
抗伯氏疟原虫GAP40蛋白免疫血清对动合子发育抑制作用的研究*
2021-02-18 08:27朱俐颖李思琦陈露萌曹雅明朱晓彤
寄生虫与医学昆虫学报 2021年4期
何 璐 朱俐颖 李思琦 陈露萌 曹雅明 朱晓彤
(中国医科大学基础医学院免疫学教研室,沈阳 110122)
疟疾是由媒介按蚊传播的寄生原虫感染性疾病。WHO疟疾报告显示:2019年全球有2.29亿例疟疾病例,40.9万人死于疟疾感染(WHO, 2020)。随着蚊帐、杀虫剂和抗疟药物的应用,疟疾的发病率和死亡率逐年下降,但无症状感染者对全球疟疾防控仍具有极大威胁。阻断疟疾从宿主到蚊虫的传播,是消除疟疾的一大挑战(Arakawaetal., 2014)。传播阻断疫苗(transmission blocking vaccine, TBV)通过提高人群免疫力,产生特异性抗体,抑制疟原虫在蚊体内发育过程,阻断蚊媒传播,形成对疟疾传播的控制。目前已发现多种具有传播阻断能力的疟原虫有性阶段特异性抗原,如P48/45、P47、P230、P25/28、HAP2/GCS1、PbPH和PSOP25(Saxenaetal., 2007; Blagboroughetal., 2009; Caoetal., 2016; MacDonaldetal., 2016)。然而,上述传播阻断候选抗原诱导产生的抗体均不能完全阻断疟原虫传播。因此,探索新型疟疾传播阻断候选抗原对于疟疾的防治仍具有重要意义。
裂殖子、动合子和子孢子发育阶段的原虫具有运动能力。疟原虫的运动与侵袭依赖顶复门生物独特的滑行运动(Dobrowolskietal., 1997)。滑动体glideosome(Opitzetal., 2002))在滑行运动中为疟原虫提供动力(Dobrowolskietal., 1997)。多项研究发现,滑动体由肌球蛋白 A(Myosin A,MyoA)、MyoA尾区相互作用蛋白(MyoA tail domain-interacting protein,MTIP)、滑行相关蛋白(gliding-associated protein, GAP)GAP40、GAP45、GAP50 等组成(Gaskinsetal., 2004; Baumetal., 2006; Frénaletal., 2010a)。MTIP蛋白通过N端与GAP45蛋白相互作用,并连接GAP40和GAP50蛋白,锚链MTIP-ELC-MyoA复合体至内膜复合体(Inner membrane complex,IMC)(Bergman, 2003; Pazickyetal., 2020)。有性发育阶段的动合子可通过滑行运动穿越蚊胃壁形成卵囊,并发育成子孢子,随蚊虫叮咬进入人体,导致疟原虫的传播。基于滑动体在动合子迁移和侵袭中的重要作用,滑动体相关蛋白可作为传播阻断疫苗研究的潜在靶点。在3种滑行相关蛋白中,GAP40和GAP45为原虫红内期生长必需蛋白。而GAP40蛋白在有性发育阶段中的功能作用尚无研究报道。
本研究利用生物信息学分析,选取伯氏疟原虫GAP40(PlasmoDB ID: PbANKA_1115300,PbGAP40)优势抗原表位片段(329~458 aa)构建表达载体,诱导表达重组蛋白,制备鼠源性多克隆抗体免疫血清,并检测血清中特异性抗体滴度及特异性。采用IFA检测PbGAP40蛋白在动合子期原虫中的亚细胞定位。同时,评估抗-PbGAP40免疫血清的传播阻断活性。本研究旨在为新型传播阻断疫苗的研发提供理论基础。
1 材料与方法
1.1 实验材料
6~8周龄雌性BALB/c小鼠(北京华阜康公司),PbANKA疟原虫株Plasmodiumberghei(日本爱媛大学惠赠);EasyGeno单片段重组克隆试剂盒、琼脂糖凝胶DNA回收试剂盒、质粒小提试剂盒、BabyBio Ni-NTA和增强型HRP-DAB底物显色试剂盒购自天根公司;限制性内切酶Fast DigestBamHⅠ和Fast DigestNotⅠ、6×His-Tag单克隆抗体、Goat anti-Mouse IgG(H+L) Secondary Antibody、HRP、ProLongTMGold Antifade Mountant with DAPI、4×LDS Sample Buffer、Alexa FluorTM488 goat anti-mouse IgG(H+L)、Alexa FluorTM594 goat anti-mouse IgG(H+L)、1640培养基和胎牛血清均购自ThermoFisher公司;氨苄青霉素、PVDF膜、LB Broth培养基、Tween-20和IPTG购自北京鼎国公司。
1.2 生物信息学分析
本研究使用的PbGAP40(PlasmoDB ID: PbANKA_111530)基因组序列来自PlasmoDB(http://www.plasmodborg)。利用SMART(http://smart.embl-heidelberg.de/),TMHMM v2.0(http://www.cbs.dtu.dk/services/TMHMM-2.0/),PRALINE(https://www.ibi.vu.nl/programs/pralinewww/)和SYFPEITHI (http://www.syfpeithi.de/0-Home.htm)分别对PbGAP40功能结构域,蛋白相互作用组、跨膜结构域、同源性以及表位进行预测分析。蛋白同源性分析使用BioEdit7.0.9.0绘制,蛋白示意图使用IBS 1.0.3软件绘制。
1.3 pbgpa40基因的扩增和原核表达载体构建
以PbANKA原虫gDNA为模板,使用特异性引物:pET32a-GAP40-BamHⅠ-F:5’-G C C A T G G C T G A T A T C G G A T C C T C T G A A G A T G A A A G A A A A A G A A A A C T GG-3’和pET32a-GAP40-NotⅠ-R:5’-T G G T G G T G C T C G A G T G C G G C C G C T G C T T G A G A A T C A A T A A C C T C C T GC-3’,经PCR反应扩增PbGAP40蛋白329~458 aa片段的编码基因,获得的目的片段纯化后连接至pET32a(+)原核表达载体并转化。通过测序和酶切鉴定选取阳性克隆后进行重组蛋白诱导表达。
1.4 重组蛋白的表达、纯化与实验动物免疫
选取正确克隆的pET32a(+)-PbGAP40菌种,使用1 mmol/L IPTG诱导蛋白表达,19 ℃摇菌过夜表达重组蛋白。次日收集菌液超声裂解,收集裂解液,BabyBio Ni-NTA磁珠纯化重组蛋白。将提纯后的重组蛋白与4×LDS Sample Buffer混合均匀,100 ℃ 3 min煮样后,进行聚丙烯酰胺凝胶电泳分离蛋白,并通过考马斯亮蓝染色检测重组蛋白纯度。同时,SDS-PAGE分离后的重组蛋白,采用半干法转膜:25 V 30 min至PVDF膜,5%脱脂奶粉室温封闭1.5 h,一抗6×His Tag 单克隆抗体(1∶2 000)4 ℃过夜孵育。采用Goat anti-Mouse IgG(H+L) Secondary Antibody(HRP)(1∶20 000)室温孵育1 h。ECL发光方法检测重组蛋白表达情况。
选取20只6~8周龄的雌性BALB/c小鼠,随机分为2组:PBS对照组和PbGAP40-his重组蛋白免疫组。初次免疫采用完全弗氏佐剂与重组蛋白(或PBS)1∶1充分混合,50 μg/只,皮下注射。二免和三免为不完全弗氏佐剂和重组蛋白(或PBS)1∶1充分混合,每次免疫间隔14 d。三免后第10天收集小鼠血清,-80 ℃保存。
1.5 ELISA法检测免疫血清效价
取10 μgPbGAP40重组蛋白加入96孔酶标板,室温过夜包被。每孔加入100 μL含1% BSA的PBS缓冲液封闭,37 ℃孵育l h。将待检测免疫血清倍比稀释后,每孔加入100 μL,37 ℃孵育 2 h。PBS-T洗板。加入 HRP标记的羊抗鼠IgG抗体(1∶5 000),每孔100 μL,37 ℃孵育1 h。PBS-T洗板。底物邻苯二胺和过氧化氢,显色5 min,加入2 mol/L H2SO4终止反应。采用Bio-Rad酶标仪检测450 nm处OD值。
1.6 伯氏疟原虫动合子体外培养
选取6~8周龄BALB/c小鼠,感染前2 d,小鼠腹腔注射6 mg/mL苯肼(200 μL/只)。第0天,采用1×107个PbANKA原虫,腹腔感染BALB/c小鼠。感染后第3天,口服给予磺胺(20 mg/L)48 h后,杀鼠取血与动合子培养基(RPMI1640,100 mg/L新霉素,50 mg/L链霉素,50 mg/L青霉素,20% (V/V) 胎牛血清,pH 8.3)以1∶9比例混合,19 ℃体外培养24 h获得动合子期原虫。
1.7 Western blot检测免疫血清特异性
采用62%的Nycodenz纯化收集体外培养的PbANKA虫株动合子期原虫,提取蛋白并测定浓度。将原虫蛋白与4×LDS Sample Buffer混合,在100 ℃水浴锅煮3 min变性后,进行SDS-PAGE电泳分离。25 V 30 min半干法转至PVDF膜上,以5%脱脂奶粉室温封闭1.5 h。一抗:抗- PbGAP40免疫血清(1∶200稀释),4 ℃过夜孵育。二抗:HRP标记的山羊抗小鼠IgG抗体(1∶20 000), ECL发光方法检测抗-PbGAP40免疫血清的特异性。
1.8 IFA检测PbGAP40蛋白在动合子期的亚细胞定位
收集PbANKA动合子期原虫,4%多聚甲醛/0.0075%的戊二醛室温固定30 min。PBS洗涤3次后,使用0.1 mg/mL硼氢化钠中和10 min。PBS洗涤3次后,采用0.1%Triton-100冰上透膜10 min,5%脱脂奶粉室温封闭1 h。采用抗-PbGAP40抗体(1∶200)室温孵育1 h。PBS洗涤5次。采用Alexa FluorTM488 goat anti-mouse IgG(H+L)(1∶500)避光室温孵育样本1 h。1 μg/mL DAPI染核10 min后封片。采用共聚焦显微镜检测PbGAP40在动合子期原虫的定位情况。
1.9 抗-PbGAP40免疫血清对疟原虫有性发育阶段抑制效果的检测
选取6~8周龄BALB/c小鼠,感染前2天,小鼠腹腔注射6 mg/mL苯肼(200 μL/只)。第0天,采用1×107个PbANKA原虫,腹腔感染BALB/c小鼠。感染后第3天,口服给予磺胺(20 mg/L)48 h后,取10 μL小鼠尾血与90 μL动合子培养基,及1∶5稀释的抗-PbGAP40免疫血清混合均匀,19 ℃ 24 h孵育后涂片,采用抗-Pbs21抗体(1∶500)染色后,计数动合子数目和动合子转化率。
1.10 统计学分析
实验数据采用GraphPad Prism 7.01软件进行分析绘图和统计学分析。血清抗体滴度、动合子数目和动合子转化率采用Student′sttest进行比较;P< 0.05提示组间差异显著。
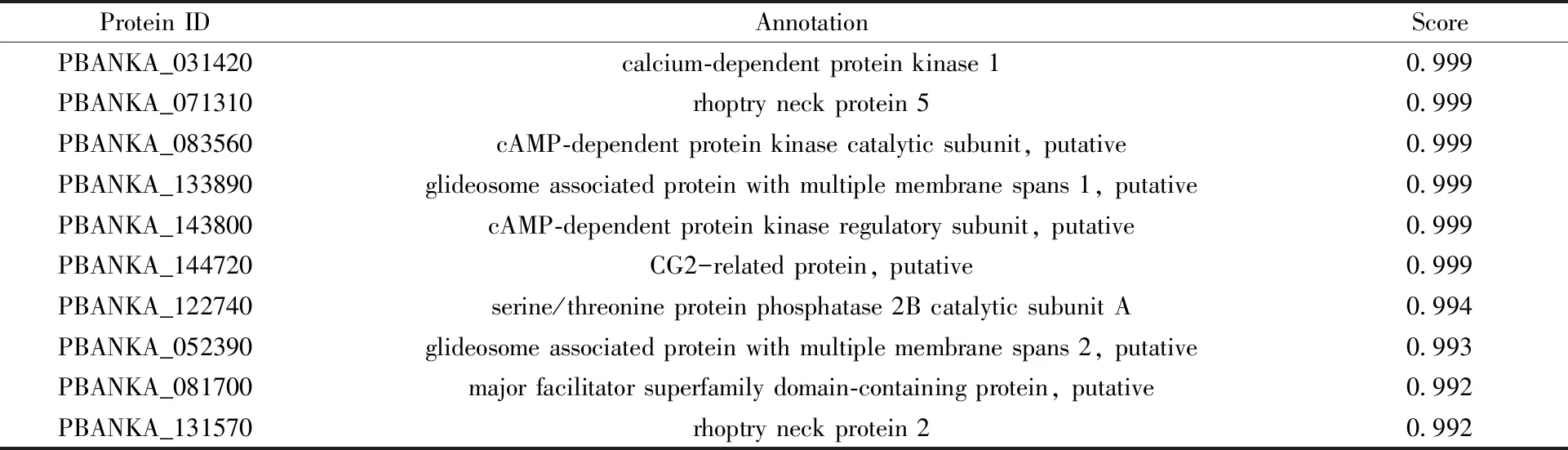
表1 PbGAP40相互作用蛋白预测结果Tab.1 Putative interactome of PbGAP40
2 结果
2.1 PbGAP40蛋白的生物信息学分析
通过SMART软件预测绘制PbGAP40蛋白(PlasmoDB ID: PBANKA_1115300) 的模式图(图1-A),并根据SYFPEITHI软件分析结果,选取伯氏疟原虫GAP40优势抗原表位(329~458 aa) 构建重组蛋白表达载体(图1-A)。对GAP40蛋白进行进化树分析(图1-C),使用PRALINE对伯氏疟原虫GAP40蛋白、约氏疟原虫GAP40蛋白(PlasmoDB ID: PY17X_1116500) 和夏氏疟原虫GAP40蛋白(PlasmoDB ID: PCHAS_1114900) 进行同源性分析(图1-B),3种蛋白具有高度同源性(全长同源性97%,329~458 aa同源性96%)。同时,通过STING软件分析预测出PbGAP40与多种参与动合子滑行运动的蛋白存在相互作用(表1)。
2.2 PbGAP40重组蛋白的表达和纯化及抗-GAP40免疫血清抗体滴度检测
本研究成功构建pET32a(+)-PbGAP40329-458表达载体,并诱导PbGAP40重组蛋白的表达。蛋白经BabyBio Ni-NTA磁珠纯化后,对洗脱及透析后样品进行检测。考马斯亮蓝染色结果显示:PbGAP40重组蛋白纯度>95%(图2-A)。抗-his-tag抗体检测显示:重组蛋白大小约38 kDa(图2-B),符合预期目的蛋白分子量。

图2 PbGAP40重组蛋白表达检测Fig.2 Detection of the PbGAP40 recombinant protein expressionA. PbGAP40-his重组蛋白考马斯亮蓝染色结果; B. Western Blot检测PbGAP40重组蛋白。A.The coomassie blue staining results of PbGAP40-his recombinant protein; B.Western blot detection of PbGAP40 recombinant protein.

图3 ELISA方法检测抗-PbGAP40重组蛋白免疫情况和三免后的血清抗体滴度Fig.3 The immunity of PbGAP40 recombinant protein and antibody titer after immunizationdetected by ELISA analysisA. 抗-PbGAP40重组蛋白免疫情况; B. 三免后的抗-PbGAP40免疫血清抗体滴度。A. The immunity of PbGAP40 recombinant protein; B. The antibody titer of anti-PbGAP40 sera post third immunization. *:P < 0.05;**:P < 0.01; ***:P < 0.001; ****:P< 0.0001.
2.3 抗-GAP40免疫血清中特异性IgG抗体的ELISA检测
BALB/c小鼠经PbGAP40-his重组蛋白3次免疫后,与对照组(PBS)相比,血清中特异性抗体水平显著升高(P<0. 001,图3-A),且ELISA结果显示,抗-PbGAP40免疫血清的抗体滴度为1∶51200(图3-B)。
2.4 PbGAP40蛋白在伯氏疟原虫动合子期的表达和亚细胞定位检测
Western blot实验结果显示,抗-PbGAP40免疫血清能检测出动合子期内源性GAP40蛋白,具有良好的免疫反应性。蛋白大小约为52 kDa, 符合预期大小上(图4-A)。IFA结果显示:GAP40蛋白荧光信号在伯氏疟原虫的合子、retort和成熟的动合子阶段均有表达,且GAP40蛋白的荧光信号定位在原虫质膜(图4-B)。
2.5 抗-PbGAP40免疫血清对伯氏疟原虫动合子发育的抑制情况
使用含有抗-PbGAP40免疫血清(1∶5倍稀释,GAP40组)的动合子培养基进行伯氏疟原虫体外动合子培养24 h后,与PBS免疫血清处理组(PBS组)相比,动合子形成数减少了36.5%(P<0.05,图5-A),动合子转化率下降8.5%(P<0.05,图5-B)。GAP40处理组中,“retort”发育阶段(未成熟动合子)原虫的比例占61.2%,成熟动合子比率仅占27.18%(图5-C)。上述结果提示,抗-PbGAP40免疫血清可在体外抑制动合子的形成和转化。

图4 PbGAP40蛋白在动合子期原虫的表达和亚细胞定位Fig.4 The subcellular localization of PbGAP40 protein in ookinetesA. Western blot检测动合子期PbGAP40蛋白的表达;B. IFA检测PbGAP40蛋白在动合子期的亚细胞定位。Zygote: 合子;Ookinete: 动合子。A. Western blot detection of PbGAP40 in ookinetes; B. IFA detection of the subcellular localization of PbGAP40 protein in ookinetes.
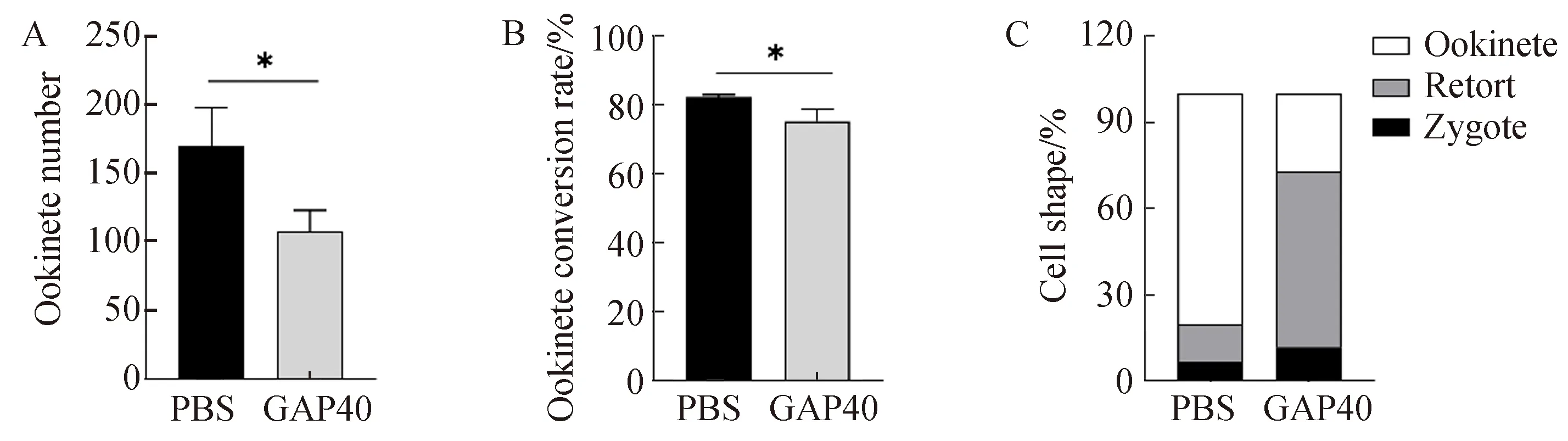
图5 抗-PbGAP40免疫血清动合子发育的影响Fig.5 Effect of anti-PbGAP40 serum on ookinete developmentA. 抗-PbGAP40免疫血清对动合子形成数的影响; B.抗-PbGAP40免疫血清对动合子转化率的影响; C. 抗-PbGAP40免疫血清对合子、retort和动合子占比的影响。A. The effect of anti-PbGAP40 serum on ookinete number; B. The effect of anti-PbGAP40 serum on ookinete conversion rate; C.The Effects of anti -PbGAP40 immune serum on the proportion of zygote, retort and ookinete. PBS: PBS immunized group; GAP40: Recombinant Plasmodium bergheiGAP40 protein immunized group.
3 讨论
顶复门原虫特殊的滑行运动由滑动体提供动力,使原虫能够完成迁移、入侵以及逸出感染红细胞等多种运动模式。滑动体相关蛋白GAP40、GAP45和GAP50与肌球蛋白MyoA-MTIP复合体相连,锚定于内膜复合体(inner membrane complex,IMC)膜,组成滑动体(Bergman, 2003)。GAP45横跨原虫质膜和IMC,其N端经肉豆蔻酰化和棕榈酰化后倾向于定位在质膜(Greenetal., 2008; Wrightetal., 2014),C端特定位点的磷酸化则辅助其定位于IMC膜(Gilketal., 2009)。GAP50通过 C端的跨膜螺旋结构与IMC结合,并通过C端6个氨基酸残基与其他滑动体相关蛋白产生相互作用(Boschetal., 2012)。GAP40是近年发现的滑动体相关蛋白,可与MyoA-MTIP/MLC1-GAP45/50复合体共同纯化(Frénaletal., 2010b)。弓形虫GAP40蛋白拥有9个跨膜结构域,广泛地分布在IMC膜,并在IMC的形成和维持其结构稳定中发挥重要作用(Leungetal., 2014),但GAP40蛋白在疟原虫中的作用尚不明确。
本研究中,我们采用原核表达系统成功表达并纯化PbGAP40重组蛋白,通过免疫小鼠获取抗-GAP40蛋白特异性免疫血清。ELISA实验结果提示PbGAP40重组蛋白具有良好的免疫原性,而Western blot结果显示抗-PbGAP40免疫血清可特异性结合内源性PbGAP40蛋白。IFA结果提示PbGAP40蛋白定位于动合子期原虫的表面。多项研究显示,动合子表面蛋白为TBV良好的作用靶位,如:抗PSOP25和PSOP28的抗体均可有效降低蚊胃内卵囊的形成数目(Sattabongkotetal., 2003);抗PSOP12蛋白的免疫血清可减少卵囊密度,且其抑制作用与抗体滴度呈正相关(Salaetal., 2015);已进入Ⅰ期临床试验的Pvs25疫苗可减少 80%的卵囊形成,降低蚊感染率20%~30%(Malkinetal., 2005),然而,上述TBV候选抗原均因免疫原性较差,难以在人体免疫后达到理想的抗体滴度(Duffyetal., 2020)。本研究中,伯氏疟原虫GAP40蛋白在鼠疟模型中显示出良好的免疫原性和免疫反应性,而其动合子表面定位特点提示PbGAP40可作为传播阻断疫苗筛选的潜在靶位。
近年研究发现,条件性敲除滑动体组成蛋白可影响疟原虫红内期生长发育,如:冈地弓形虫GAP40和GAP50蛋白敲减可导致IMC缺失,弓形虫增殖被抑制(Hardingetal., 2016)。条件性敲除恶性疟原虫GAP45蛋白后,疟原虫丧失侵袭宿主红细胞能力,红内期发育被阻断(Perrinetal., 2018)。上述结果均提示滑动体蛋白可作为抗疟治疗的靶点。然而,滑动体蛋白在疟原虫有性发育过程中是否可作为治疗靶位尚未明确。本研究中,我们发现抗-PbGAP40免疫血清的传播阻断效果虽不如PbPSOP25(动合子转化率减少62.5%)等部分已在鼠疟中研究的TBV候选抗原(Zhengetal., 2017),但其免疫血清仍可显著抑制动合子形成数目和转化率。可见,抗-PbGAP40免疫血清对动合子成熟具有显著的抑制作用。该结果为开发GAP40蛋白作为抗疟治疗靶点提供了必要的理论依据。
综上所述,本研究成功制备了抗-PbGAP40免疫血清,对PbGAP40在疟原虫动合子期原虫的定位进行分析,并对抗-PbGAP40免疫血清的传播阻断效应进行了评估。结果表明:PbGAP40重组蛋白具有良好的免疫原性和免疫反应性。抗-PbGAP40 免疫血清具有显著的传播阻断效果,为后续实验中深入探讨抗-PbGAP40免疫血清对蚊体内卵囊和子孢子形成的影响提供了技术基础。也为以滑动体蛋白为靶点开发传播阻断疫苗提供了前期实验基础。
猜你喜欢
现代畜牧科技(2021年4期)2021-07-21
新农业(2020年18期)2021-01-07
发明与创新(2020年5期)2020-12-20
科学导报(2020年69期)2020-11-09
中国兽医杂志(2019年2期)2019-06-25
郑州大学学报(医学版)(2018年5期)2018-10-10
中国感染与化疗杂志(2018年6期)2018-01-19
现代检验医学杂志(2016年3期)2016-11-15
肝博士(2015年2期)2015-02-27
川北医学院学报(2015年5期)2015-02-22
