
云斑白条天牛幼虫肠道细菌的分离与鉴定
2020-11-03 06:39朱林慧方小英段慧娟杨振德
广西林业科学 2020年3期
朱林慧,方小英,段慧娟,徐 蕾,杨振德
(广西大学林学院,广西南宁 530004)
云斑白条天牛(Batocera lineolata)为天牛科(Cerambycidae)多食性昆虫,主要危害枇杷(Eriobotrya japonica)、无花果(Ficus carica)、柑橘(Citrus reticulata)、乌木(Echeveria agavoides'Ebony')、泡桐(Paulownia fortunei)、苹果(Malus pumila)、桃(Amygdalus persica)和核桃(Juglans regia)等[1]。其分布范围广,繁殖能力强,适应性强,是短周期用材林和经济林的重要蛀干害虫[2-3]。在中国,云斑白条天牛有害生物风险性PRA值为2.04,属于高度危险性林业有害生物[1]。
昆虫的肠道中存在着大量微生物,这些微生物与昆虫联系广泛[4]。一些微生物与昆虫宿主在长期协同进化中形成了紧密的共生关系[5]。昆虫为肠道微生物提供稳定的生存环境和必要的营养物质,肠道微生物参与昆虫体内的多种代谢过程,在昆虫的营养、代谢和免疫等诸多生理功能上发挥重要作用[6-7]。李丹红等[8]研究发现,不同类型白蚁的肠道细菌均在白蚁取食木材后帮助其对木材进行降解。蚜虫共生菌(Buchnera)能产生多种氨基酸,弥补蚜虫单一吸食植物汁液导致的营养不足[9]。
周峻沛[10]利用传统的微生物培养方法从云斑白条天牛幼虫的肠道内分离出100 株细菌,发现假单胞菌属(Pseudomonas)、拉乌尔菌属(Raoultella)和环丝菌属(Brochothrix)为其优势菌属。谢谷艾等[11]分离与鉴定了松褐天牛(Monochamus alternatus)幼虫肠道微生物,发现固氮菌在松褐天牛幼虫肠道中普遍存在。陈金华[12]利用常规16S rDNA 克隆文库和RFLP 分析,发现桑粒肩天牛(Apriona germari)幼虫肠道中有丰富的微生物,优势菌属为克雷伯氏菌属(Klebsiella)、乳球菌属(Lactococcus)和肠球菌属(Enterococcus)。目前,关于云斑白条天牛具体龄级幼虫肠道微生物尚未有报道,不同龄级间肠道微生物差异还需进一步研究。云斑白条天牛4龄幼虫生长速度最快,取食量大,是控制其生长与危害的关键时期,研究此龄期的肠道微生物尤为重要。本研究采用传统细菌培养结合16S rDNA 基因测序的方法,对云斑白条天牛4龄幼虫肠道内的可培养细菌进行分离与鉴定,为运用生物防治云斑白条天牛的新技术提供可利用的微生物资源,也为云斑白条天牛具体龄级幼虫肠道共生菌的进一步研究提供参考。
1 材料与方法
1.1 材料
1.1.1 供试虫源
2018年5—6月上旬,在广西国有高峰林场六里分场(108°20'E,22°58'N)的桉树林中采集云斑白条天牛4 龄幼虫,作为样品。该地年均相对湿度80%以上,年均气温19.3 ℃。
1.1.2 主要器材及试剂
主要器材为昆虫针、解剖剪、镊子、蜡盘、玻璃匀浆器、涂布器、接种环、高压蒸汽灭菌锅、高速冷冻离心机(德国Sigma 公司)、电子天平、垂直流超净工作台(上海智城分析仪器制造有限公司)、博迅BSC-400 恒温恒湿培养箱、细菌基因组DNA 抽提试剂盒(天根DP302)、Life Touch PCR、Nanodrop 2000 可见分光光度计和DYY-8C 电泳仪等。在南宁市羽瑞生物试剂经营部购买培养基原料、细菌基因组DNA 抽提试剂盒和其他分析纯试剂,由生工生物工程(上海)股份有限公司合成引物(27F 和1492R)。LB 基础培养基配方参考万伦[13]的配制方法,将10 g 蛋白胨、10 g NaCl、5 g 酵母膏和18 g 琼脂用蒸馏水定容至1 000 mL,pH 值7.0,121 ℃灭菌20 min。去掉琼脂后得到LB 液体培养基。
1.2 方法
1.2.1 云斑白条天牛幼虫肠道的提取
参考周峻沛[10]的方法进行解剖。所有操作均在紫外杀菌过的超净工作台中进行,所用器械均进行高压灭菌或75%酒精消毒。取健康的云斑白条天牛4 龄幼虫放入4 ℃冰箱30 min,使其活动能力减弱,用无菌水冲洗虫体表面杂质,再用75%酒精冲洗并浸泡3 min,随后用无菌水冲洗3 次。在蜡盘上用昆虫针固定虫体,用解剖剪将虫体剪开,用镊子取出肠道,将肠道上连带的脂肪等无关物质用无菌水冲洗并用镊子拨离后放入匀浆器中,加入1 mL无菌水充分研磨,制成幼虫肠道匀浆液。
1.2.2 肠道细菌的分离和纯化
将制好的肠道匀浆液在4 ℃、4 000 rpm 条件下离心10 s,取上清液,用无菌水梯度稀释103、104和105倍,分别取不同倍数的匀浆液各100 μL,均匀涂布在LB 培养基上,每种稀释倍数重复3 次,以无菌水涂布及消毒后的云斑白条天牛幼虫在平板上滚动摩擦后的平板为对照(CK)。在27 ℃培养箱中倒置培养48 h,长出菌落后,用接种环从每种稀释梯度的培养基上挑取大小、形态和颜色有差异的单菌落,在新的LB 培养基上进行数次4 区划线分离,直至得到纯的单克隆菌株。将纯的单克隆菌株接种到LB 液体培养基中,加入25%的甘油后,放置冰箱(-80 ℃)贮存备用。
1.2.3 细菌鉴定
对分离纯化得到的菌株进行细菌形态及培养性状鉴定、染色反应和生理生化特性鉴定,根据《伯杰细菌鉴定手册》[14]和《常见细菌系统鉴定手册》[15]进行判定。
将分离纯化得到的单克隆菌株接种到LB液体培养基中,30 ℃条件下振荡培养48 h后,取2 mL细菌悬液,11 500 r离心1 min,倒去上清液,使用细菌基因组DNA抽提试剂盒(操作步骤按说明进行)分别提取所有试验菌株的总DNA,所提取DNA的OD260/OD280比值使用Nanodrop 2000 可见分光光度计进行测定,继而确定DNA 纯度和浓度,并利用1%琼脂糖凝胶电泳检测条带是否可以进行后续试验。PCR扩增以提取出来的各细菌DNA为模板,以细菌通用引物为扩增引物,即正向引物27F(5’-AGAGTTTGATCCTG-GCTCAG-3’)和反向引物1492R(5’-GGTTACCTTGT-TACGACTT-3’)。PCR反应体系为12.5μLMix,1.0μL27F(10mmol/L)和1.0μL1492R(10mmol/L),1.0μLDNA模板(50ng/μL)12.5 μL ddH2O。PCR反应条件为预变性条件95℃5min;进行35个循环的条件94℃30s,56℃30 s,72 ℃1 min;终末延伸条件72 ℃10 min;最后4 ℃条件下保存。PCR扩增反应结束后,利用1%琼脂糖凝胶对PCR产物进行电泳检测,若检测合格,送往北京睿博兴科生物技术有限公司进行16S rDNA 基因测序。将测序结果在NCBI 平台(https://www.ncbi.nlm.nih.gov/)上进行Nucleotide BLAST序列比对,根据数据库中与测试菌株序列同源性最高的序列确定菌株的分类,并利用MEGA 7.0建立系统发育树。
2 结果与分析
2.1 形态鉴定
通过对云斑白条天牛幼虫体内可培养细菌的分离与纯化,共获得14株不同的单克隆菌株(表1)。将14株菌株划线接种到LB培养基上培养1天后,进行革兰氏染色鉴定,根据染色结果,10 株菌株革兰氏反应为阴性,4 株为阳性。使用油镜观察后发现11 株菌株形态呈杆状,3 株呈球状;5 株菌株有柱状芽孢,9 株没有;7 株菌株有周生鞭毛,1 株有端生鞭毛,1株有侧生鞭毛,5株没有鞭毛;5株菌株有荚膜,9株没有。
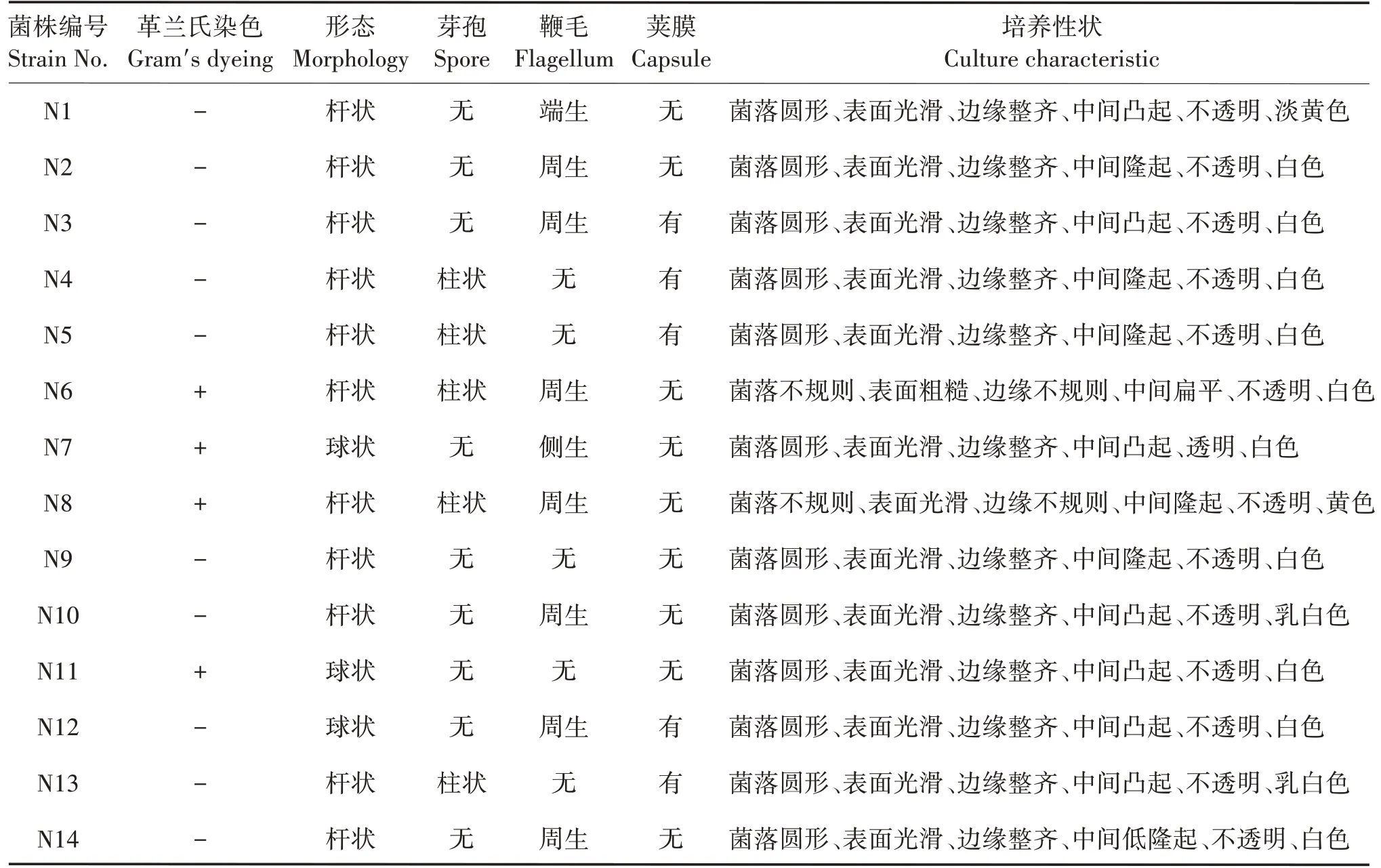
表1 云斑白条天牛幼虫肠道可培养细菌形态及培养性状Tab.1 Morphology and culture characteristics of culturable bacteria in intestinal tracts of B.lineolata larvae
2.2 生理生化
对14株菌株进行不同的碳素化合物、氮素化合物和酶的生理生化特性鉴定,14株菌株属于糖、醇类发酵型,且不产生色氨酸脱氨酶、苯丙氨酸脱氨酶和吲哚;除N11菌株外,其余菌株均产生过氧化氢酶;除N5菌株外,其余菌株V-P试验均为阴性反应(表2)。

表2 云斑白条天牛幼虫肠道内可培养细菌的生理生化特性Tab.2 Physiological and biochemical characteristics of culturable bacteria in intestinal tracts of B.lineolata larvae
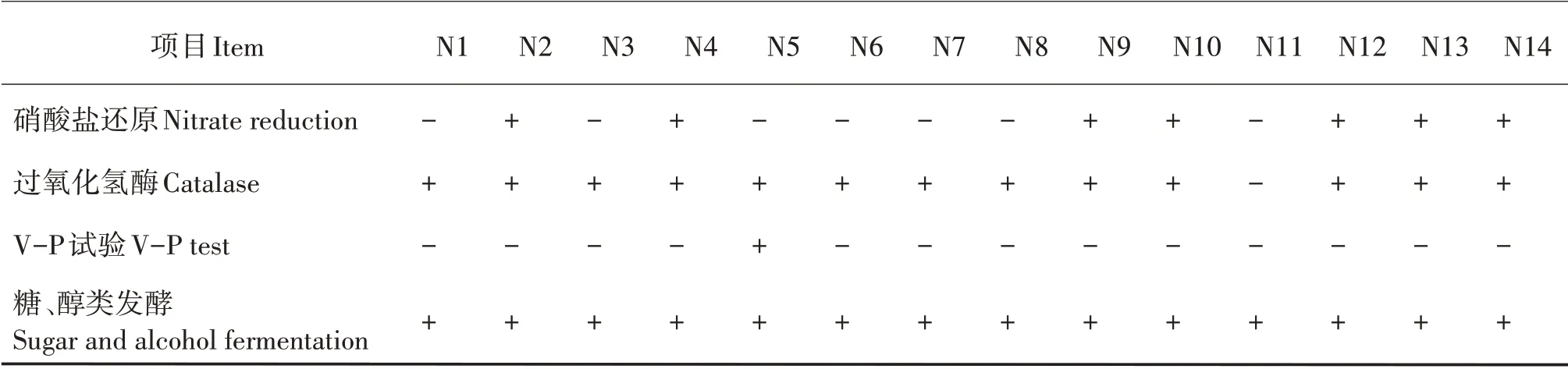
续表2 Continued
2.3 可培养细菌的16S rDNA 基因序列分析
将测序结果在NCBI 上进行序列比对,结合RDP数据库Classifier分析,14种细菌分属于9个属,分别为芽孢杆菌属(Bacillus)、肠球菌属、肠杆菌属(Enterobacter)、柠檬酸杆菌属(Citrobacter)、拉乌尔菌属、苍白杆菌属(Ochrobactrum)、寡养单胞菌属(Stenotrophomonas)、Gibbsiella和Kosakonia(表3)。其中,芽孢杆菌属2株(N6和N8),肠球菌属2株(N7和N11),肠杆菌属1 株(N14),柠檬酸杆菌属2 株(N2 和N10),拉乌尔菌属3 株(N4、N5 和N13),苍白杆菌属1 株(N3),寡养单胞菌属1 株(N1),Gibbsiella1株(N9),Kosakonia1株(N12)。

表3 云斑白条天牛幼虫肠道内可培养细菌16S rDNA 序列比对结果Tab.3 16S rDNA sequence identification results of culturable bacteria in intestinal tracts of B.lineolata larvae
2.4 可培养细菌的系统发育分析
将测序得到的云斑白条天牛4龄幼虫肠道内可培养细菌的16S rDNA 序列进行系统发育进化分析。结果显示,其肠道内可培养细菌可归类于9 个属,5个科,分别在进化树上形成独立的分支(图1)。其中,N14、N1、N2、N10、N9、N4、N5、N13和N12形成以变形菌门的γ-变形菌纲(γ-proteobacteria)为家族的大分支,除N1 属于假单胞菌科(Pseudomonadaceae)的寡养单胞菌属外,其余均属于肠杆菌科(Enterobacteriaceae)的肠杆菌属、柠檬酸杆菌属、Gibbsiella、拉乌尔菌属和Kosakonia;N3 独自形成以变形菌门(Proteobacteria)的α-变形菌纲(α-proteobacteria)为家族的分支,为布鲁氏菌科(Brucellaceae)的苍白杆菌属;N6、N8、N7 和N11 形成以厚壁菌门(Firmicutes)为家族的分支,均为芽孢杆菌纲(Bacilli),分别属于芽孢杆菌科(Bacillaceae)的芽孢杆菌属和肠球菌科((Enterococcaceae)的肠球菌属。系统发育树进一步支持了RDP Classiffier的分类结果。

图1 基于16S rDNA序列的云斑白条天牛幼虫肠道内可培养细菌系统发育进化树Fig.1 Phylogenetic tree of culturable bacteria in intestinal tracts of B.lineolata larvae based on 16S rDNA sequences
3 结论与讨论
本研究采用传统微生物分离培养的方法从云斑白条天牛4 龄幼虫肠道内分离和纯化出14 种细菌,通过形态观察并结合16S rDNA 基因测序技术,鉴定出这14种可培养细菌分属于寡养单胞菌属、柠檬酸杆菌属、苍白杆菌属、拉乌尔菌属、芽孢杆菌属、肠球菌属、肠杆菌属、Gibbsiella和Kosakonia。其中,变形菌门的拉乌尔菌属为优势菌属。
本研究显示,14株细菌中有10株归类于变形菌门,4 株归类于厚壁菌门,说明在门水平上,变形菌门是云斑白条天牛4龄幼虫肠道内可培养细菌的优势菌群。已有研究发现,鞘翅目(Coleoptera)昆虫中,光肩星天牛(Anoplophora glabripennis)[16]、暗黑鳃金龟(Holotrichia parallela)[17]和新西兰助翅鳃金龟(Costelytra zealandica)幼虫[18]的肠道优势菌均为变形菌门和厚壁菌门。鳞翅目(Lepidoptera)昆虫中的稻纵卷叶螟(Cnaphalocrocis medinawlis)[19]、舞毒蛾(Lymantria dispar)[20]、小菜蛾(Plutella xylostella)[21]以及双翅目(Diptera)的南亚果实蝇(Bactrocera tau)[22]和柑橘大实蝇(Ceratitis capitata)[23]的肠道优势菌也均为厚壁菌门和变形菌门。说明变形菌门与厚壁菌门是不少昆虫体内微生物的优势菌门,它们可能对昆虫体内的生理代谢活动有重要影响。
云斑白条天牛4龄幼虫体内的优势菌属为拉乌尔菌属,其次是芽孢杆菌属、肠球菌和柠檬酸杆菌属等。有研究发现,拉乌尔菌属中的很多种类具有降解多种环境污染物的作用,昆虫肠道内的拉乌尔菌可能有助于昆虫的抗药性[24]。室内飞蝗(Locusta migratoria)种群相比于田间飞蝗种群,其后肠细菌中的优势菌(拉乌尔菌属)所占比例有所下降,这可能是野外昆虫抗药性相对较强的原因之一[25]。芽孢杆菌属为严格好氧或兼性厌氧菌[26],可消耗肠道内氧气,使其形成厌氧环境,为有益微生物(乳酸杆菌等)的生长提供良好环境,且有较强的分泌胞外酶的能力和丰富的代谢产物[27]。肠球菌在很多昆虫肠道微生物中均有发现,相辉等[28]发现家蚕(Bombyx mori)幼虫肠道微生物的主要优势菌群为肠球菌,其丰度高达82%;鲁兴萌等[29]研究发现在家蚕肠道中起着抵御病原微孢子虫发芽增殖作用的是优势菌(肠球菌)。柠檬酸杆菌属常在双翅目昆虫中分布,橘小实蝇(bactrocera dorsails)[30]和南亚实蝇(Bactrocera tau)[31]的优势菌属均为柠檬酸杆菌属,该类细菌为化能有机营养型,可利用糖酵解与戊糖磷酸途径降解碳水化合物[32]。云斑白条天牛幼虫体内相同的菌群是否具有相似的功能还需做进一步研究。
昆虫体内微生物多样性主要受昆虫性别、发育龄期和种群等方面的影响[33]。暗黑鳃金龟幼虫肠道微生物多样性在不同龄期存在差异,其1 龄幼虫的优势菌门(拟杆菌门(Bacteroidetes))的丰度最高;在第2 和3 龄幼虫中,厚壁菌门的丰度最高[34]。周峻沛[10]从江西省上饶市虫龄不一的云斑白条天牛幼虫胃肠道中分离和鉴定出100 株细菌,分别归类于5个门、13 个属,其中优势菌门为变形菌门和厚壁菌门,优势菌属为假单胞菌属、拉乌尔菌属和环丝菌属(Brochothrix)。本研究结果中云斑白条天牛4 龄幼虫肠道内细菌的优势菌门与周峻沛的结果一致。在属的水平上,虽然均发现拉乌尔菌属为云斑白条天牛幼虫的优势菌属,但本研究未发现假单胞菌属和环丝菌属类型,分离鉴定出的细菌种类也较少。这可能与云斑白条天牛幼虫的虫龄、寄主、采集地和采集时间等的不同有关。周峻沛的采集地点为江西上饶,与广西南宁处于不同纬度,在气候、地形和温度等方面均存在差异。俞英豪[35]分别对采集于江苏南京和浙江永康的大青叶蝉(Cicadella viridis)进行肠道细菌多样性分析,发现两地叶蝉的肠道菌群存在明显差异。陈金华[12]对桑粒肩天牛幼虫的肠道多样性进行研究,发现不同月份间细菌多样性不同。据此推测不同采集时间导致温度和湿度的不同,进而影响云斑白条天牛肠道的细菌组成。本研究中云斑白条天牛的寄主为桉树,周峻沛研究的云斑白条天牛寄生于栎属(Quercus)植物。相对于栎属植物,桉树木屑成分主要为木质素,含氮量少[36],膳食基质的组成和含量对昆虫肠道细菌的丰度有一定影响[37],所以取食不同寄主的云斑白条天牛肠道微生物多样性有差异。
很多因素影响昆虫体内微生物的组成与丰度,系统探讨造成云斑白条天牛肠道细菌差异的因素对于阐明云斑白条天牛幼虫肠道细菌的功能、开发基于肠道细菌为靶标的生物防治药剂具有重要意义。
猜你喜欢
河南医学研究(2022年19期)2022-10-19
小哥白尼(野生动物)(2021年1期)2021-07-16
中国比较医学杂志(2020年4期)2020-05-26
水生生物学报(2019年4期)2019-07-20
生态学报(2019年11期)2019-07-08
生物安全学报(2019年3期)2019-02-15
川北医学院学报(2019年6期)2019-02-10
小学生必读(低年级版)(2018年10期)2019-01-04
故事作文·低年级(2018年10期)2018-10-25
作文与考试·小学低年级版(2015年11期)2015-07-17
