
真菌毒素毒性及其在生物样本中检测技术研究进展
2021-01-18 13:16:24孙梅峰豆小文车轶群欧阳臻杨美华
中国药理学与毒理学杂志 2020年10期
孙梅峰,豆小文,张 磊,车轶群,赵 明,欧阳臻,杨美华
(1.江苏大学药学院,江苏 镇江 212013;2.中国医学科学院北京协和医学院药用植物研究所,中草药物质基础与资源利用教育部重点实验室,北京 100193;3.国家癌症中心,国家肿瘤临床医学研究中心,中国医学科学院北京协和医学院肿瘤医院检验科,北京 100021)
真菌毒素是由真菌产生的一类具有高水平生物蓄积效应的低分子质量的有害次生代谢物[1]。已知真菌毒素种类多达400余种[2],广泛存在于食品、农作物、饲料及中药材中。在一定生态环境如温度、相对湿度[3-4]和杀菌剂[5]等因素影响下,可导致真菌毒素的产生。其中对人和动物危害大并引起世界多国高度关注的主要有黄曲霉毒素(aflatoxins,AF)、赭曲霉毒素(ochratoxin,OT)、呕吐毒素(deoxynivalenol,DON)、T-2毒素、玉米赤霉烯酮(zearalenone,ZEN)、伏马毒素(fumonisin,FB)和展青霉素(patulin,PAT)等[6]。真菌毒素可通过受污染的粮食、霉变中药、受侵染饲料喂养的动物等途径进入人类食物链,对人体和动物产生严重危害,主要表现为肝、肾、造血系统、免疫系统和生殖系统等毒性[7]以及致癌和致畸作用[8-9]。国际癌症研究机构(International Agency for Research on Cancer,IARC)根据危险程度的高低,在致癌物列表[10]中将真菌毒素分类为Ⅰ类致癌物质(AF)和ⅡB 类致癌物质(OTA,FB1,FB2)、Ⅲ类致癌物质(PAT)等。联合国粮食及农业组织和世界卫生组织/WHO食品添加剂委员会多次针对AF,OTA和FB等真菌毒素进行了危险性分析,并且暂定了每周最大耐受摄入量和每日最大耐受摄入量,世界各国也针对真菌毒素制定了限量标准。
真菌毒素发现至今,于2004-2005年便出现过肯尼亚AF急性中毒事件,因对农副产品和中药真菌毒素污染防控措施有限,导致真菌毒素可通过多种摄入途径在人体内蓄积,进而可能引发其潜在慢性毒性。如今,真菌毒素毒性研究已从单一毒素靶器官损伤深入至多毒素在细胞、分子水平上的联合作用机制,检测技术也由传统液相色谱发展至LC-MS/MS、免疫传感器、分子印迹等高通量、高灵敏、高特异性的痕量检测。通过对临床样本检测分析人体实际摄入量,即可客观评价某种毒素对人体健康的危害[11]。本文基于文献研究成果,就单一真菌毒素毒性研究及混合真菌毒素基质共存的现状,对单一毒性和联合毒性研究进行归类分析,简析不同类型生物样本的前处理手段和检测方法,以期为体内真菌毒素检测研究及人体真菌毒素暴露风险评估提供参考。
1 典型真菌毒素毒性
1.1 曲霉属真菌毒素毒性
曲霉属真菌毒素是研究较多的真菌毒素,按产毒菌类别可分为黄曲霉类、赭曲霉类和杂色曲霉类等。
AF是一类结构相似的二氢呋喃香豆素衍生物,常见种类有AFB1,AFB2,AFG1,AFG2和AFM1等,其中AFB1毒性和致癌性最强,其毒性为氰化钾的10倍,砒霜的68倍[12]。早在1993年,WHO国际癌症研究机构就将其列为Ⅰ类致癌物质。VESEL等[13]通过对鸡胚给药观察几种AF的生长毒性,发现AF之间存在毒性差异,其毒性大小依次为:AFB1>AFG1>AFM1=AFB2>AFG2。近期,葛军华等[14]研究表明,AFB1的心肌毒性可能是通过诱导心肌细胞线粒体损伤以及调控凋亡相关蛋白的表达,促进心肌细胞凋亡而产生的。徐明芳等[15]实验发现,AFB1还可诱导体外培养的人胚肝细胞L-02中晚期凋亡,发生G0/G1期阻滞,进一步丰富了AFB1毒性作用机制。刘珊等[16]研究显示,AFB1对原代培养的人皮肤成纤维细胞HSF也具有明显细胞毒性。
OT主要是由赭曲霉(Aspergillus.ochraceus)、纯绿青霉(Penicillum.verrucosum)和炭黑曲霉(A.carbonarius)产生的1-氧代异色满-7-甲酰胺类化合物,主要有 OTA,OTB,OTC 和OTα[17-18],其中OTA毒性最大,包括肾毒性、肝毒性、免疫毒性和致癌致畸等[19]。有研究指出,巴尔干地方性肾病可能就与OTA有关[20]。ZHU等[21]通过ig给予大鼠,进一步阐明了OTA是利用miR-122通过细胞周期蛋白G1/P53通路和BCL-W/胱天蛋白酶3通路诱导肝细胞凋亡,产生肝毒性。
杂色曲霉毒素是由以杂色曲霉为主的包括构巢曲霉、黄曲霉和寄生曲霉等50多种真菌所产生的聚酮类化合物[22],具有致肝损伤、基因毒性和致癌作用,被IARC归为ⅡB类致癌物[10]。多个动物实验证实,杂色曲霉毒素可以通过抑制T细胞表达来削弱免疫系统对致癌物质的监测功能表现出免疫毒性[23-24]。
1.2 镰刀菌属真菌毒素毒性
镰刀菌属真菌毒素通常按不同结构可分为单端孢霉烯族类、伏马类和玉米烯酮类等毒素。
单端孢霉烯族毒素是一类倍半萜烯化合物,主要有A,B,C和D 4类分型,其中A和B类较为常见且毒性较大。单端孢霉烯A类毒素种类主要有T-2毒素和HT-2;B类毒素主要有雪腐镰刀菌烯醇(nivalenol,NIV)、DON和镰刀菌烯酮(fusarenon-X,Fu-X)等[25],其毒性作用主要有生长毒性、生殖毒性、基因毒性、皮肤毒性及严重的血液毒性[25]。此外,ZHANG等[26]利用小鼠神经母细胞瘤(N2a)细胞研究表明,T-2毒素是通过诱导N2a细胞产生氧化应激、线粒体功能障碍和通过抑制核因子E2相关因子2/血红素加氧酶-1通路和激活P53通路引起细胞凋亡,从而产生神经毒性。
FB是主要由串珠镰刀菌产生的一类长链多羟基醇丙三酯类水溶性化合物,可引起肝、肾损伤甚至坏死及马脑白质软化症、猪肺水肿[27-28]等毒性反应。其结构与神经鞘氨醇类似,可以阻断体内二氢神经酰胺的形成,进一步阻断神经酰胺的合成,从而影响鞘脂类物质的代谢,扰乱体内叶酸的摄取和利用,人体摄入后可能引起食管癌和神经管型缺陷病[29-30]。
ZEN是主要由镰刀菌属真菌产生的一类具有二羟基苯酸内酯结构化合物,是世界范围内粮食饲料主要污染物之一,在动物源性食品中也有检出[31]。研究表明,ZEN对动物具有很强的生殖毒性和致畸作用,其生殖毒性主要是由于ZEN具有类雌激素作用,可能与胞浆中雌激素受体高度亲和,从而对雌性动物的生殖系统产生毒性作用[32]。
1.3 混合真菌毒素的联合毒性
在实际检测研究中,食品及饲料中往往存在不止一种真菌毒素。据报道,全球各类商品中污染的真菌毒素有127种[33],同时存在的混合真菌毒素主要包括AF+OTA,AF+FB及DON+ZEN,检出频率依次为21%,20%和13%。2018年BioMin公司统计,在全球13 629份样品中频繁检出的真菌毒素为FB,DON和ZEN。在多种真菌毒素共存模式下,其毒性作用远比单一毒素毒性简单相加更复杂,真菌毒素毒性之间可能存在拮抗、加和以及协同等效应。JIAN等[34]通过小鼠对比实验发现,DON和ZEN对小鼠肝代谢的联合毒性弱于单一毒素组,两者表现出明显的拮抗作用。李文竹等[35]发现,DON和AFB1作用于HepG2/C3A细胞引起的细胞凋亡表现为加和作用。此外,混合真菌毒素在不同剂量下可能表现不同的相互作用。WAN等[36]利用IPEC-J2细胞研究镰刀菌属类真菌毒素时发现,在低浓度DON和NIV联合毒性表现出拮抗作用,而在高浓度时为协同作用。因此,对混合真菌毒素联合毒性效应进行深入研究十分必要。
2 生物样本中典型真菌毒素及其标志物检测
2.1 样品前处理
生物体中真菌毒素含量低且样本基质复杂,为提高检测的选择性和灵敏度,样品净化必不可少。生物样本前处理主要采用的净化方法有蛋白质沉淀(protein precipitation,PPT)、液液萃取(liquidliquid extraction,LLE)、固相萃取(solid phase extraction,SPE)、免疫亲和净化(immunoaffinity column,IAC)和直接稀释(dilute-and-shoot)等1种或2种方法联用。
2.1.1 血清和血浆
血浆的组成包括水、蛋白质、磷脂、氨基酸和尿酸等有机物及无机盐等[37]。以磷脂酰胆碱为例,作为磷脂类两性化合物,在电喷雾离子源(ESI)正、负离子模式下均能引起待测物离子抑制效应[38],从而影响检测结果的准确性,蛋白质、磷脂大分子去除不完全则会堵塞色谱柱。因此,在体内痕量真菌毒素及其标志物检测分析中,有效的净化处理必不可少。目前血浆和血清样本中主要采用LLE法,或LLE法联合SPE或IAC法对目标物进行纯化富集处理(表1)。其中单一真菌毒素OTA研究最多。萃取溶剂以三氯甲烷、二氯甲烷和乙腈最为常用,尤其是多种真菌毒素及标志物共同检测研究中,乙腈和乙腈-丙酮混合溶剂作为极性至中等极性化合物的良好溶媒,毒性相对较小,应用最广。SLOBODCHIKOVA等[39]研究人血浆中17种真菌毒素及其标志物时发现,与SPE和PPT法相比,三步LLE法能极大程度降低基质效应的同时保留良好回收率。此外,LLE法的成本相对较低,更加经济。
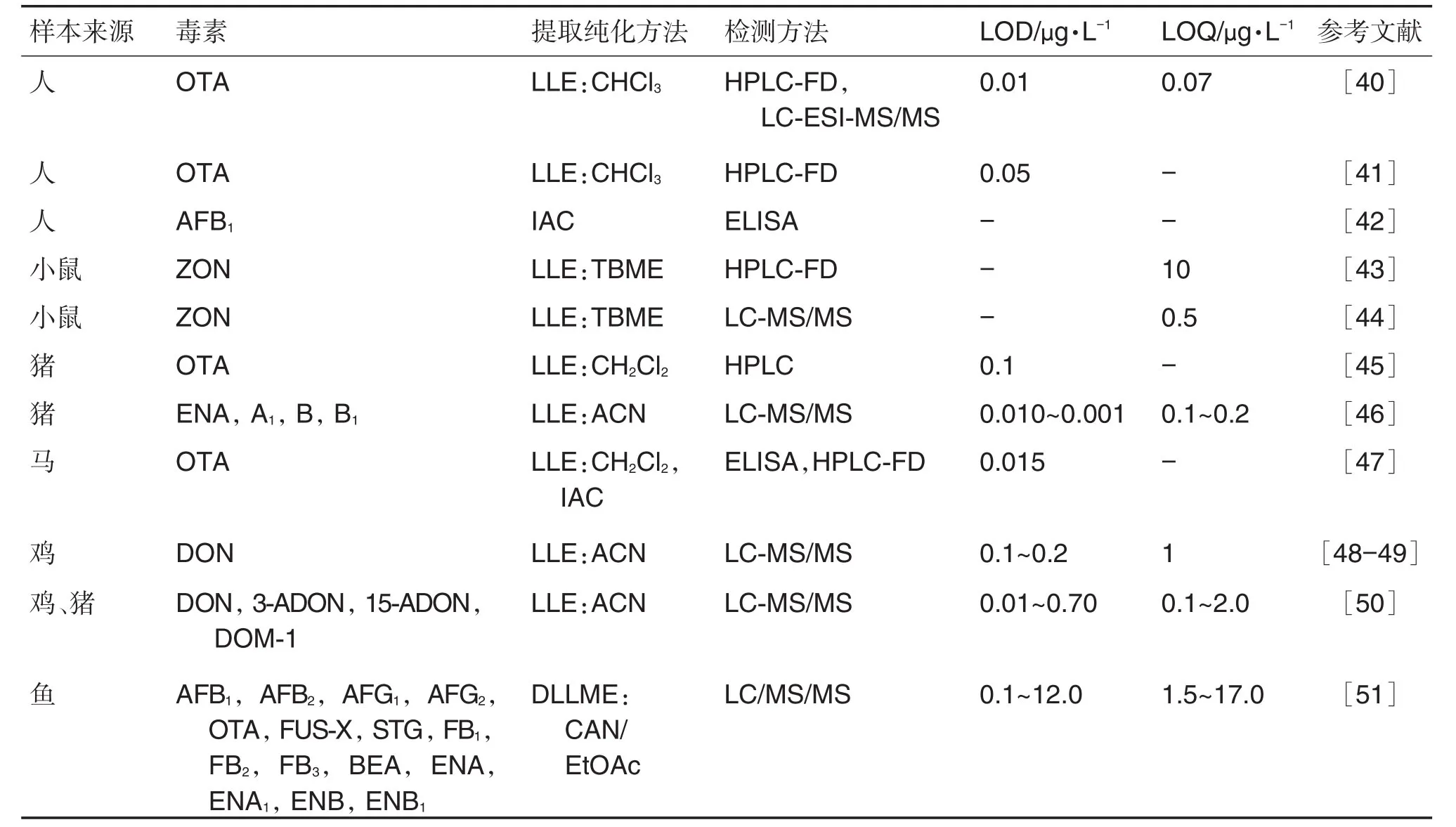
表1 血清和血浆样本中真菌毒素的检测
目前色谱科技领域的各大公司相继推出了除磷脂、蛋白质的前处理产品如Phree磷脂去除板、Oasis PRiME MCX小柱和Cleanert®PPT 96孔蛋白沉淀板等。与传统LLE法和PPT法相比,此类产品利用多功能化的复合吸附材料通过离子交换、反相、氢键等原理有效去除磷脂蛋白干扰基质,同时因其96孔板的独特设计免去了离心和液体转移的繁琐步骤,大大节省了前处理时间,减少了工作量,还能减少上样量,有望在检测血样真菌毒素及其痕量标志物中得以推广应用。
2.1.2 尿液
尿液样本多以真菌毒素及其代谢标志物为研究目标,且尿液基质干扰以无机盐为主。因此,多采用直接稀释法或过滤稀释法联合SPE柱、IAC柱等前处理方法,以确保最大程度上获得样本中的真菌毒素(表2)。痕量检测研究中往往需要同时考虑目标物的富集和样本净化,AHN等[52]在研究人尿液中的真菌毒素时,利用IAC柱抓捕靶标的特性对样本中的目标物进行净化富集,并利用同位素内标的方法进行定量分析,方法准确可行但成本较高。盐析液液萃取(salting-out liquid-liquid extraction,SALLE)利用盐析原理促进均相溶液分层达到对待测物分离净化的目的,适合常规LLE法无法萃取的亲水性目标物,RODRÍGUEZ-CARRASCO等[53]在分析人尿液样本中10种镰刀菌毒素及其代谢物水平时,对比了分散液液微萃取(dispersive liquid-liquid microextraction,DLLME)和盐析液液萃取(saltingout liquid-liquid extraction,SALLE)法。结果表明,SALLE法提取效率优于DLLME法,回收率84%~96%,同时成本也比DLLME法更经济。
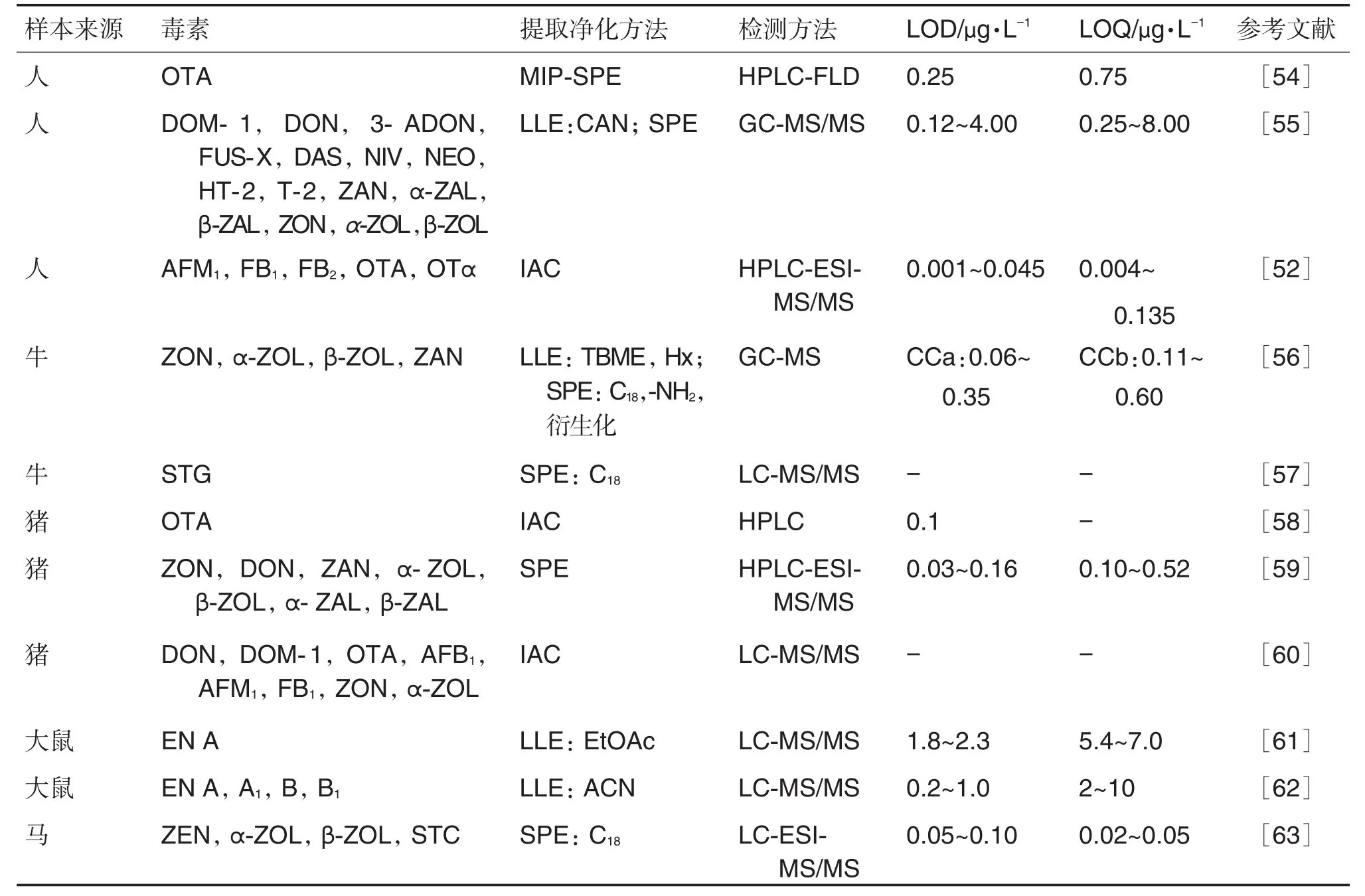
表2 尿液样本中真菌毒素的检测
2.1.3 组织器官
组织器官中除真菌毒素及其代谢物外还有毒素加合物的存在,且基质相比其他体内样本复杂。因此,器官组织中真菌毒素研究多采用均质法处理样本,以正己烷等非极性溶剂脱除脂肪干扰,并加入酸或酶等促使加合物(如AFB1-DNA加合物:AFB1-甲酰嘧啶和iro-AFB1-DNA等;AFB1-白蛋白加合物等)还原为毒素原型,再利用LLE法联合IAC和SPE等净化样本(表3)。游丽娜等[64]利用β-葡萄糖苷酸/硫酸酯复合酶对样本酶解后,利用LLE法联用IAC进行处理,对鸡蛋中6种玉米赤霉烯醇类化合物残留进行检测,方法灵敏度高,重现性好。李玉平等[58]在测定OTA时对猪肾组织进行酸解、萃取等步骤后,采用IAC进行净化处理。
2.2 检测技术
2.2.1 薄层色谱法(thin-layer chromatography,TLC)
自20世纪60-70年代,有关真菌毒素问题进入人类视线,10年间与其有关的研究方法大多基于AF自然荧光的TLC法及溶剂提取优化[79]。魏润蕴等[80]建立了猪组织中OTA的TLC测定方法,利用甲醇-水-碳酸氢钠提取猪组织中OTA,经过石油醚、三氯甲烷萃取净化后,采用双向展开法对样品进行分析。由于TLC法检测效率低,试剂易挥发,灵敏度有待提高,其应用在生物样本中已鲜有报道。
2.2.2 高效液相色谱法(high performance liquid chromatography,HPLC)
HPLC也称高压液相色谱,区别于纸色谱、TLC等经典液相色谱法。因其高灵敏度、高自动化、特异性强、分析效率高等特点而被广泛用于体内微量真菌毒素检测。方晓明等[81]基于玉米赤霉醇(zearalanone,ZAN)262 nm处具有紫外吸收,利用二极管阵列检测器(photo-diode array,PDA)建立了一种HPLC测定鸡肝中ZAN的方法。SANGARE-TIGORI等[82]利用OTA的荧光特性,采用荧光检测器(fluorescence detector,FLD)检测,在22名(34.9%)健康志愿者中检出OTA平均含量为0.83 μg·L-1;在8名(20.5%)肾病患者体内检出OTA,平均含量是1.05 μg·L-1。基于紫外或荧光检测方法的前提是样本中待测真菌毒素或其衍生物具有可引起紫外吸收或产生荧光光谱的化学官能团,然而,并不是所有真菌毒素具备上述条件。因此,HPLC-PDA或FLD在生物样本中痕量真菌毒素及其代谢标志物的检测应用存在局限性。
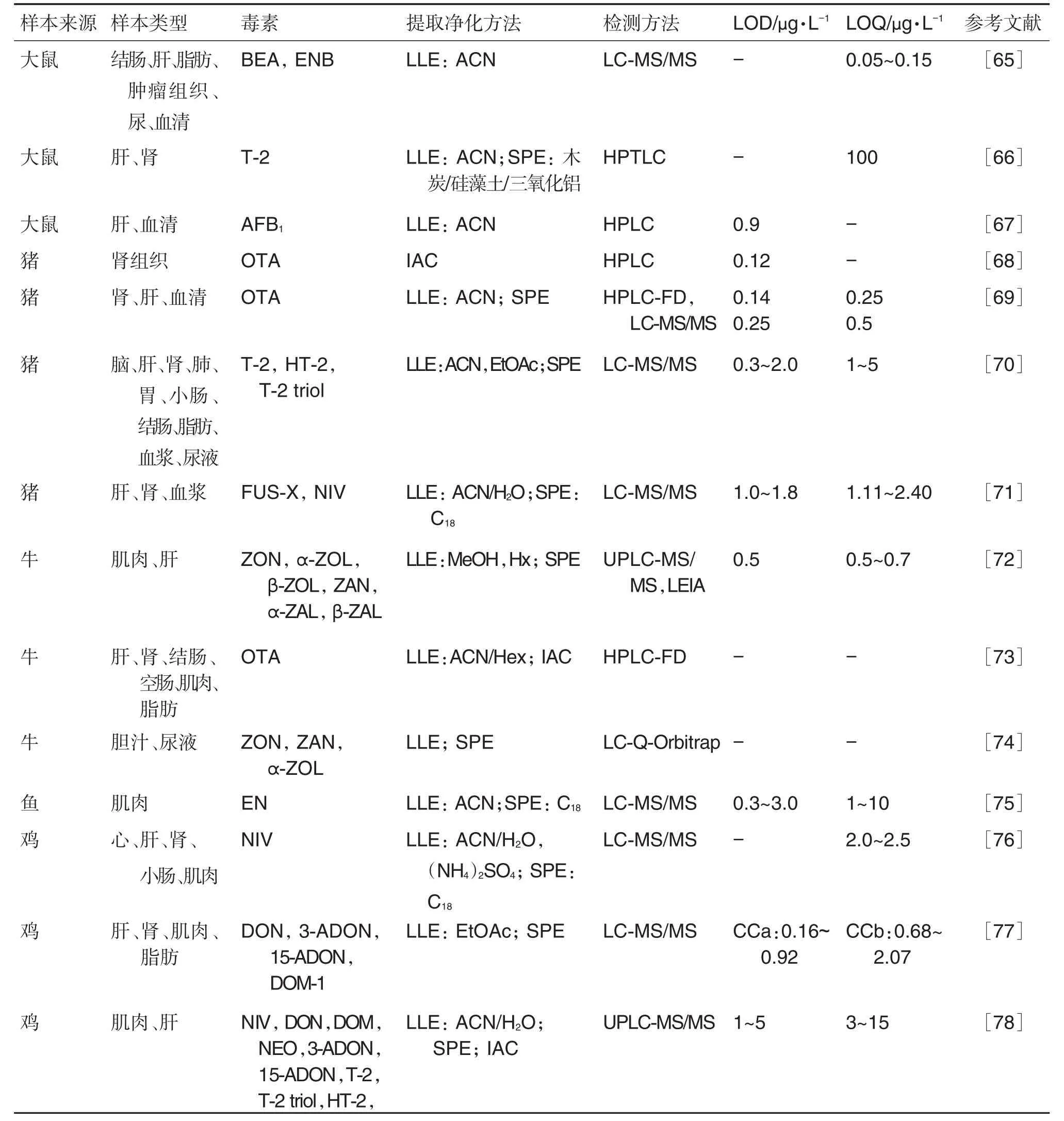
表3 器官组织中真菌毒素的检测
2.2.3 色谱-质谱联用法
液相色谱-串联质谱法(liquid chromatographytandem mass spectrometry,LC-MS/MS)不仅具备液相色谱法的准确定量、高分离效能等优点,同时利用质谱的高灵敏度、可定性和高特异性等特点,弥补了液相色谱法无法准确定性的缺点,提高了方法适用性,为检测多种毒素提供了同时定量定性的方法,因此在生物样本中应用广泛。QIU等[83]利用UPLC-MS/MS分析国内15个省份的母乳中10种真菌毒素的暴露水平。结果表明,我国母乳中真菌毒素含量总体处于较低水平,但仍有AFB1和AFM1等6种真菌毒素检出。WANG等[84]建立了LC-MS/MS检测动物血浆中21种真菌毒素及其代谢产物方法,可同时对样本中多种真菌毒素进行快速、准确的检测,但所需个体样本量(2 mL)较大,不适用人体暴露情况的分析研究。LC-MS/MS法相比HPLC-PDA或FLD法,灵敏度有很大改善,但实际人体样本研究中,如何降低样本采样量并兼顾其检测灵敏度和稳定性仍是体内分析的研究热点。BERND等[85]开发了无创采集样品制备方法,将血液或血清样本100 μL收集于滤纸卡上干燥保存,建立了人血液样本中27种真菌毒素及代谢物HPLCMS/MS检测方法,该法的样品制备过程具有无创、制备简单、成本损耗低和储存便利等特点。
气相色谱-质谱联用法(gas chromatographymass spectrometry,GC-MS)主要用于测定具有热稳定性、挥发性以及化学衍生的非挥发性真菌毒素,因此具有一定局限性。BLOKLAND等[56]用GC-MS对牛尿液中的ZAN及其代谢产物进行了测定。RODRÍGUEZ等[53]基于气相色谱-质谱联用建立了一种检测人尿液中10种镰刀菌属真菌毒素及其代谢物的高灵敏度、定量、定性准确的方法。
2.2.4 免疫分析法
近年来,随着真菌毒素抗体制备技术的发展,基于免疫分析原理的快速筛查方法及传感技术已逐渐应用到体内真菌毒素的检测中。以示踪标记物分类,体内生物标志物免疫分析法主要包括酶联免疫吸附法(enzyme linked immunosorbent assay,ELISA)、时间分辨免疫荧光技术(time-resolved fluoroimmunoassay,TRFIA)和放射免疫测定法(radioimmunoassay,RIA)。
ELISA采用酶高效催化实现检测信号的放大标记,具有灵敏度高、特异性强、操作简便、成本低的优点。PESTKA等[86]利用ELISA快速、灵敏的特点,对大鼠体内DON代谢分布情况进行监测研究。此外,有关伊朗等国家地区共计183例母乳中OTA,AFM1和ZEN等真菌毒素暴露情况的研究也采用了ELISA[87-88],该方法最大的优势是简易的前处理方式,无需复杂的样品净化、富集等处理即能达到良好的灵敏度;所需的样本量较少(<100 μL),适用于大样本量的筛选检测。
TRFIA是将具有特殊荧光的镧系离子与螯合剂结合标记抗原、抗体、蛋白等作为信号源,经过生物识别反应(抗原抗体的特异性结合、核酸探针的杂交反应等),利用时间分辨荧光仪通过测定荧光强度变化实现目标物的定量分析[89-90]。TRFIA法具有测量范围宽、检测效率高、重复性好等特点[91-93],已应用于临床肿瘤标志物CA19-9和乙型肝炎病毒(HBV)等研究[91,94-95]。在真菌毒素的检测中也有广阔的应用前景。TANG等[96]首次利用抗特异性纳米与单抗结合的方式,建立了2种具有竞争性的时间分辨条带法用于检测玉米制品中的AF和ZEN。结果显示,方法定量可靠,灵敏度高且避免了剧毒抗原的使用。
RIA是通过同位素标记的和未标记的抗原同抗体发生竞争性抑制反应,通过测量反应后反应产物放射性而求得未标记抗原(目标物)含量的分析方法。孙桂菊等[97]建立了一种敏感特异的AFB1-白蛋白加合物(AFB1-ALB)的测定方法,利用Microcon-50微型浓缩器对白蛋白快速分离,最后制得的样品用液体闪烁仪进行检测,该法通过装置内的酶解消化,减少了转移损耗,提高了回收率和分析方法的准确性。唐耘天等[98]采用RIA法探究了肝癌高危人群中AFB1-ALB水平和谷胱甘肽转移酶的基因多态性之间的关系。结果表明,谷胱甘肽转移酶的基因多态性在决定AF暴露水平上起重要作用,方法灵敏度虽高,但操作步骤繁琐,耗时长,稳定性不佳。
2.2.5 其他检测技术
随现代科学技术发展,传统检测技术不断完善的同时诞生了诸多快速检测技术,如适配体(aptamer)传感器技术、噬菌体展示技术(phage display technology,PDT)、胶体金技术等。
适配体传感器技术,是以通过指数富集的配体系统进化技术(systematic evolution of ligands by exponential enrichment,SELEX),从人工合成的寡核苷酸文库中筛选得到的能够与靶分子高亲和力、高特异性结合的寡核苷酸配基作为核心部件的检测技术。因其在高温和不良环境下的稳定性、靶分子类型无限制、无免疫原性、无毒等特性被广泛应用于各类研究,大量基于OTA的适配体生物传感器已有报道,但在体内样本的检测应用鲜有报道。DINCKAYA等[99]通过电化学阻抗谱和循环伏安法测量结合在胱胺层中的适配体修饰的金纳米粒子的相互作用,检测了牛奶中的AFM1。搭建的生物传感器特异性强,灵敏度高,重复性和再现性良好,可用于实际样本的检测分析,但电化学阻抗谱的方法相对耗时,传感器的搭建和使用较为精细,因此在推广使用上还存在一定局限性。
PDT是以噬菌体或噬菌粒为载体,通过基因工程技术使外源肽或蛋白随噬菌体外壳蛋白一同表达在噬菌体表面的技术,PDT利用众多的单一融合型噬菌体组成的噬菌体展示库来实现高通量的筛选[100]。PDT筛选得到的真菌毒素模拟抗原表位代替真菌毒素偶联抗原建立的免疫学检测方法,不仅增加了人员的操作安全,而且降低了成本。WANG等[101]利用PDT制备的AF无毒替代抗原的纳米抗体Phage2-5建立AF的ELISA检测方法,方法灵敏度高,基质效应低,且对甲醇、盐离子的耐受能力强。WANG等[102]同样利用PDT获得了对黄曲霉毒素具有高度反应的PO8-VHH抗体,并开发了一种高选择性的纳米抗体夹心ELISA用于快速检测农产品中的曲霉菌。PDT与其他生物检测技术相结合为同时检测多种真菌毒素提供了更广阔的前景。
3 结语
真菌毒素发现至今,在各类农副产品及中药材中均有检出的报道,随真菌毒素毒性作用研究的深入,其危害风险受到了越来越多的重视,世界各国政府均对真菌毒素制定了相应的限量标准。我国作为中药使用大国,对中药中真菌毒素也制定了相关限量标准和检测方法。
日常生活中,人和动物难免会摄入真菌毒素并通过吸收代谢在体内蓄积,最终对人和动物健康产生危害。因此,人们逐步展开了对真菌毒素暴露情况的研究。由于人与动物复杂的食物链和代谢系统,体内通常存在一种以上的真菌毒素。但由于基质效应以及其他影响因素存在,从体内检出的真菌毒素及其代谢物种类有限,因此针对不同生物样本和研究目标,采取高效的前处理手段和检测技术至关重要。合适的前处理方法不仅可提高方法回收率,简化操作步骤,节约成本,还可降低基质效应影响,提高方法灵敏度。目前最为普遍的前处理方法是一步法的LLE或LLE与柱净化联用的两步法。常用的萃取溶剂有乙腈、乙酸乙酯、二氯甲烷、丙酮和甲醇等。而根据实际情况选择合适的检测技术有利于提高方法灵敏度和重复性。LC-MS/MS因其出色的分离能力、定性能力、广泛的适用性以及高灵敏度成为目前对多目标同时进行高效定量、定性检测的常用方法之一。生物传感器技术等快检技术的兴起,为体内样本真菌毒素快速检测提供了研究方向。同时为进一步研究人体真菌毒素暴露情况,应尝试向样本制备的低成本和微量样本,检测技术的高通量和高特异性等方面进行突破。
猜你喜欢
考试与评价·高二版(2021年3期)2021-09-10 07:22:44
数学物理学报(2020年5期)2020-11-26 06:06:28
科学(2020年2期)2020-08-24 07:56:56
小哥白尼(野生动物)(2019年5期)2019-08-27 00:53:38
天然产物研究与开发(2018年8期)2018-09-10 05:48:38
小学科学(学生版)(2018年8期)2018-08-21 12:05:50
公民与法治(2016年14期)2016-05-17 04:15:03
吉林大学学报(医学版)(2015年4期)2015-12-17 07:48:10
微生物与感染(2015年5期)2015-12-08 07:03:20
发明与创新(2015年33期)2015-02-27 10:40:02
