄芪甲苷可促使A549和SPC-A1细胞凋亡。为了进一步确认黄芪甲苷抑制A549 和SPC-A1 细胞的增殖,促进其凋亡,我们还检测了增殖相关基因Ki67、PCNA 和凋亡相关基因caspase-3、caspasse-9的mRNA 和蛋白的表达量。研究发现,在黄芪甲苷处理的细胞中,Ki67和PCNA 的表达水平有显著性下降;而caspase-3 和caspasse-9的表达水平则是显著性上调,进一步表明黄芪甲苷可以抑制A549 和SPC-A1 细胞的增殖,促进A549和SPC-A1细胞的凋亡。
Akt 信号通路是调控细胞增殖和生长的关键信号通路,PI3K在Akt的上游,生长因子可以通过激活PI3K 信号转导通路诱导Akt 激活为磷酸化状态,即p-Akt,来调控细胞增殖。已有相关研究结果显示,黄芪甲苷通过抑制PI3K/AKT 信号通路,抑制DNA损伤修复,诱导肝癌细胞凋亡,使细胞周期G2/M 期发生阻滞,可以使细胞微核率增加,发挥对肝癌细胞的放射增敏作用[21]。本研究中,我们通过免疫印迹法检测了t-Akt 以及p-Akt 的表达水平,发现在黄芪甲苷处理后的A549 和SPC-A1 细胞中,t-Akt 的表达没有发生明显的改变,而p-Akt 的表达水平有显著性的下降,提示黄芪甲苷对A549 和SPC-A1 细胞的增殖和凋亡的调控作用可能由Akt 信号通路介导的。
综上所述,黄芪甲苷可以抑制非小细胞肺癌细胞A549 和SPC-A1 的增殖,促进A549 和SPC-A1 的凋亡,且调控作用具有剂量依赖性。黄芪甲苷对A549 和SPC-A1 细胞的增殖和凋亡调控作用可能是通过抑制Akt信号通路中p-Akt的表达量来实现的,这将为非小细胞肺癌的治疗提供新的理论依据和潜在作用靶点。本课题组将进一步针对黄芪甲苷对Akt 信号通路的下游机制进行验证并探讨黄芪甲苷治疗非小细胞肺癌的潜在通路。
猜你喜欢
新农业(2022年21期)2022-11-18
浙江中西医结合杂志(2022年10期)2022-10-25
中日友好医院学报(2022年4期)2022-10-15
今日农业(2022年13期)2022-09-15
世界最新医学信息文摘(2021年82期)2021-07-13
Digital Chinese Medicine(2020年4期)2020-12-11
看世界·学术下半月(2020年7期)2020-09-10
安徽中医药大学学报(2020年2期)2020-04-21
中国老年学杂志(2019年20期)2019-10-17
海峡姐妹(2019年3期)2019-06-18
