
KDM2B调控FAK信号通路在卵巢癌细胞骨架排列中的作用*
2021-02-10 06:21张国平顾秀玉翟光华闫美娜南京医科大学姑苏学院南京医科大学附属苏州医院苏州市立医院北区检验科江苏苏州215008
临床检验杂志 2021年12期
张国平,顾秀玉,翟光华,闫美娜(南京医科大学姑苏学院&南京医科大学附属苏州医院&苏州市立医院北区检验科,江苏苏州215008)
卵巢癌是致死率最高的妇科恶性肿瘤,高转移率是卵巢癌致死的主要原因[1]。细胞骨架的破坏可以抑制卵巢癌细胞的迁移及侵袭,进而抑制转移[2]。组蛋白去甲基化酶[lysine(K)-specific demethylase 2B, KDM2B],又名 FBXL10、JHDM1B 或NDY1,是一类重要的组蛋白脱甲基酶,主要发挥组蛋白 H3上赖氨酸4的三甲基化(tri-methylation of lysine 4 on histone H3,H3K4me3)和组蛋白H3上赖氨酸36的二甲基化(dimethylation of histone H3 at K36,H3K36me2)的脱甲基酶活性。研究发现, KDM2B可以通过PI3K/AKT信号通路,调控结肠癌干细胞的干性[3];KDM2B还可以调控前列腺癌细胞的骨架排列,促进其迁移及侵袭[4]。然而,KDM2B在卵巢癌细胞骨架排列中的作用机制尚不明确。本课题组前期研究发现,KDM2B在卵巢癌组织中的表达水平升高,并可促进卵巢癌细胞迁移。本研究拟探讨KDM2B在卵巢癌细胞骨架排列中的调控作用,并进一步分析KDM2B的调控机制。
1 资料与方法
1.1细胞系、仪器及试剂 人胚肾细胞293T由江苏大学医学院临床血液学检验教研室许文荣教授惠赠,人卵巢癌细胞系 HO8910由江苏大学医学院组织胚胎学教研室卢小东教授惠赠。实时荧光定量PCR仪(Light Cycler480 Ⅱ,瑞士Roche公司),超微量核酸分析仪(杭州奥盛仪器公司),化学发光凝胶成像系统(上海天能科技公司),共聚焦显微镜(德国蔡司公司)。KDM2B过表达慢病毒质粒购自美国Addgene公司,DMEM培养基和胎牛血清(美国Gibco公司),胰蛋白酶(美国Sigma公司),RNA提取试剂、反转录试剂盒(PrimeScriptTMRT Master Mix)、实时荧光定量PCR试剂盒(SYBR Pre-mix Ex TaqⅡ)购自日本TaKaRa公司,鬼笔环肽染色试剂盒、嘌呤霉素和聚凝胺试剂(上海翊圣生物科技公司),RIPA裂解液及蛋白酶抑制剂以及青、链霉素(上海碧云天公司),兔抗人KDM2B多克隆抗体(美国Millipore公司),兔抗人FAK多克隆抗体(美国CST公司), 鼠抗人GAPDH单克隆抗体、羊抗兔IgG(H+L)抗体(武汉三鹰生物公司),HRP偶联的抗小鼠/兔IgG二抗(杭州联科生物公司)。
1.2细胞培养 HEK-293T细胞、人卵巢癌HO8910细胞用DMEM培养液(含10%胎牛血清+1%青、链霉素)培养,并置于5%CO2、37 ℃恒温培养箱中。每2~3 d换液1次,待细胞融合度达80%时,用2.5 g/L胰蛋白酶消化传代,取生长状态良好的细胞用于后续试验。
1.3稳定敲减和过表达KDM2B卵巢癌细胞系的建立 在美国Sigma Aldrich公司网站(https://www.sigmaaldrich.com)检索人KDM2B shRNA序列:CCGGCGGCCTTTACAAGAAGACATTCTCGAGA
ATGTCTTCTTGTAAAGGCCGTTTTTGAATT,送广州艾基生物公司构建KDM2B shRNA慢病毒载体。将KDM2B shRNA慢病毒质粒、KDM2B过表达的质粒分别与包装质粒psPAX2和pMD2.G共转染HEK-293T细胞。各质粒的比例为目的质粒∶psPAX2∶PMD2.0G=3∶2∶1。 按照Lipofectamine 3000转染试剂说明书进行转染,收集病毒,感染细胞。分别将3组慢病毒原液(空载慢病毒、KDM2B shRNA病毒和KDM2B过表达病组)加入HO8910细胞,聚凝胺浓度为 8 μg/mL,感染24 h后换液, 48 h 后进行二次感染。待细胞融合度达70%时,用嘌呤霉素(1 mg/mL)筛选2周,获得稳定敲减和过表达KDM2B的HO8910卵巢癌细胞系。
1.4F-actin荧光染色法 实验分为3组:HO8910对照组(慢病毒空载组)、HO8910-KDM2B敲减组和HO8910-KDM2B过表达组。将多聚赖氨酸处理的无菌细胞圆形爬片置于24孔细胞培养板内,2.5 g/L胰蛋白酶消化各组细胞成单个细胞悬液,铺板,待细胞融合度达50%时,按照F-actin染色说明书进行细胞骨架蛋白染色,步骤:4%多聚甲醛溶液固定细胞,PBS清洗3次;0.5% Triton-X 100溶液透化处理细胞,PBS清洗3次;每组加200 μL TRITC标记的鬼笔环肽工作液,浓度为5 μg/mL,PBS洗涤细胞3次;每孔加入200 μL DAPI溶液,对细胞核进行复染,PBS洗涤,封片。荧光共聚焦显微镜下观察3组细胞骨架蛋白F-actin的表达及排列的变化。实验重复3次。
1.5KDM2B敲减组转录组学测序 采用10 cm细胞培养皿培养HO8910对照组及KDM2B敲减组细胞,待细胞融合度达80%时,加入TRizol试剂,由上海康成生物公司采用Illumina Hiseq 4000测序仪进行转录组测序,并对2组细胞差异表达基因进行Pathway富集分析。利用统计学算法(Fisher′s exact test)找出1组差异表达基因和KEGG数据库(http://www.genome.jp/kegg)中的哪些生物学通路功能条目联系最大,分析结果中每个Pathway条目对应1个统计值P-value(表示显著性),P-value越小表示该Pathway条目和输入的差异表达基因联系越大,即该组差异表达基因大部分具有该Pathway条目对应的描述功能。
1.6免疫荧光染色试验 试验分组同上,待细胞融合度达50%时,参照文献[5]进行免疫荧光染色,步骤:4%多聚甲醛固定细胞,PBS洗涤3次;加入Triton X-100C透化处理,PBS洗涤3次,每次5 min;3% BSA封闭1 h,加入兔抗人FAK多克隆抗体(1∶100稀释),4 ℃过夜;PBS洗涤3次,每次5 min,避光温育FITC标记的荧光二抗(1∶200稀释)1 h;加入抗荧光猝灭剂,封片,荧光共聚焦显微镜下观察3组细胞FAK蛋白荧光强度及细胞分布变化。实验重复3次。
1.7western blot 试验分组同1.4,各组细胞达80%融合度时,用预冷PBS缓冲液洗涤2次,每孔加入200 μL RIPA 裂解液,冰上裂解30 min,刮下细胞,4 ℃、12 000 r/min离心10 min,吸取蛋白质上清, BCA法测定蛋白质浓度,加入5×loading buffer煮沸,进行SDS-PAGE电泳,每组蛋白质样本40 μg,电泳条件60 V 40 min,110 V 80 min,使用转膜仪将蛋白质转移至PVDF膜上,用50 g/L脱脂奶粉室温封闭2 h,温育加入兔抗人KDM2B多克隆抗体(1∶1 000稀释),鼠抗人GAPDH单克隆抗体(1∶5 000稀释);使用HRP偶联的抗小鼠/兔IgG二抗(1∶5 000稀释)室温温育2 h,将 PVDF 膜与曝光液充分混匀后,进行曝光,以检测KDM2B蛋白的表达水平。采用ImageJ软件进行蛋白质条带灰度值分析,以目标蛋白灰度值/对应内参灰度值作为该组目标蛋白的相对表达量,再将对照组设置为1进行标准化,实验组蛋白质表达量=(实验组蛋白质相对表达量/对照组蛋白质相对表达量)。
1.8RNA提取、cDNA逆转录及RT-qPCR 按照RNA提取试剂盒说明书提取细胞总RNA,采用Nano-Drop分光光度仪检测核酸浓度,吸光度(A260 nm/A280 nm)在1.8~2.0的样本用于逆转录实验。根据逆转录试剂盒说明书将RNA逆转录为cDNA。根据NCBI数据库中GenBank提供的人KDM2B(Gene ID:84678)、ITGA6(Gene ID:3655)、ITGB1(Gene ID:3688)和β-actin(Gene ID:60)的基因序列,采用Primer Premier 5.0软件进行引物序列设计,并由上海生工生物公司合成,引物序列见表1。 qPCR反应体系为20 μL,包括SYBR Premix ExTaq Ⅱ(2×)10 μL,10 μmol/L上、下游引物各0.8 μL,cDNA模板2 μL,ddH2O 6.4 μL。每组设3个复孔。循环参数:95 ℃预变性5 min;95 ℃ 5 s,58 ℃ 30 s,72 ℃ 30 s,共39个循环。采用荧光定量PCR仪配套的LC480软件采集65 ℃时的荧光信号并进行熔解曲线分析。实验重复3次。以β-actin为内参,采用2-ΔΔCt法计算基因相对定量的表达水平,公式:ΔCt=Ct目的基因-Ctβ-actin,ΔΔCt=ΔCt实验组-ΔCt对照组。

表1 RT-PCR引物序列
1.9统计学分析 采用GraphPad Prism5软件对数据分析和作图。两组间数据比较采用两样本的t检验,以P<0.05为差异有统计学意义。
2 结果
2.1KDM2B敲减及过表达组中KDM2BmRNA和蛋白质的水平 RT-qPCR结果显示,与对照组相比,KDM2B敲减组KDM2BmRNA水平下降(1.0±1.0 vs 0.48±0.05,t=20.10,P<0.01),KDM2B过表达组KDM2BmRNA水平升高(1.0±1.0 vs 2.33±0.23,t=9.89,P<0.05)。western blot结果显示,与对照组相比,感染shKDM2B慢病毒的HO8910细胞中KDM2B蛋白的表达水平下降(图1),蛋白质灰度扫描结果显示,两组间差异有统计学意义(1.0±1.0 vs 0.53±0.05,t=17.22,P<0.01);此外,KDM2B过表达组KDM2B蛋白的表达水平升高(图1),蛋白质灰度扫描结果显示两组间差异有统计学意义(1.0±1.0 vs 2.37±0.28,t=8.563,P<0.05)。

图1 western blot检测HO8910卵巢癌细胞中KDM2B敲低及过表达后的水平
2.2F-actin免疫荧光染色检测KDM2B敲减及过表达组中细胞骨架排列 F-actin免疫荧光染色结果显示,与对照组相比,过表达KDM2B后,F-actin表达明显增多,细胞骨架排列更规则有序;而敲低KDM2B后,F-actin表达减少,排列受到破坏(图2)。
2.3转录组测序法分析KDM2B敲减组基因富集的信号通路 Pathway分析结果显示,差异表达的基因主要集中在FAK(Focal-adhesion)信号通路(图3A)。采用RT-qPCR检测FAK信号通路上游分子ITGA6、ITGB1mRNA的表达,结果显示敲低KDM2B后,ITGA6(1.0±1.0 vs 0.51±0.03,t=28.29,P<0.05)及ITGB1(1.0±1.0 vs 0.53±0.10,t=8.266,P<0.05)mRNA水平降低,过表达KDM2B后,ITGA6(1.0±1.0 vs 1.9±0.36,t=5.129,P<0.05)及ITGB1(1.0±1.0 vs 2.39±0.48,t=4.958,P<0.05)mRNA水平升高。免疫荧光结果显示,KDM2B促进了FAK蛋白的表达(图 3B)。

注:F-actin荧光染色检测KDM2B对卵巢癌细胞骨架排列的调控作用,蓝色代表细胞核,红色代表细胞骨架F-acin(×40)。
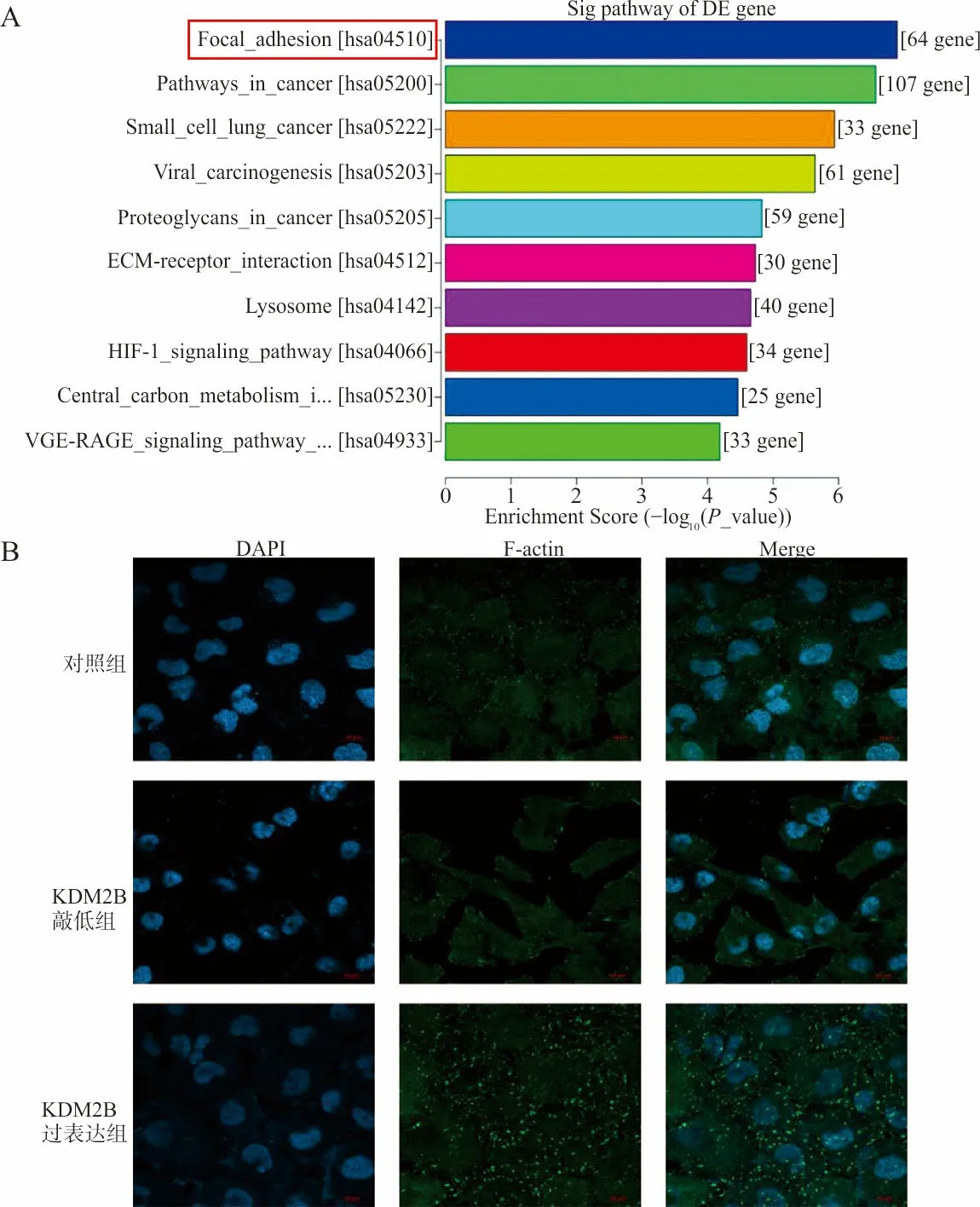
注:A,干扰KDM2B后,转录组测序细胞信号通路分析图; B,免疫荧光试验检测KDM2B对FAK蛋白的分布及表达水平的调控作用,蓝色代表细胞核,绿色代表FAK蛋白,放大倍数×40,标尺20 μm。
2.4GEPIA肿瘤数据库分析FAK在卵巢癌组织中的表达及其与KDM2B的相关性 利用GEPIA肿瘤数据库分析卵巢癌组织中FAK的表达水平,结果显示相对于正常对照组织,卵巢癌组织中FAK的表达上调(图4);相关性分析显示,卵巢癌组织中,FAK与KDM2B存在正相关(r=0.37,P<0.01)。

图4 GEPIA肿瘤数据库卵巢癌大样本中FAK的表达水平
3 讨论
尽管分子靶向治疗、免疫疗法等技术快速发展,但是晚期卵巢癌患者的五年生存率仍低于30%[6]。肿瘤转移是影响卵巢癌患者预后的一个重要因素。研究证实,肿瘤转移早期需要依赖细胞骨架蛋白F-actin聚合形成特殊细胞突起来破坏基底膜,侵入组织、血管或淋巴管[7]。破坏细胞骨架排列已经成为一种具有临床应用前景的抗肿瘤策略[8]。本研究成功构建了稳定敲低和过表达KDM2B的卵巢癌细胞系,并且证实KDM2B调控细胞骨架蛋白F-actin排列。进一步的试验结果证实,高表达KDM2B后,F-actin表达增加,排列更加规则有序,成矛状伸展,提示细胞具有更高的动力;而敲低KDM2B后,F-actin表达减少,排列受到破坏,没有规则。已有的研究证实,KDM2B可以调控ERRα蛋白的稳定性,进而促进乳腺癌细胞的增殖[9];KDM2B还可以调控Hippo信号通路,促进胰腺导管腺癌(PDAC)进展[10]。另外,研究还发现KDM2B可以通过let-7b-EZH2轴促进卵巢癌的进展[11]。上述研究表明,KDM2B具有促进肿瘤进展的作用。本研究结果支持KDM2B可能通过调控细胞骨架排列来促进卵巢癌细胞迁移,侵袭,进而促进卵巢癌转移。但KDM2B如何调控F-actin排列尚需深入研究。
本研究采用转录组测序技术在KDM2B敲低细胞组中发现差异表达的基因主要集中于FAK相关的信号通路。FAK是一种非受体酪氨酸激酶,可通过整合素和其他细胞表面受体介导信号转导,从而调节细胞的黏附、迁移、存活、增殖和分化等[12]。研究表明,FAK可以调控细胞骨架排列,促进细胞迁移[13];抑制FAK信号通路,可以破坏F-actin的聚合,进而抑制肿瘤细胞迁移侵袭[14]。此外,FAK信号通路激活还可以导致卵巢癌细胞化疗耐药,细胞迁移、侵袭能力增强[15-16]。上述研究提示,KDM2B可能通过激活FAK信号通路来调控F-actin排列。笔者进一步的研究结果证实,KDM2B上调FAK通路上游分子ITGA6及ITGB1的表达,免疫荧光试验证实KDM2B能促进FAK的表达。因此,笔者推测KDM2B通过活化ITGA6-ITGB1/FAK信号通路,促进卵巢癌细胞骨架重塑。
综上所述,本研究初步探讨了KDM2B在卵巢癌细胞骨架排列中的调控作用,证实KDM2B促进F-actin表达及重塑,并进一步证实KDM2B活化了ITGA6-ITGB1/FAK信号通路,据此推测,KDM2B在卵巢癌转移过程中可能发挥重要角色。本研究亦存在一定局限性,首先,本研究仅在HO8910卵巢癌细胞系验证了KDM2B对细胞骨架的调控作用,后续需要在其他卵巢癌细胞系验证,以揭示KDM2B对卵巢癌细胞骨架调控作用的普遍性。其次,本研究并没有深入探讨KDM2B调控ITGA6-ITGB1/FAK/F-acin信号通路的具体分子机制。最后,本研究并未在动物实验中验证KDM2B对卵巢癌转移的调控作用。
猜你喜欢
昆明医科大学学报(2022年1期)2022-02-28
科学与生活(2021年16期)2021-11-25
青岛大学学报(医学版)(2021年5期)2021-11-17
伴侣(2021年6期)2021-08-03
家庭百事通·健康一点通(2020年12期)2020-12-31
小资CHIC!ELEGANCE(2018年28期)2018-09-14
分析化学(2016年12期)2017-02-04
小资CHIC!ELEGANCE(2016年15期)2016-07-26
中国民族民间医药·下半月(2014年4期)2014-09-26
中国实用医药(2009年5期)2009-05-08
